
Abstract
The main focus of this article is the role of lipids in biomineralization. Much of the discussion on biomineralization focuses on proteins in these decades. Indeed, collagen and acidic noncollagenous proteins effectively serve as templates for mineralization. However, other macromolecules such as lipids and polysaccharides have received less attention despite their abundance at mineralization sites. The matrix vesicle (MV) theory is widely accepted as the induction of early mineralization. Although ion concentration within the vesicles has been discussed in the initial mineralization in this theory, the role of phospholipids that constitute the vesicle membrane has not been discussed much. Comprehensive considerations, including pathological mineralization, exist regardless of the localization of MVs, the involvement of bacteria in dental calculus formation, and biomineralization caused by marine organisms such as corals, suggesting that initial mineralization found in these biological conditions might be a common reaction relating to lipids. In contrast, despite the abundance of lipids, mineralization occurs only in the limited tissue within our body. In other words, gathering knowledge and creating a path to understanding about lipid-based mineralization is extremely important in proposing new bone disease treatment methods. This article describes how lipids influence nucleation, mineralization, and expansion during hard tissue formation.
Impact statement
Recent studies have accumulated evidence of mineralization involving phospholipids and the matrix vesicle (MV) theory. Mineralization occurs not only in the conventional vesicle form but also in flat membranes arrested by the matrix. The flat membrane is derived not only from MVs but also from various causes, such as cell rupture and cell apoptosis. Mineralization is greatly affected by alkaline phosphatases derived from cell membranes. By understanding phospholipid-based mineralization, it will be possible to design new mineralization-inducing materials centered on cellular components for early bone formation. This article is important for developing new strategies to induce bone mineralization.
Introduction
Researchers from various disciplines pursuing the same goal have used various methods for testing and analysis to study bone mineralization. While most studies on bone mineralization have focused on biological variables, material science approaches have considered mineralization, nucleation, growth, and the physical and chemical environment of the surroundings of mineral formation. The primary emphasis of research has shifted in response to both the passage of time and the advancement of analytical methods. According to the findings of the Web of Science investigation, 24,492 publications were written on this subject from 1993 to 2023 (keywords: bone mineralization, Fig. 1). 1 The figure demonstrates that the research topic was aligned with various subjects and areas. Most publications covered endocrinology and metabolism, biomaterials, biomedical engineering, and cell biology.
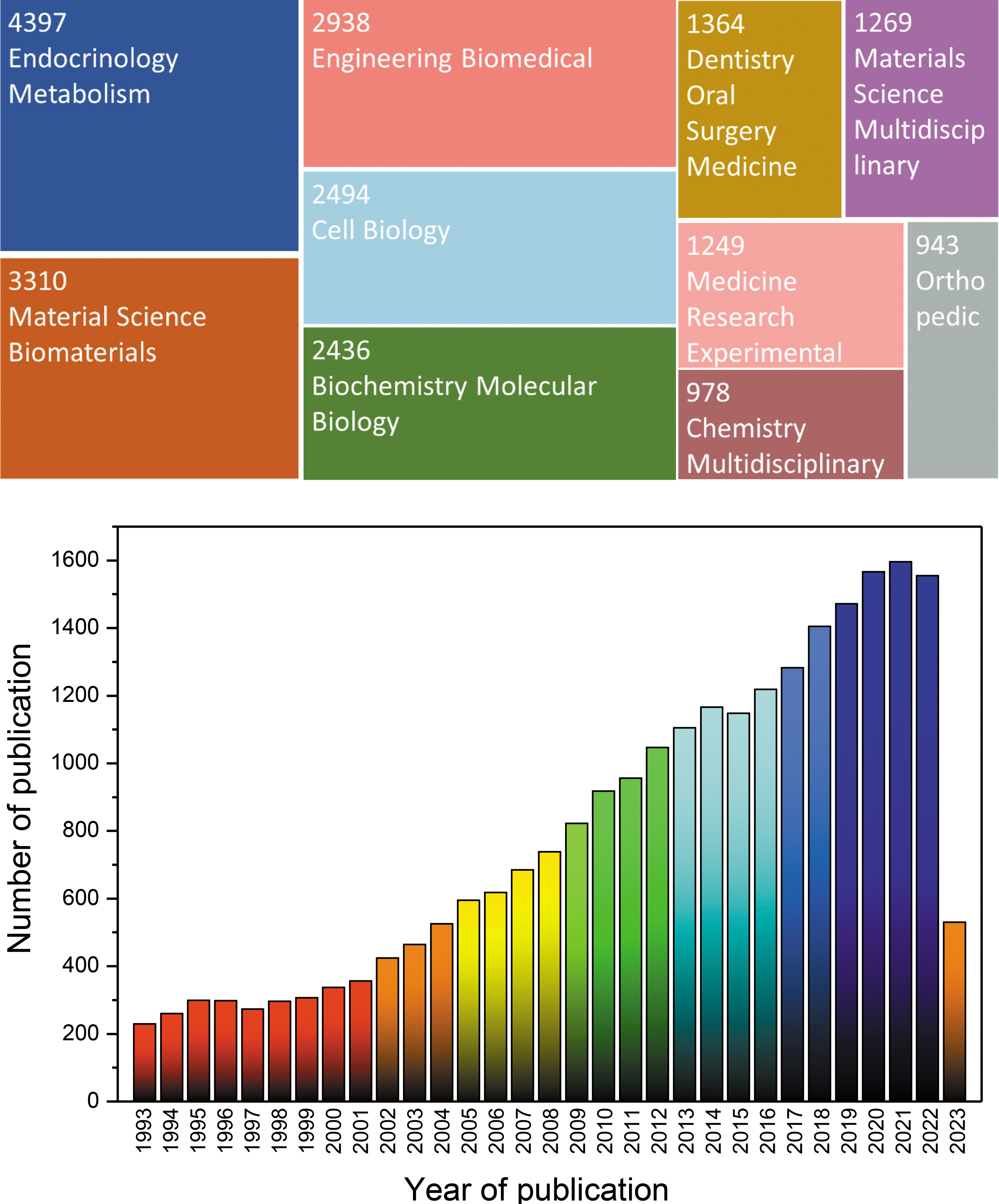
Publications for 24,492 findings from Web of Science Core Collection in 1993–2023 displayed as a tree map (Data last updated: June 14, 2023). 1 Color images are available online.
