
Abstract
Cartilage restoration continues to present a tremendous clinical challenge due to its nonvascular nature. Many studies have demonstrated that chondrogenesis of progenitor cells can be achieved in vitro by manual dose of growth factors; however, it remains a vital difficulty in feeding growth factors to implanted therapeutic cells in vivo. Herein, we constructed recombinant adenovirus encoding human transforming growth factor-β3 (hTGF-β3) and practiced it in rat bone marrow–derived mesenchymal stromal cells and articular chondrocytes for cartilage regeneration. Optimal viable transduction and transgenic hTGF-β3 production were achieved; consequently, positive expression of cartilage marker-collagen type II was enabled in the infected progenitors. We thus conclude that recombinant adenovirus encoding TGF-β3 gene has been successfully established and validated for cartilage tissue engineering applications.
Introduction
In the recent decade, novel strategies with tissue engineering have evolved from bench-side exploration to clinical applications. With the development of gene delivery to chondrocytic/progenitor therapeutic cells such as mesenchymal stem cells, treating disorders of articular cartilage by tissue engineering has borne much promise. For this purpose, various growth factor genes have been involved,2–11 among which members of transforming growth factor-β (TGF-β) have been acknowledged most effective to induce chondrogenesis in vitro.2–4
Three isoforms of TGF-β (TGF-β1, TGF-β2, and TGF-β3) have been identified in mammalian species till today. TGF-β1 was the first to be discovered and most studied. However, TGF-β1 and TGF-β2 can lead to a fibrotic scarring response, while TGF-β3 elicits a scar-free or regenerative healing response.12,13 With a better understanding of the biological functions of TGF-β3, researchers have been attempting to enable in vitro chondrogenesis by dosing the progenitor cells with recombinant TGF-β3 protein in culture medium.14–18
However, more sophisticated investigations and practical applications are required before the growth factors are used in clinic. Further, how to fulfill and administrate in situ dose of this growth factor–based “drug” to the transplanted (progenitor) therapeutic cells in vivo still remains a significant challenge for both clinicians and tissue engineers. Since growth factor has a very short half-life, it is necessary and important to establish a method to provide sufficient, sustained, and effective supply of growth factors for desired cell development.
Toward this end, we intend to explore an effective tool that can generate growth factor TGF-β3 from the therapeutic cells themselves for the use of their own. Recombinant adenovirus encoding TGF-β3 (Ad.TGF-β3) is herein constructed to infect candidate therapeutic cells, rat bone marrow–derived mesenchymal stromal cells (BMSCs), and chondrocytes, for cartilage regeneration. In this proposal, the growth factor genes together with the viral vectors are delivered into target cells in vitro instead of being directly applied to recipients per se, and the viral immunogen would have been metabolically removed from the cells during the period of in vitro culture; thus; the posttransplanting safety concerns could be minimized.
Materials and Methods
Construction of the recombinant adenoviral vector encoding human TGF-β3
The target gene was obtained from human transforming growth factor-β 3 (hTGF-β3)-pCMV6-XL5 plasmid DNA (Origene Technologies, Rockville, MD) by PCR. The sequences of PCR primers were shown in Table 1. Both the PCR product and the shuttle plasmid were digested with SalI and PstI restriction enzymes (New England Biolabs, Ipswich, MA). The digestion products were then ligated using T4 DNA Ligase (Invitrogen, Carlsbad, CA) at 16°C overnight. The ligation product was prepared from DH5α, Escherichia coli cells, purified with an QIAfilter plasmid extraction kit (Qiagen, Hilden, Germany) and verified by digestion analysis and DNA sequencing.
The resultant shuttle vector was then used to transfer hTGF-β3 to pLP-Adeno-X ViraTrak acceptor vector, which contained an E1, E3-deleted, replication-deficient, type 5 adenovirus genome (Ad5), by Cre-loxp site-specific recombination (Adeno-X™ ViraTrak™ ZsGreen1 Expression System 2; Clontech Laboratories, Mountain View, CA). Recombinants were isolated and amplified by transforming DH10B E. coli cells with electroporation and screened against donor vector pDNR-CMV and nontransfected cells by selection on medium containing both chloramphenicol and sucrose. Then, the purified recombinant plasmid was confirmed by XhoI restriction analysis.
