
Abstract
Adult tissues contain stem cells that can transdifferentiate into other cell lineages besides forming differentiated cells of their own tissue of origin. However, human adult skin-derived stem cells have a very low efficiency. Here we established a novel culture system involving bone morphogenetic protein-4 and a floating culture system with sphere-producing medium that can enrich adult stem-cell populations in vitro. Adult stem cells were isolated from useless human scar tissue. Like mesenchymal stem cells, cultured human scar tissue-derived stem cells (hSTSCs) altered their morphology and significantly increased the number of Nestin-positive cells in proportion to the alkaline phosphatase–positive cell ratio. Moreover, the expression of the pluripotency regulator Oct-4 and its target transcripts, Sox-2, c-kit, and Rex-1, was also stimulated by this culture system. Differentiation of neurogenic progenitor cells using basic fibroblast growth factor and Neurogen 2 was successfully performed in vitro more rapidly than previous reports. Neuronal differentiation results showed that our hSTSCs expressed marker of neurogenic genes, such as glial fibrillary acid protein, neural cell adhesion molecules, neuron filament-M, and microtubule-associated protein 2. These results suggest that bone morphogenetic protein-4 and the floating culture system with sphere-producing medium induced significant proliferation of hSTSCs and mediated reprogramming of the cells from adult somatic tissue into precursor state to some degree. It is thought that this new culture system might be a simple, effective, and easily manageable process for regenerative tissue repair and autotransplantation.
Introduction
Materials and Methods
Human scar tissue cultures
Human scar tissue was obtained from four female donors aged 35–45 years following second cesarean sections. These procedures were performed in compliance with the local ethical guidelines and have been approved by the Institutional Review Board (IRB) of the Kyung-Hee University Medical Hospital in Korea (Approve no.: KMC IRB ER0603-01). To isolate the hSTSCs, scar samples (composed of both dermis and epidermis) were dissected, cut into small pieces, and incubated with growth medium (GM) consisting of Dulbecco's modified Eagle's medium (DMEM; Gibco BRL, Grand Island, NY) containing 1 mM glutamine (Invitrogen, Carlsbad, CA), 1% (v/v) minimal essential medium (MEM) nonessential amino acids (Invitrogen), 10 μM 2-mercaptoethanol (Sigma-Aldrich, St. Louis, MO), 10 mg/mL penicillin/streptomycin glutamine (Invitrogen), and 20% ESC-qualified fetal bovine serum (FBS; Hyclone, Logan, UT) in a humidified atmosphere of 5% CO2/95% air at 37°C. To ensure a sufficient number of cells, each tissue piece was carefully removed from the dish with fine forceps after 7–10 days of primary culture. The cultures were subsequently treated with 0.02% trypsin–ethylenediaminetetraacetic acid (EDTA) (Invitrogen) for 5 min at 37°C and then dissociated into single cells by pipetting. Then 1 × 106 viable cells/mL were seeded onto 100-mm bacteriological culture dishes with GM for 5 days. Cumulative population doubling at each subculture was calculated by the formula 2X = NH/NI, where NI = the inoculum cell number, NH = the cell harvest number at confluence (>80%), and X = population doublings. 43 The population doubling increase that was calculated was then added to the previous population doubling level to yield the cumulative population doubling level.
BMP-4 treatment and analysis of endogenous alkaline phosphatase activity
To determine the optimal concentration of BMP-4 treatment, various concentrations (0, 1, 10, 20, 30, 50, and 100 ng/mL) of BMP-4 (R&D Systems, Minneapolis, MN) were used to evaluate the differences in response. Two types of cultures were prepared in 12-well cell culture plates, that is, low-density and high-density cell cultures. Cells were cultured in the GM described above, and various concentrations of BMP-4 were added. Endogenous alkaline phosphatase (AP) activity was assessed using the Sigma 86C-1 kit according to the manufacturer's instructions to ensure the presence of stem or stem-like cells.
Sphere-like organoid body formation
The viability of the isolated cells was >90% as determined by trypan blue staining. Cell numbers were enumerated using a Coulter counter. Isolated dermal fibroblast cells were washed with basal medium (DMEM + 10% defined FBS) and resuspended in sphere-producing medium (SPM) consisting of DMEM/F12 (Invitrogen) with 20% knockout serum replacement (KSR) (Invitrogen), supplemented with 30 ng/mL BMP-4 and seeded with a density of 5 × 105 cells/mL in 100-mm bacteriological culture dishes, and then cultured in 5% CO2 at 37°C. After a 24-h treatment, the generated sphere-like organoid bodies (OBs) were washed with culture medium and harvested for immunofluorescence studies and molecular studies or were dissociated and plated for differentiation.