Intracellular and extracellular biomineralization involves complicated interactions between biological processes and inorganic materials to generate intricate mineral formations with distinct characteristics. During extracellular biomineralization, minerals are deposited outside the cells in the extracellular matrix (ECM) or specific biological compartments. Proteins, lipids, and polysaccharides are used as templates or regulators of mineral synthesis during extracellular biomineralization. Organic molecules affect the nucleation, growth, and morphology of minerals. Extracellular biomineralization results in the formation of vertebrate bones and teeth, calcium carbonate mollusk shells, and coral reefs. Biomineralization is also classified into bioinduced and biologically controlled biomineralization.
In addition to research related to mineral formation, extensive research has been conducted on mineralized ECMs, including calcified cartilage, bone, dentin, cementum, and enamel. This research focused on the mineralized ECM protein composition and how the ECM influences cell behavior and the organization of skeletal tissue.2–4 The structural approach was utilized to understand the hierarchical three-dimensional spatial arrangement and the structure of the various components of skeletal material.5–7 In particular, according to published research, the composition of bone is 70% mineral and only 30% organic, mainly collagenous and noncollagenous proteins (NCPs). Recent advances in protein qualification and quantification methods have accelerated research on protein components and their roles. Collagen molecules are structural materials for bone tissue, whereas NCPs supply nucleation sites to collagenous proteins through specific collagen-binding domains.
Macromolecules, such as lipids, polysaccharides, and water, are also prominent in the mineralization process; however, less attention has been paid to them. This review addresses the role of lipids in nucleation, mineralization, and expansion. This underestimates the biomolecules governing the microstructure and the physical and chemical properties of the mineralized tissue.
Lipid in Hard Tissue
In healthy individuals, physiological mineralization occurs only in certain hard tissues, such as skeletal and dental tissues. Biomineralization is the process by which minerals are incorporated into the organic ECM, which is primarily associated with collagen fibrils. In a conventional understanding of mineralization, osteoblasts start this process by creating and releasing matrix vesicles (MVs) at certain sites throughout the mineralized matrix, also known as the osteoid, before mineralization starts. Mineralization is initiated either within or around MVs in the ECM because of their high concentrations of calcium (Ca2+) and phosphate (PO43–) ions, which are ideal environments for forming minerals. MVs are vesicles composed of lipid bilayer membranes released from cells, indicating the importance of lipids in mineralization. However, recent studies have also indicated the role of other types of lipids in biomineralization.
In mineralized tissue, the significant role of lipids is primarily an old but novel concept. 8 Preliminary research from 1960s had already characterized osseous tissues' lipids from various sites in the anatomy, as well as inside the tissues that mineralized like mature dentin, bones, and enamel from bovine, human, and rat, as well as in coral skeletons.9–11 The fact that coral skeleton and enamel consist of acellular structures indicates that lipids may exist in the cemented matrix even if they are not linked to cellular membrane components. 12
Lipids may directly contribute to the nucleation process during the in vitro mineralization of cartilage. Remineralization can be readily detected within 12 h if the demineralized hypertrophic cartilage is placed in a metastable calcium phosphate solution. However, the rate of remineralization is significantly decreased if the majority of the lipids (∼80%) are removed from the decalcified matrix.13,14 In bone tissue, lipids are found in the matrix's mineralization as closely bound lipids combined with minerals and proteins to form complexes and as loosely bound lipids in the bone cells that is readily extractable. 15
The two general categories of lipids are polar and nonpolar lipids. Nonpolar lipids are insoluble in water and extremely hydrophobic. In addition to their lipid-like structures, polar lipids contain one or more ionizable or hydrophilic groups, such as organic bases (choline), acids (phosphoric acid), or carbohydrates (galactose). As a result, polar lipids can position themselves in phase, forming films or micelles in an aqueous solution. The lipid content in the form of polar, nonpolar, and total lipids from bovine fetuses was measured at different zones of calcifying tissues by three sequential extraction processes (Table 1). 16 Although it cannot be generalized, polar lipids seem more involved in mineralization.
The Proportion of Polar to Nonpolar Lipids Obtained from the Successive Extraction of Calcifying Tissues 16
Before demineralization, extract 1-chloroform-methanol at a ratio of 2:1; after demineralization, extract 2-chloroform-methanol at a ratio of 2:1, and then extract 3-chloroform-methanol-conc. HCl at a ratio of 200:100:1. Values represent the mean of at least three different samples. For total lipids, the values are presented as mean.