Producing, amplifying, and purifying recombinant adenovirus with hTGF-β3
Lower-passage HEK 293 cells (ATCC, Manassas, VA) were transfected with PacI-digested recombinant plasmid using lipofectamine to produce infectious adenovirus. Approximately 5–7 days after transfection, 50–90% of the cells detached from the plate due to cytopathic effect (CPE). Then, recombinant adenovirus was harvested by lysing cells with three consecutive freeze–thaw cycles. This stock was named “primary amplification stock” and stored at −80°C.
The expression of hTGF-β3 was characterized by Western blots using rabbit anti-human polyclonal TGF-β3 (Abcam, Cambridge, UK) as primary antibodies and goat anti-rabbit IgG-horseradish peroxidase (IgG-HRP) (Santa Cruz Biotechnology, Santa Cruz, CA) as secondary antibody. Reaction products of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (Imgenex, San Diego, CA) were used to normalize the intensities between lanes. Nontransfected HEK 293 cells were used as negative controls.
This virus stock was then titered (Adeno-X™ Rapid Titer Kit; Clontech) and subsequently used to prepare a high-titer stock. The high-titer virus stock was purified (Adeno-X™ Virus Purification Kit; Clontech) and titered before being used to infect target cells. Five batches of purified recombinant adenoviruses have been used in the following experiments.
Isolation of rat BMSCs and chondrocytes
A 6-week-old male Wistar rat was acclimatized for 1 week at a local animal facility. The rat was anaesthetized with CO2 inhalation. After sterilized in 75% ethanol for 5 min, the anaesthetized rat was euthanized with cervical dislocation. Then, the femur and tibia were dissected, and the attached tissues were removed. The cartilage was removed for isolating chondrocytes later, and then the bone marrow cavity was rinsed with Dulbecco's modified Eagle's medium (DMEM; Gibco, Carlsbad, CA) + 10% fetal calf serum (FBS; Gibco) repeatedly to collect the BMSCs. The cells were harvested by centrifugation at 1000 rpm for 5–7 min. After being resuspended with MSCGM (Cambrex Bio Science, Walkersville, MD), the cells were seeded into 75 cm2 flasks with a density of about 100,000 cells/cm2. The cultures were maintained at 37°C in an atmosphere of 5% CO2. The medium was changed every 3 days.
The cartilage was cut into small pieces with size about half of a rice grain, followed by digestion with 1 mg/mL collagenase type II (250 units/mg; Worthington, MN) with gentle rotation at 60 rpm at 37°C in 5% CO2 atmosphere for 12–14 h. Isolated chondrocytes were cultured in DMEM containing 10% FBS, 100 μg/mL streptomycin, 100 units/mL penicillin, and essential amino acids. The medium was changed every 3 days. Four batches of rat BMSCs and chondrocytes have been used for the following experiments.
Adenoviral transduction of rat BMSCs and chondrocytes
Passage 5 rat BMSCs and passage 3 rat chondrocytes were, respectively, seeded into six-well plates with the density of 2 × 105 cells/well 12–24 h before infection. Then, the cells were incubated in 1 mL of serum-free DMEM with various doses of Ad.TGF-β3 ranging from 10 to 1000 multiplicity of infection (MOI, defined as pfu/cell) for 2 h at 37°C in 5% CO2. Control cultures remained nontransduced. Following infection, the culture fluids were aspirated and replaced with 2 mL DMEM with 10% FBS.
Flow cytometry analysis
The percentage of GFP-positive cells and cell viability by exclusion of propidium iodide (50 μg/mL, BD) were investigated at 48 h after infection by using a FACSCalibur flow cytometer and CellQuest software (BD, Heidelberg, Germany). The percentage of cells emitting fluorescence of each sample was measured three times. The percentage of GFP-positive cells was determined by counting 10,000 cells in each measurement.
Quantitative analysis of hTGF-β3 expression by specific ELISA
Passage 5 rat BMSCs and passage 3 rat chondrocytes were infected with Ad.TGF-β3 at MOI 100 and then encapsulated into 2% agarose hydrogels (1 × 107 cells/mL). Rat BMSCs or chondrocyte-seeded hydrogels were cultured in complete growth media in 24-well plate. Conditioned medium was collected at various time points as indicated in the figure and stored at −80°C for further analysis by specific ELISA (R&D Systems, Minneapolis, MN). Media from uninfected rat BMSCs and uninfected chondrocyte-seeded hydrogels were collected as negative controls.
We also detected the dynamic changes of recombinant TGF-β3 concentrations in culture medium. In this group, uninfected chondrocyte-seeded hydrogels were cultured in the medium containing 10 ng/mL recombinant TGF-β3 (Origene Technologies, Rockville, MD). The medium was changed every 3 days.
Triplicates were performed for each time point. Briefly, before the experiment, a 96-well microplate was coated with capture antibody. Latent hTGF-β3 in the samples was activated with hydrochloric acid. The standards with twofold serial dilutions and the pretreated samples were added onto a precoated 96-well plate, followed by incubation for 2 h at room temperature. After washing with phosphate-buffered saline + 0.05% Tween 20 for three times, the plate was incubated with detection antibody, streptavidin-HRP, and substrate solution in sequence. Finally, the absorbance in each well was determined using a microplate reader set to 450 nm.
Reverse-transcription PCR (RT-PCR)
Total RNA was isolated from rat BMSCs before infection and 2 h, 2 days, and 3 days postinfection using TRIZOL® Reagent (Invitrogen). For cDNA synthesis, 1 μg of total RNA was reverse transcribed using ™ First-Strand Synthesis System (Invitrogen) for RT-PCR. The reaction product containing approximately 400 ng of cDNA was used as a template for PCR amplification. Collagen type II was used as a marker for chondrocytes. The PCR conditions were as follows: denaturation at 94°C for 30 s, annealing at 50°C