Mineralization on Lipid Surface
Interactions between the organic and aqueous phases, involving the physical, chemical, and molecular realms, influence the growth of minerals in naturally occurring and developed structures on organic surfaces. A significant amount of research has been conducted to gain a thorough understanding of the interfacial interactions connected to surface- or template-directed crystallization and biomineralization.17,18
Electrostatic interactions play a key role in surface-directed mineralization upon charged surfaces, 19 demonstrated through research on synthesized structures with negatively charged surfaces, such as self-assembled monolayers and thin films, Langmuir monolayers, and functionalized polymers, which can affect the production of minerals. Biomineralization is strongly associated with negatively charged macromolecules.17,20,21
Lipids may also play a complex role in the ECM. Calcium binding can be facilitated by negatively charged groups on the acidic polar lipids that are exposed on the surface of the macromolecule (Fig. 2). The cationic groups of the nitrogenous lipids likely serve as phosphate-binding sites. The fact that the phosphate binding of mixed phospholipids was reliant on earlier calcium binding suggests that a secondary complexation with inorganic phosphate is likely to occur once calcium is bound.16,22,23 With the alkaline earth element Ca, phosphatidyl serine functions as a tridentate ligand. Three of the six metal coordinates were available to receive new donor groups (e.g., phosphates). 22
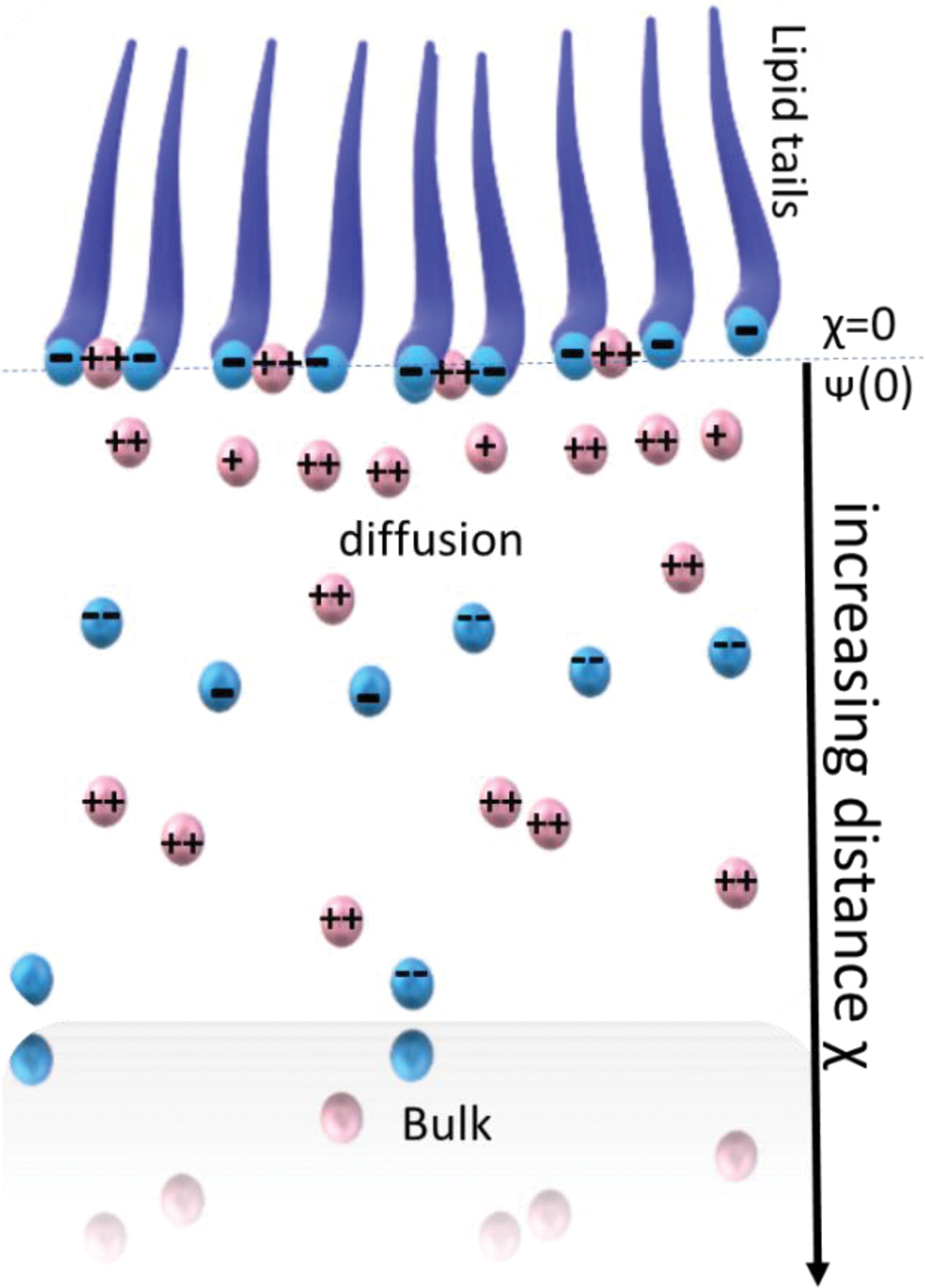
The charged organic–aqueous interface area is shown as a schematic here. One may find an orderly array of negatively charged lipid molecules at the boundary between the air and the water. The headgroup charge is assumed to be spread out in the plane at x) 0. Counterions, which are cations in this instance, may be found in two distinct states: either electrically attracted to the negative surface or bonded to it. The x) 0 plane is where cation binding takes place. The Stern surface potential is denoted by (0). 24 Color images are available online.
To gain new knowledge and a better comprehension on a molecular level of the electrostatic contributions to the formation of orientated mineral nuclei, Vogel and colleagues examined the functional link between the nucleation of minerals at planar surfaces that are charged monolayers and conventional electrostatic characteristics such as specific ion binding, surface charge density, and interfacial ion distributions (Fig. 3). 24 Negatively charged lipid molecules arranged in monolayers are connected with an electrolyte-rich aqueous solution. The two counterions in the vicinity of the interface are positively charged. The electrostatic attraction of all the counterions created a diffuse electric double layer. Some counterions, particularly the divalent ones, also exhibit surface-specific binding properties. The ratio of cations to anions found in close proximity to negatively charged surfaces is used to explain mineral nucleation. It has been hypothesized that anion speciation and directed mineral nucleation are related.