for 30 s, and extension at 72°C for 30 s. Thirty cycles were performed. PCR products were visualized on 1% agarose gels. Differences in staining intensities between lanes were normalized using β-actin reaction products.
Statistical analysis
All data were collected from various batches of viruses and cells; therefore, the statistical analysis covers the batch-wise variation. Comparison among different groups for flow cytometry analysis was made by one-way ANOVA. Student's t-test was used for comparing the ELISA data among the groups at different time points. Differences between experimental groups were considered to be significant at p < 0.05.
Results
Construction of recombinant adenovirus encoding hTGF-β3
The DNA sequencing indicated that the sequence of inserted hTGF-β3 in shuttle plasmid get 99% match with hTGF-β3 sequence from the NCBI blast website. A point mutation—T in the place of C—has occurred, which is part of the Codon UGC. This mutation has no effect on the expression of TGF-β3 protein as the Codon UGC codes for cysteine, which is also coded by UGU or TGT (in terms of DNA sequence). It was also verified by Western blots later.
After digestion of recombinant adenoviral vector with XhoI, the expected bands 14.5, 9.7, 5.6, 4.2, 2.5, 1.4, and 0.6 bp were seen for the recombinant adenoviral vector with both TGF-β3 and a green fluorescent gene, confirming successful recombination and transfer of hTGF-β3 from shuttle plasmid to acceptor vector (Fig. 1).
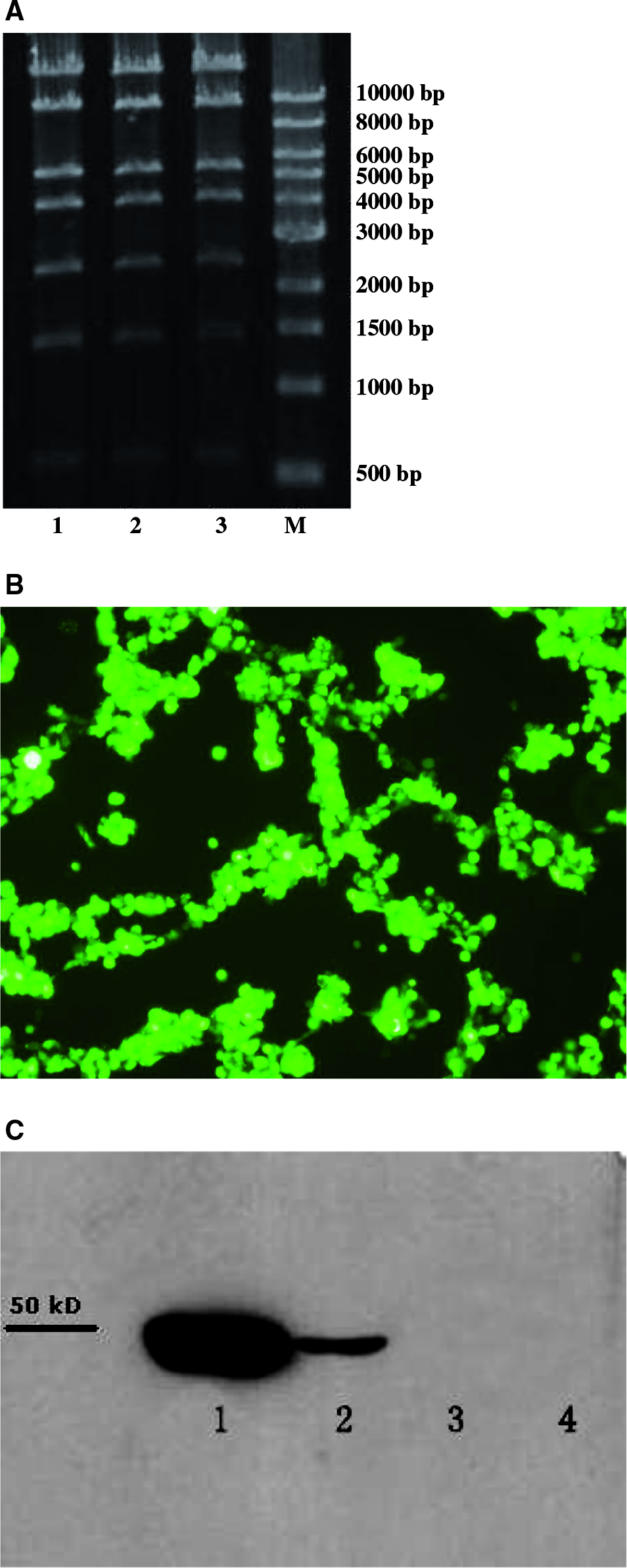
Construction of recombinant adenovirus encoding hTGF-β3. (
The HEK 293 cells were transfected with recombinant adenoviral vector when they reached 50–70% confluency. Two days later, expression of the Vira Trak fluorescent protein was checked by viewing the cells under fluorescent microscope. Remarkable fluorescence was observed in the transfected HEK 293 cells under microscope (Fig. 2).
Flow cytometry analysis. (
When CPE appeared, the medium was collected to precipitate protein (labeled as Pro 1) by ethanol, and cells were used for isolating protein (labeled as Pro 2) with cell lysis buffer. Expression of hTGF-β3 was verified by Western blots. As shown in Figure 3, both Pro 1 and Pro 2 collected from transfected HEK 293 cells had the bands around 47 KD, indicating the existence of hTGF-β3, while there was no band around 47 KD in those of negative control.
Dynamic changes of hTGF-β3 concentrations by ELISA.
All the results above confirmed that recombinant adenovirus encoding hTGF-β3 has been constructed successfully.
Transduction efficiencies and cell viabilities of infected rat BMSCs and chondrocytes with Ad.TGF-β3
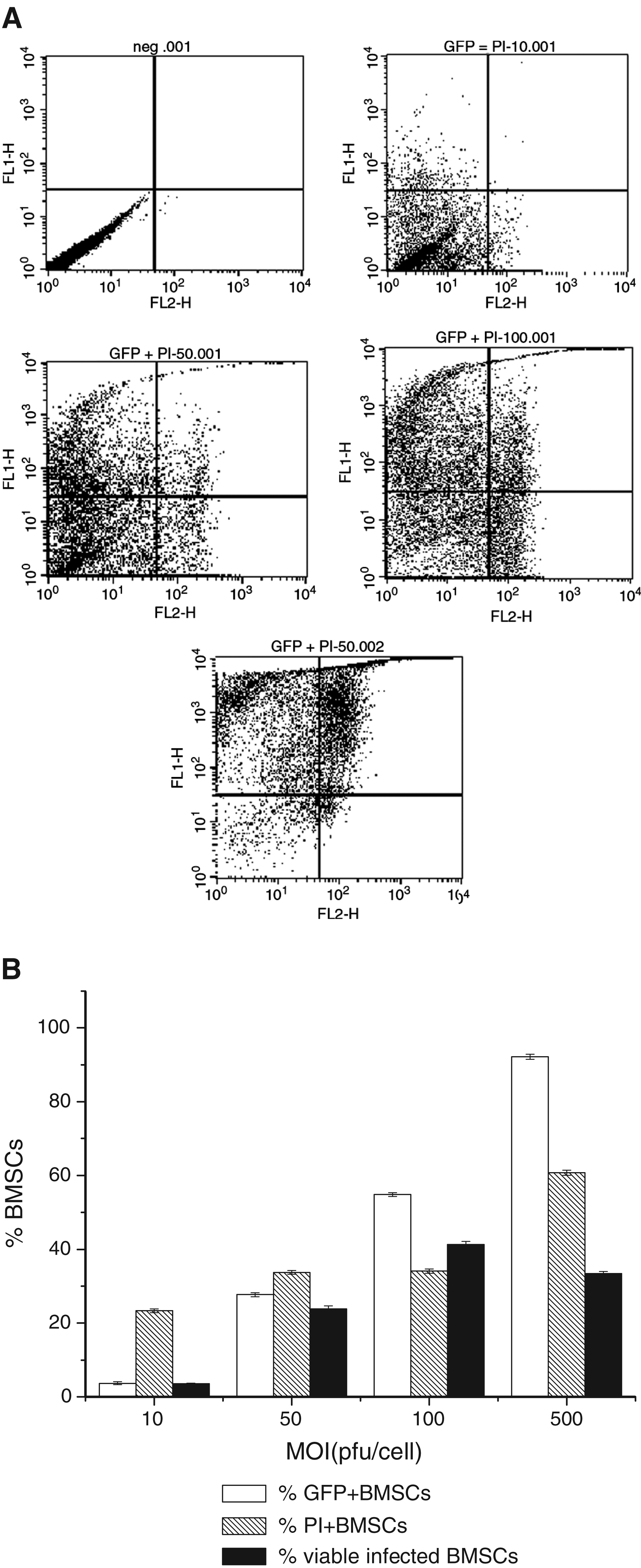
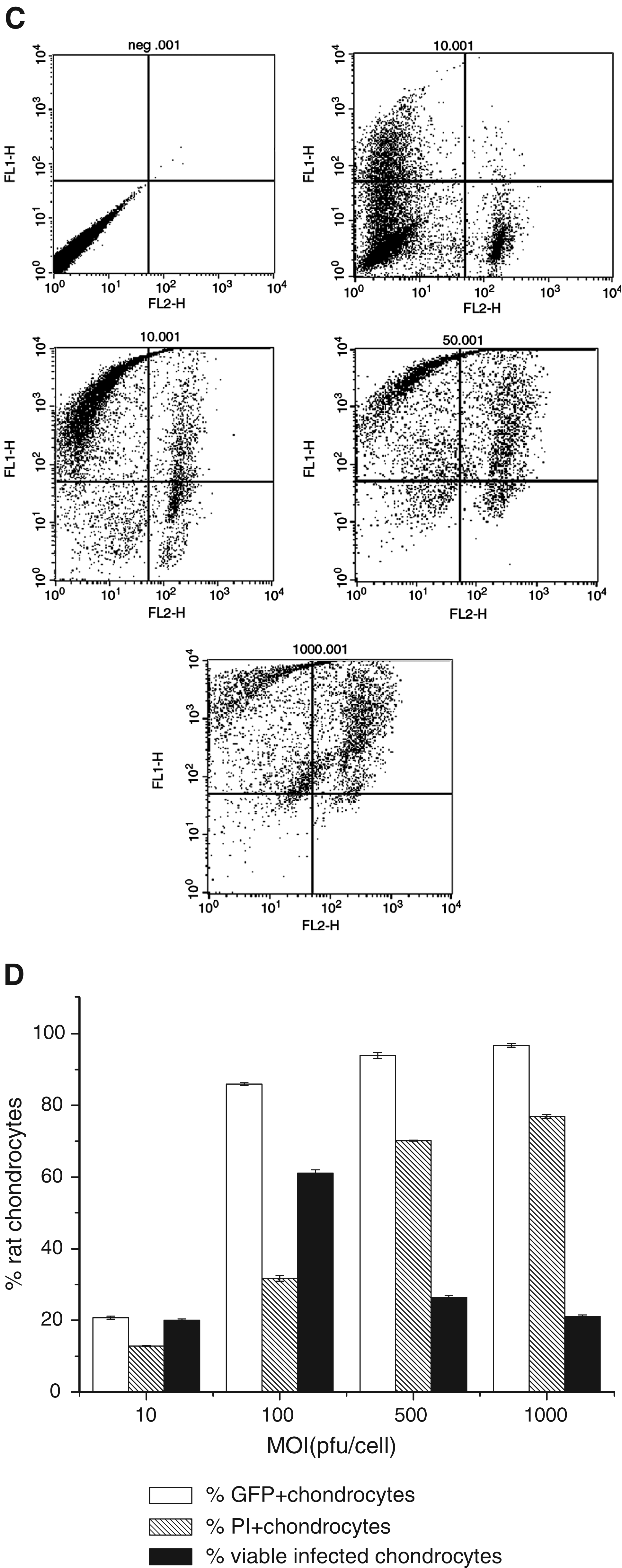
In order to search for the optimal viral titer for rat BMSC and chondrocyte infection, we infected passage 5 rat BMSCs and passage 3 chondrocytes in monolayer culture with increasing amounts of Ad.TGF-β3.