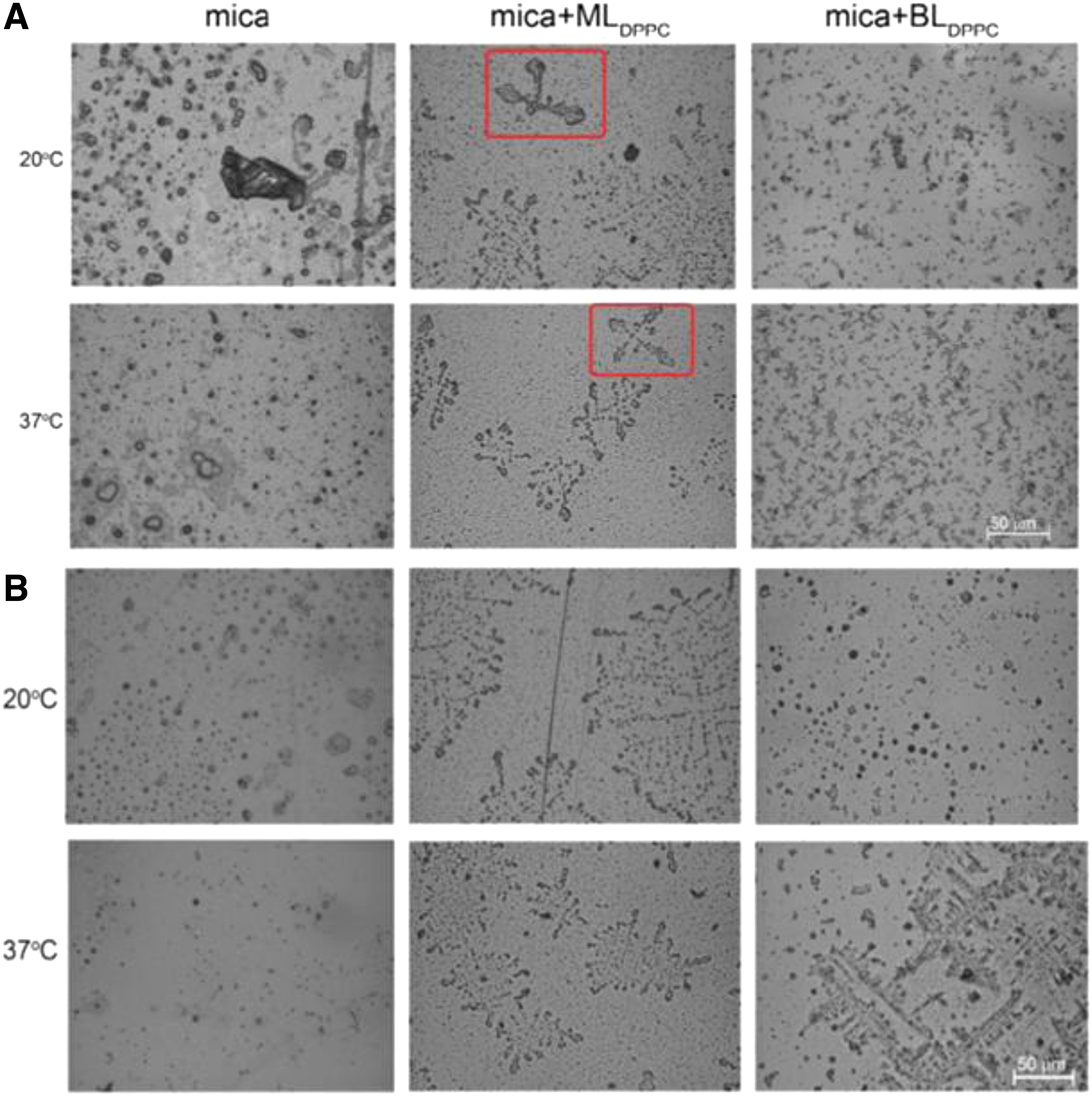
Images of minerals deposited on bare mica and DPPC-coated mica slides while they were horizontally submerged
In a physiological context, complex acidic phospholipids and proteolipids may lead to calcium phosphate mineralization. In addition to reconstituted collagen, the only lipid components involved are those separated from mineralizing tissue that are capable of triggering mineralization in vivo without cellular involvement. 25
The ability of a material to connect with bone has been assessed using acellular simulated body fluid (SBF), specifically the capacity of apatite to develop on a substance's surface. Supported phospholipid layers can be used as effective models for mineralization processes in tissues where membrane–solution interactions appear to play a major role in biological mineralization research. 26 Dipalmitoylphosphatidylcholine (DPPC) mono- and bilayers supported on mica surfaces were soaked for 2 weeks in SBF with ion concentrations approximately equal to those of human blood plasma. The phospholipid layers cause the deposition of minerals of noticeably smaller sizes. In addition, the mineral deposit on the surface of mica coated by a DPPC monolayer is arranged in an orderly manner (regularly arranged leaf-like crystals are indicated with a red box; Fig. 3).
Cruz et al. investigated calcium phosphate nucleation in Langmuir monolayers to better define the processes of calcium phosphate nucleation and growth at phosphatidylserine (PS)-enriched interfaces. PS enrichment in lipid monolayers generates regions of regionally enhanced supersaturation, which in turn initiates amorphous calcium phosphate (ACP) nucleation at the interface through a series of steps. Rapid induction of calcium phosphate nucleation was observed in 1,2-dipalmitoyl-sn-glycero-3-phosphatidylserine-enriched monolayers. Owing to the lack of mineralization in the pure DPPC monolayer, it is interesting to note that PS enrichment is the driving force that initiates mineral nucleation, thus replicating in vitro, under conditions of a membrane-like structure, the nucleation ability of the nucleation core (NC; Fig. 4). 27 These studies suggest that the phospholipid surface alone has sufficient potential to induce and modulate mineralization.