As shown in Figure 2A–D, the percentage of GFP-positive cells exhibited a viral dose–dependent increase, and the propidium iodide–stained cells (dead cells) also increased when more virus infected rat BMSCs or chondrocytes. Both BMSCs and chondrocytes exhibited higher transduction efficiency and cell viability at MOI of 100 pfu/cell (Fig. 2B, D). Based on these results, an adenoviral dose of 100 pfu/cell was chosen for subsequent experiments.
Dynamic expression of hTGF-β3 by ELISA analysis
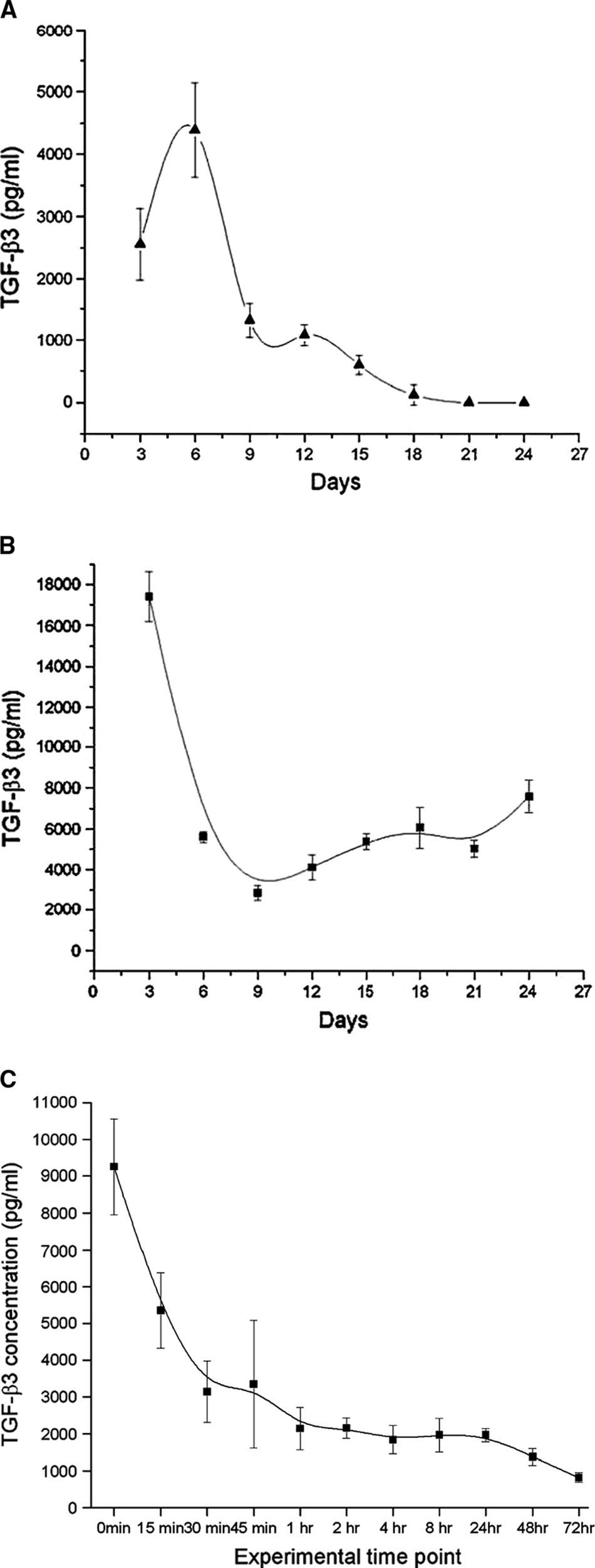
TGF-β3 could be detected neither in the medium of uninfected rat BMSC–seeded hydrogels nor in that of uninfected chondrocyte-seeded hydrogels (data not shown).