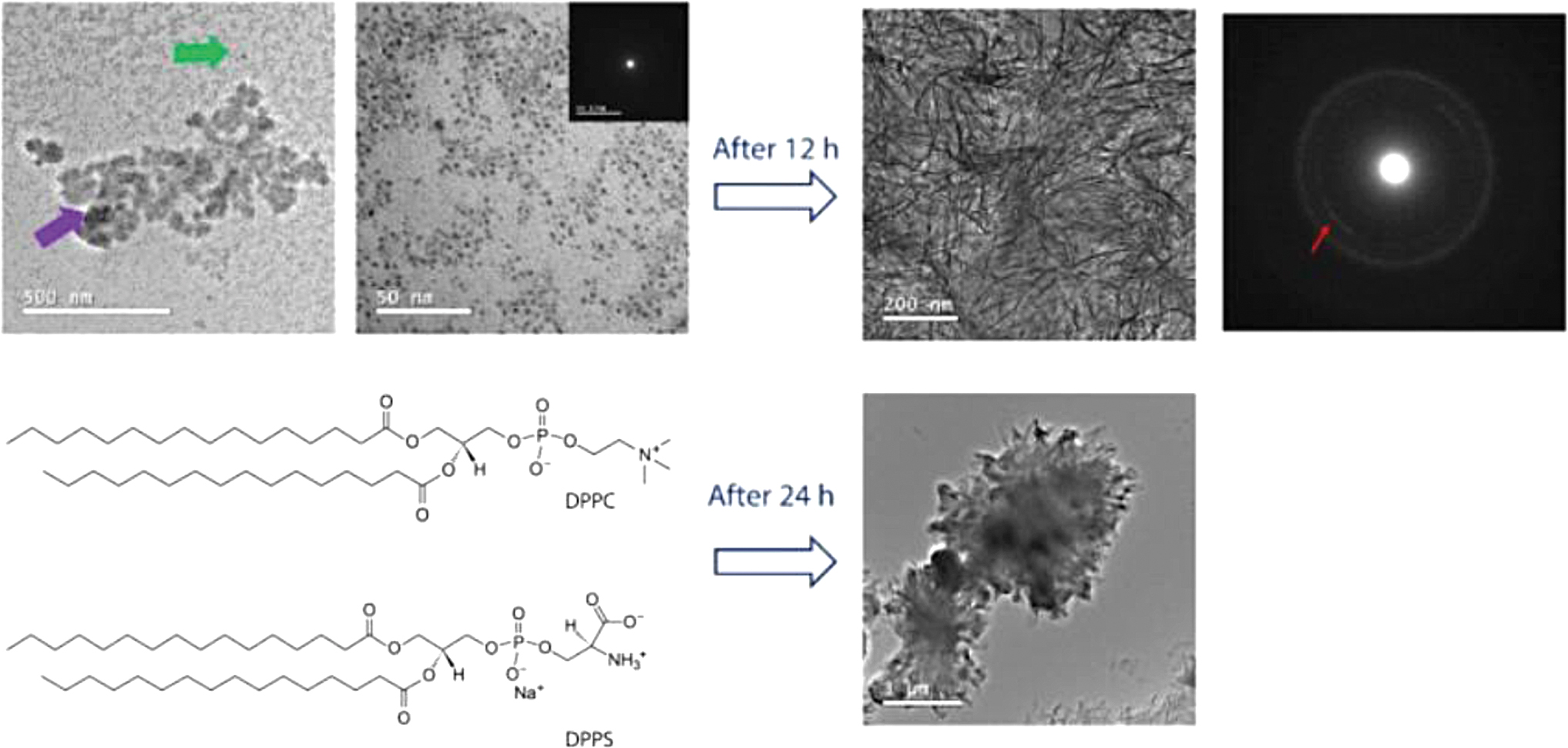
Following mineralization, the morphology of DPPS-enriched monolayers was seen. TEM pictures and their related SAED electron diffraction patterns for the monolayers of DPPC:DPPS (8:2) molar ratio. These images were transmitted after 240 min of mineralization at 25°C. The existence of nanometric complexes (<5 nm in size), as shown by the green arrow, may combine into bigger structures, as shown by the purple arrow, for the mixed DPPC:DPPS monolayer. After 12 h, the originally amorphous complexes crystallized (indicated by a red arrow in the SAED pattern). After 24 h, the creation of micrometric aggregates, as well as the total rupture of the transferred monolayer, could be detected. 27 DPPS, 1,2-dipalmitoyl-sn-glycero-3-phosphatidylserine; SAED, selected area electron diffraction; TEM, transmission electron micrographs. Color images are available online.
Mineralization using MVs
While the effectiveness of phospholipid monolayer membranes has been discussed, mineralization induction by MVs is an important process in biomineralization. Hard tissue biomineralization includes a complex spatiotemporal series of occurrences controlled by cells capable of mineralization.
Previous research has led to the theory that the initial mineral nuclei in a bone that mineralize precipitate as a result of a specific class of extracellular vesicles called MVs.28–31 Because of two primary characteristics, MVs function as mineralization nanoreactors: they exhibit a complex biochemical apparatus that orchestrates the local pyrophosphate/inorganic phosphate ratio, and the lipid/protein composition of their lumen can act as a template for the minerals that develop there. In addition, phosphate transporters and annexins, which are thought to take part in the process of binding and Ca2+ transport into the lumen of the MV, are abundant in MVs.32–35
MVs vary from their progenitor cells due to a specific lipid composition. 36 The idea that osteoblasts produce MVs that include a high concentration of pyrophosphatase in addition to tissue-nonspecific alkaline phosphatase (TNAP) is central to the understanding of intramembranous ossification. Initial crystal formation occurs within the MVs as a result of calcium influx, which is thought to be caused by annexin-V's capacity to attach to calcium. 37 The crystals develop, break through the membrane of the MV, and disperse outside the MV boundary into the ECM.37–39 During the initial phases of mineralization, the MVs of both wild-type and deficient cartilage contained needle-shaped crystals of the apatitic mineral. In contrast to wild-type mice, TNAP-knockout mice showed fractured and more granular electron-dense mineral deposits (Fig. 5). 40
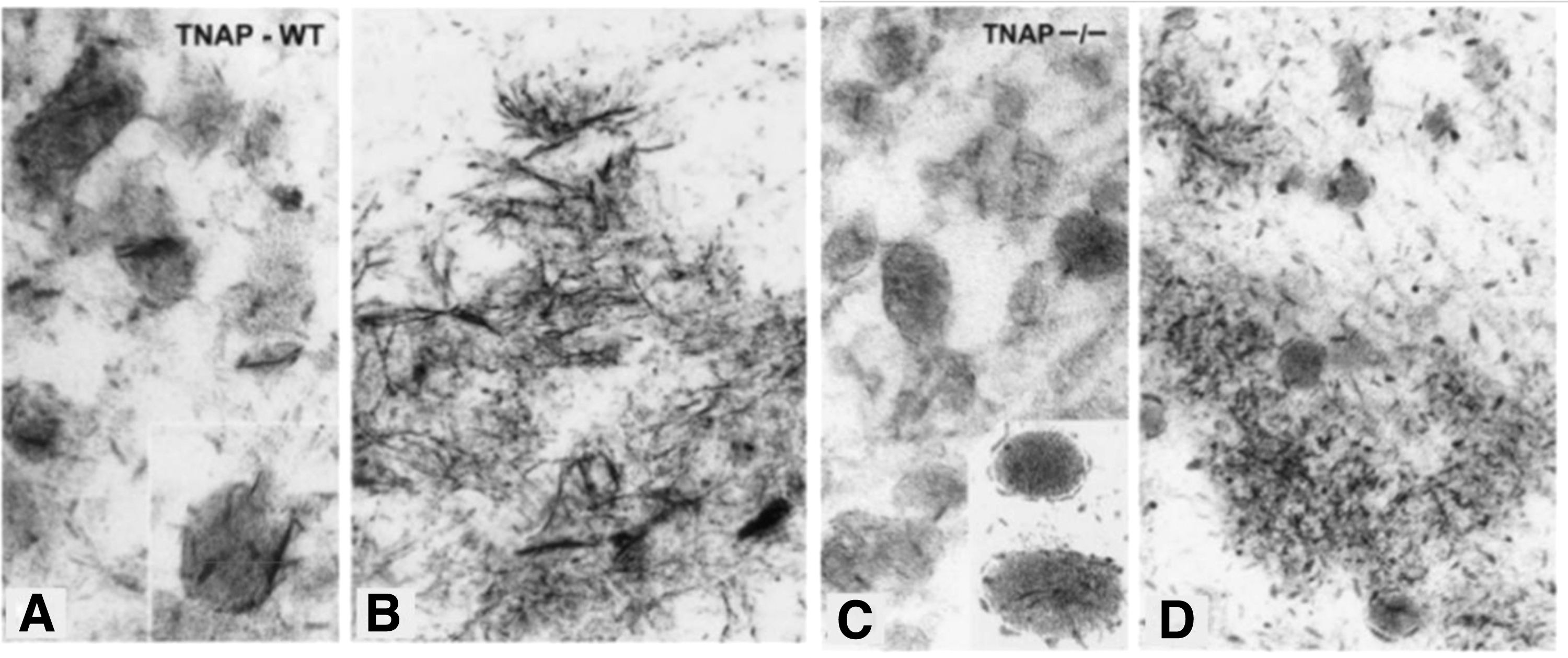
At the beginning of the mineralization process, TEM of TNAP wild-type growth plates
These findings demonstrate that MVs can regulate the formation of mineral nuclei and the growth of apatite crystals on collagen fibrils, regardless of the nature and origin of the mineralizing vesicles (Fig. 6). An intraluminal NC possesses a phospholipid-dependent method for the formation of calcium phosphate complexes. 41 The significance of researching the ways in which these vesicles regulate mineral nucleation is highlighted by mounting evidence that MVs are involved in collagen mineralization.
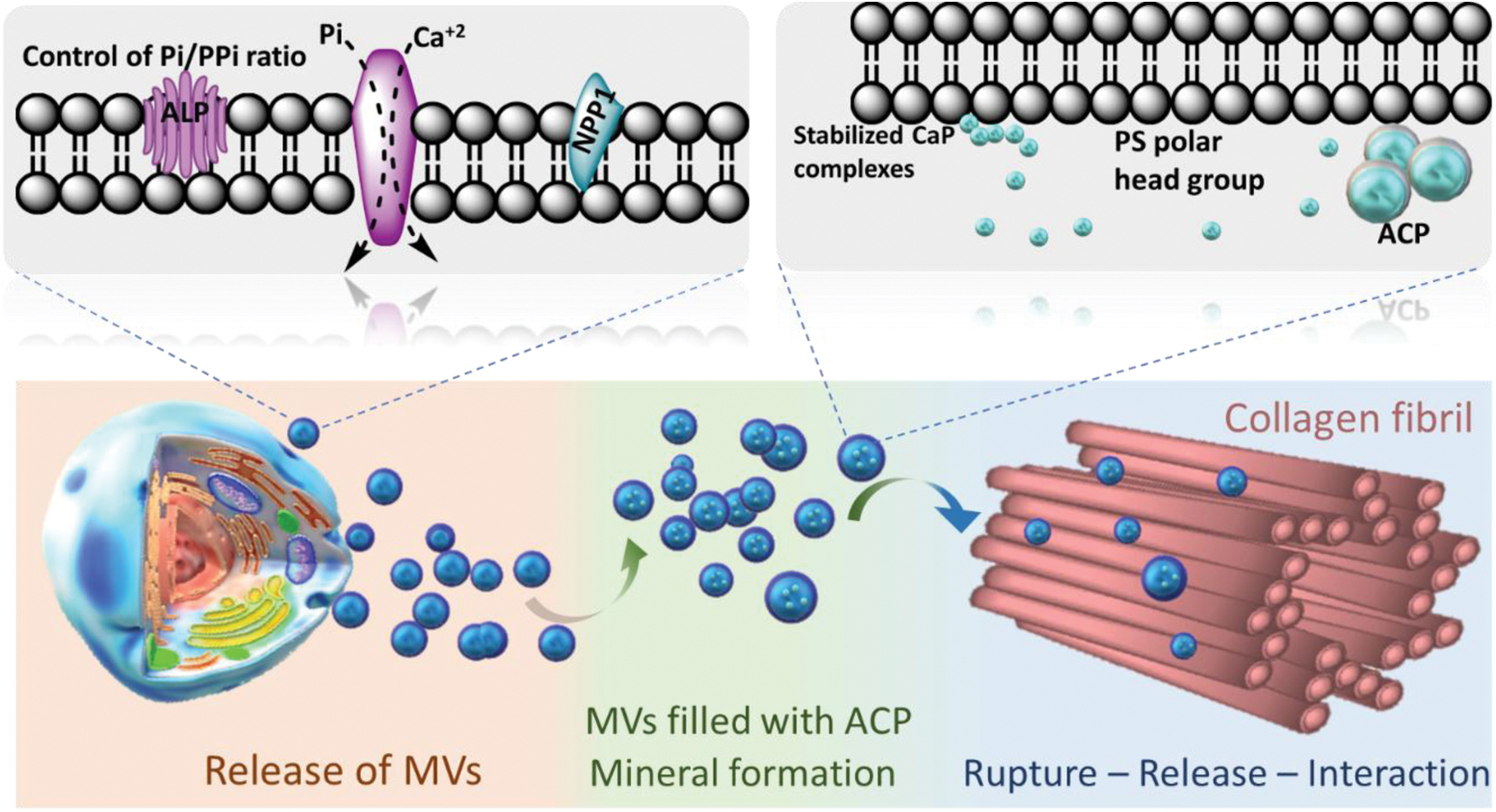
Role of MVs in the mineralization of collagen fibrils and the formation of calcium phosphate complexes. Cells that are capable of triggering mineralization secrete MVs with a metabolic apparatus that can regulate the ratio of phosphate to pyrophosphate. The inner membrane leaflet of these vesicles is rich in PS, which produces a strongly negatively charged interface for the nucleation and stability of calcium phosphate complexes. The very first mineral phase in the MV lumen, ACP, is created as a result of these complexes. ACP-filled vesicles may directly penetrate the collagen fibrils scaffold before crystallizing into platelet-like structures. ACP, amorphous calcium phosphate; PS, phosphatidylserine. Color images are available online.