As shown in Figure 3A, the level of hTGF-β3 secreted by infected rat BMSCs fluctuated between 2.5 and 4.5 ng/mL in the first week. Later, hTGF-β3 expression decreased rapidly, even to less than 1 ng/mL, and maintained at low level for about 10 days. Infected rat chondrocyte-seeded hydrogels secreted much more hTGF-β3 than infected rat BMSCs did as a whole (Fig. 3B). Transgenic hTGF-β3 secreted by infected chondrocytes exhibited higher concentration in an early stage. Then, the concentrations of hTGF-β3 were stable at around 6 ng/mL for about 2 weeks.
By contrast, the recombinant TGF-β3 added manually degraded in the medium amazingly since its concentration dropped rapidly from nearly 10 ng/mL to around 2 ng/mL in only 1 h. Then, the recombinant TGF-β3 levels continued to decrease although the attenuation was not as sharp as in the first hour (Fig. 3C).
In negative controls, TGF-β3 expression could be detected neither in noninfected BMSCs nor in noninfected chondrocytes.
Chondrogenic differentiation of rat BMSCs following adenoviral-mediated gene transfer of hTGF-β3
As shown in Figure 4, there was no collagen type II expression in normal rat BMSCs, while the infected rat BMSCs expressed mRNA for collagen type II at 2 and 3 days after infection. For noninfected control, there was β-actin expression without collagen type II expression (data not shown). These preliminary results indicate that rat BMSCs can differentiate into chondrocytes with the presence of transgenic TGF-β3.
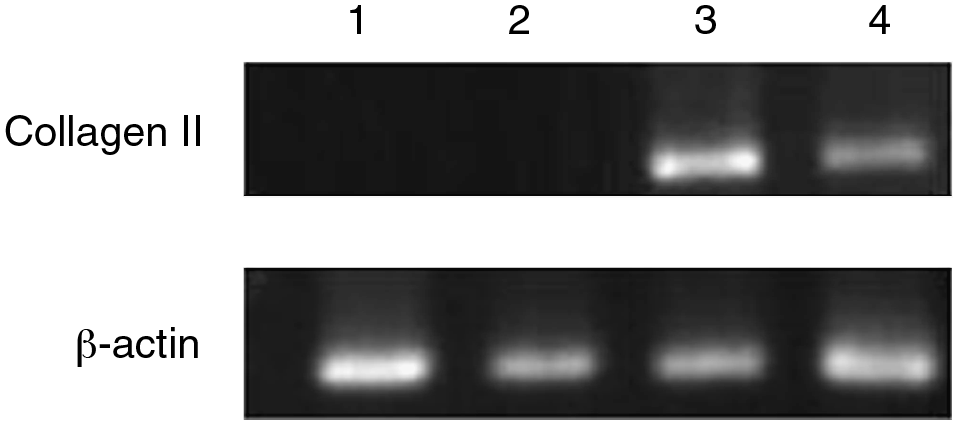
Expression of collagen type II in infected rat BMSCs. RT-PCR products of RNA isolated before infection and 2 h, 2 days, and 3 days postinfection were shown in lanes 1–4, respectively. Expression of collagen type II in infected rat BMSCs was significant at 2 and 3 days postinfection.
Discussion
Growth factors have long been recognized to promote cell proliferation, maturation, and differentiation. Candidate growth factors that induce chondrogenesis include members of the TGF-β superfamily (which includes bone morphogenetic proteins, BMPs),2–8 members of the fibroblast growth factor (FGF) family, 9 and insulin-like growth factor (IGF)-1.10,11 Apart from growth factors, genes for transcription factors, such as Sox-5, Sox-6, and Sox-9,19–21 matrix proteins, 22 and inhibitors of articular cartilage degeneration 23 have also been employed in cartilage regeneration.