NCs isolated from natural MVs were found to be made of ACP, serving as the major mineral component, and subunits of clusters of calcium and phosphate ions with a diameter of 1 nm. 41 MVs loaded with ACP could immediately enter the collagen fibrils that made up the scaffold and subsequently change into crystals resembling platelets. Similar structures (i.e., mineralized globules) have been observed in the collagen scaffold of zebrafish fin tails 42 and avian leg tendons. 43 This direct amorphous-apatite transformation has been proposed in vitro using confined polymer domains. 44
MVs may then break down and release their components to the mineralizing front, either as a result of mechanical stress or the activity of phospholipases. 45 Phospholipases are highly active enzymes in the growth plate. 46 This process may result in the release of phospholipid-mineral complexes, whether or not they are connected with proteins, and may make it possible for the biomineral precursor phase to infiltrate the gap area of collagen fibers by a mechanism analogous to that suggested for NCPs. 47 Alternately, these phospholipid-mineral complexes might engage with NCPs and subsequently be guided to the collagen matrix for mineralization after completing their interactions.
Mineralization Based on Other than MVs
In this study, the question arises as to whether the actual mineralization in vivo is mainly due to MVs or to lipids other than MVs. A recent study has shown that chondrocyte membrane nanofragments serve as nucleation sites for the creation of minerals during endochondral ossification. These cell membrane nanofragments are produced by chondrocyte rupture and maintain a linear structure rather than a vesicular structure owing to integrin-mediated collagen binding. In addition, synthetic cell membrane nanofragments can be used as raw materials for in vitro enhanced mineralization. 48
In our most recent study, we explored the process of early dentin development in the mouse first molars from a material science perspective. We determined the timing and location of the early dentin formation. Importantly, in predentin, which is at the forefront of dentin mineralization, the localization of collagen and phospholipids coincided; however, the presence of MVs was not observed. Interestingly, Annexin V accumulation was observed at this site (Fig. 7). 49 Annexin V is a marker of apoptosis and indicates cell membrane fragmentation. Many cells are prebound to collagen in this region through cellular focal adhesion systems without Ca deposition. In other words, mineralization did not originate from the secretion of MVs. This suggests that cell membranes (phospholipids), which are prebonded through focal adhesion systems, are fragmented by physical and chemical stimuli and remain on the collagen and that the remaining phospholipids induce mineralization.
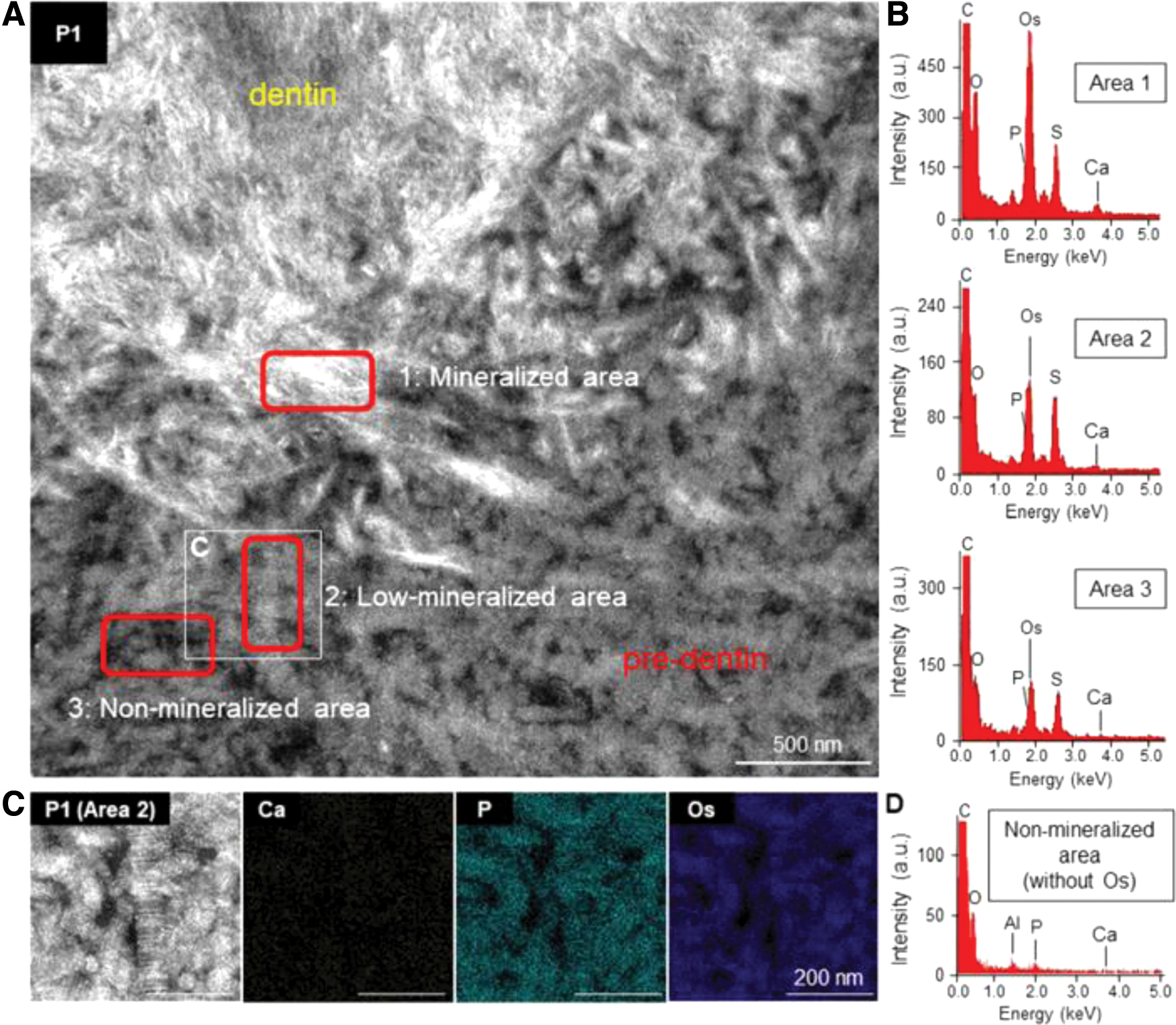
Elemental investigation of the dentin mineralization front.
These results led to the following hypotheses: (1) cell fragments composed of phospholipids, even if they are not MVs, serve as sites for mineralization. (2) Fragmented phospholipids are generated not only by MV secretion but also by mechanical effects, including cell rupture and tearing. (3) The idea that phospholipids are involved in most mineralization in vivo is supported by the fact that almost no mineralization occurs in collagen, which is abundant in the body. (4) PS, a component of the inner layer of the cell membrane, promotes the induction of mineralization, although mineralization does not generally occur on the cell surface. It can be said that most cell destruction occurring at the terminal stage of cells is a trigger for biomineralization. This idea is supported by the ectopic mineralization observed in diseased tissues.
Conclusion and Future Perspective
The primary focus of the present article was to elucidate the importance of lipids in the biomineralization process. Understanding mineralization based on lipids implies that the same mechanism also functions in mineralization obtained from bacteria, such as calculus, 50 and in the mineralization of marine species, such as corals. 12 This is because lipids play a central role in mineralization. It is also crucial to note that the lipids engaged in this mineralization process are released from the cells and obtained from the components that make up the cells. This is another significant aspect of this work. It is essential to view the components of cells as substances that promote mineralization rather than viewing cells themselves as entities that govern mineralization, as this is the only way to properly understand this process. Biomineralization is triggered by cell death or dead cells themselves. For greater comprehension, engaging in interdisciplinary work that combines cell biology, molecular biology, and materials science is essential.5,51,52
In addition, biological mineralization involves more than just protein and lipid components. Water is one of the most important contributors to the microenvironment. It has been proposed that the denaturation of collagen may vary depending on the quantity of water present. 49 Collagen that has been denaturized is associated with the adherence of minerals related to the expansion of bone. 53 In addition, it plays a significant role in the regulation of this aqueous environment. In the natural state of collagen, proteoglycans, such as decorin, are localized in the gaps between collagen fibrils to serve as space maintainers. Carbohydrate–lipid interactions in this area may play a key role in the localization of ions and preservation of the energy state necessary for lipid-mediated mineralization.54–56 In addition, changes in surface hydrophilicity and hydrophobicity associated with collagen denaturation will likely be the major factors regulating lipid penetration. Thus, identifying and understanding the roles of other organic molecules are crucial for further understanding biomineralization.
Footnotes
Authors' Contributions
Conceptualization, B.N. and T.M.; writing/original draft preparation, B.N.; writing/review and editing, B.N., M.O., and T.M.
Disclosure Statement
No competing financial interests exist.
Funding Information
The authors received the KAKENHI (JP22F22413) from Japan Society for the Promotion of Science and by CREST (Grant No.: JPMJCR22L5) from Japan Science and Technology Agency for support.