Members of the TGF-β superfamily have been shown to play a major role in bone and cartilage development. TGF-β signaling stimulates proliferation and inhibits terminal differentiation of chondrocytes during chondrogenesis. 24 Several studies have shown that TGF isoforms differ in their effects on chondroprogenitor and other cells. 25 TGF-β1 has shown anti-inflammatory properties in inflamed joints, 26 while TGF-β2 mediates hypertrophic differentiation of chondrocytes by regulating Indian hedgehog (Ihh) and parathyroid growth hormone (PTHrP) expression. 27 Both TGF-β1 and TGF-β2 have been reported to have an osteoinductive potential,28,29 while TGF-β3 gets upregulated during dedifferentiation of human chondrocytes in cell culture. 30 Na et al. have also reported that TGF-β3 can induce differentiation and cartilage-specific ECM production in chondrocytes in hydrogel constructs in vivo. 31 Therefore, compared with TGF-β1 and TGF-β2, TGF-β3 may play a more important role in chondrogenesis. As shown in our studies, collagen type II gene was expressed in transgenic BMSCs 2 days postinfection, which indicates that the progenitor cells have started to demonstrate chondrogenic commitment in the presence of TGF-β3.
Although in vitro chondrogenesis has been achieved by dosing the progenitor cells with recombinant TGF-β3 protein in culture medium,14–18 it is difficult to feed growth factors to implanted therapeutic cells in vivo. Another problem also need to be addressed that there is insufficient local stimulation of the implanted cells by the protein factors necessary to drive differentiation in vivo. 32 Given that gene-based growth factor supply works in an endogenous way that fulfills the in vivo requirement of clinical applications, gene transfer approaches are more favored and have been applied in regenerative researches. Till now, less is known about the details of progenitor cells to undergo chondrogenesis when TGF-β3 is delivered via gene transfer.
Gene delivery systems mainly include nonviral and viral systems. Since viral gene transfer method exhibits less (acute) toxicity and relatively high transduction rate, 33 viral delivery systems thus have attracted more attention and have been used in many clinical trials. Adenovirus is the most widely used gene therapy vector due to its ability to infect a broad range of human cells, including both dividing and nondividing cells,34,35 without integrating into the host genome. 36 Transduction efficiencies in stem cells reached up to 50% with adenoviral vectors. 37 Our results indicate that at MOI 100 pfu/cell the transduction efficiency in BMSCs can reach 55%, while in chondrocytes, 86%.
However, immune responses need to be addressed before using adenovirus in vivo. The host immune response induced by adenoviral vectors is strong enough to attenuate the efficacy of adenovirus-based approaches. Molinier-Frenkel et al. 38 demonstrate that it is capsid components from viral input that serve as targets for vector-specific cytotoxic T lymphocyte recognition. Contrastively, with our strategy the target cells were infected by in vitro gene delivery approaches and the gene-delivered cells were kept culturing for some time before being transplanted in vivo. In other words, recombinant adenovirus encoding TGF-β3 is not injected directly in vivo but delivered into target cells in vitro before future use. As soon as capsids specifically dock to the nuclear pore complex and release the transgene into nucleus, the final disassembly of the viral capsids ensues. 39 Therefore, the existence and effects of capsids are not of concern, and the transgenic cells can be used safely in regenerative medicine.
For cartilage regeneration, the gene-delivered cells are to be seeded into engineered constructs and transplanted to articular defects. The regenerative functions of the gene-enhanced implants can be significantly improved with the sustained, localized, and periodically over-expressed produc-tion of genetic products—the therapeutic growth factors. Our results indicate that transgenic BMSCs and chondrocytes secrete hTGF-β3 constantly in a couple of weeks. In hydrogel scaffold, infected rat chondrocytes secreted much more hTGF-β3 than BMSCs did as a whole. The quantities of TGF-β3 secreted by infected chondrocytes were around 4–8 ng/mL from 10 days postinfection, which was comparable to the amount of the recombinant TGF-β3 added manually. 40 However, unlike the manual dosing of TGF-β3, which has extremely short pharmacological half-life, the release of TGF-β3 by infected cells could be maintained for weeks.
In conclusion, recombinant adenovirus encoding TGF-β3 gene has been successfully constructed and employed to infect rat chondrocytes and BMSCs with high transduction efficiency. The transgene has been expressed from the infected cells in multiple weeks, which could further catalyze the positive expression of downstream cartilage markers such as type II collagen.
Footnotes
Acknowledgments
This research was financially supported by Grant ARC 10/16, Ministry of Education, Singapore, and Startup Grant, College of Engineering, Nanyang Technological University, Singapore.