Cell differentiation
To induce neuronal differentiation, cells were mechanically dissociated by 0.05% trypsin–EDTA treatment for 5 min at 37°C, washed once with basal medium, and resuspended in neuronal differentiation medium, which contained DMEM/F12 (Invitrogen) in a 3:1 ratio with 10% FBS (HyClone), 10 mg/mL nerve growth factor, (Invitrogen) and antibiotics (100 U/mL penicillin G and 25 mg/mL gentamycin). Cells were plated at 1 × 106 cells/cm2 onto the poly-D-lysine (Sigma)–coated culture dishes or cover slips. Half of the medium was replaced by a fresh differentiation medium three times a week.
Immunocytochemistry
The hSTSCs and OBs were washed with Dulbecco's phosphate-buffered saline (Invitrogen) and cultured on glass slides. These hSTSCs and OBs were fixed with 4% paraformaldehyde (Sigma) and incubated with appropriate dilutions of antibodies against the following stem-cell proteins or neurotransmitters were used for immunostaining: Oct-4 (ESC marker sample kit; Chemicon International, Temecula, CA), stage-specific embryonic antigen-4 (SSEA-4) (ESC marker sample kit; Chemicon International), nestin (neural stem-cell marker characterization kit; Chemicon International, Temecula, CA), neuron-specific class III beta-tubulin (TuJ1) (neural stem-cellmarker characterization kit; Chemicon International), glial fibrillary acidic protein (GFAP) (neural stem-cell marker characterization kit; Chemicon International), and CD90 (Abcam, Cambridge, CA). The hSTSCs were incubated in 1:500 normal goat serum (Vector Laboratories, Burlingame, CA) for 1 h to block nonspecific binding. Primary antibodies were localized with fluorescein isothiocyanate–conjugated immunoglobulin G or rhodamine-conjugated immunoglobulin G second antibody, mounted on Vectashield mounting medium containing 1.5 μg/mL 4′,6-diamidino-2-phenylindole (Vector Laboratories), and then observed with an Olympus BX 61 fluorescence microscope (Tokyo, Japan).
Preparation of RNA and reverse transcriptase-polymerase chain reaction
RNA was extracted using RNeasy extraction kits (Qiagen) with on-column DNase digestion, according to the manufacturer's recommendations. The cDNA was synthesized by RT-Premix (AccuPower RT-Premix; Bioneer, Daejon, Korea). cDNA amplification was performed in a total volume of 20 μL using polymerase chain reaction (PCR) premix (AccuPower®, PCR-Premix; Bioneer) according to the manufacturer's instructions. Glyceraldehyde 3-phosphate dehydrogenase was used as an internal control.
Western blot analysis
CellS were lysed with ice-cold lysis buffer (150 mM NaCl, 50 mM Tris–HCl, pH 7.4, 2 mM EDTA, 1% Nonidet P-40, 10 mM NaF, 1 mM Na3VO4, 10 mM sodium pyrophosphate, 1 mM phenylmethylsulfonylfluoride, 10 mg/mL aprotinin, 10 mg/mL leupeptin, and 0.1 mg/mL soybean inhibitor). Cell lysates were centrifuged at 15,000 rpm for 5 min at 48°C. Protein concentration was measured by the Bradford method using bovine serum albumin (BSA) as a standard. Proteins (40 mg) were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred onto the nitrocellulose transfer paper. The membranes were blocked with 5% fat-free dry milk or 5% BSA for 1 h in Tris-buffered saline (TBS; 25 mM Tris–HCl, pH 7.6, and 150 mM NaCl) containing 0.1% Tween 20 (TBS-T), and then incubated with the following primary polyclonal antibodies: Oct-4 (Chemicon International), SOX-2 (Chemicon International), Nestin (Chemicon International), Smad5, Smad4, and STAT3. These were diluted 1:1000–1:5000 in 1% skim milk or BSA in TBS-T and incubated at 48°C overnight. After washing, the membranes were incubated with secondary peroxidase-conjugated anti-mouse and anti-rabbit antibodies diluted 1:1000 in 1% fat-free dry milk in TBS-T at room temperature for 1 h. Detection was achieved using an enhanced chemiluminescence kit (ECL Plus Western blotting kit; GE Healthcare Bio-Sciences, Piscataway, NJ). Quantitation of western blots by densitometry was performed with Image Master VSD software (Kodak Company, Tokyo, Japan).
Statistical analysis
All experiments consisted of at least three replicates. Statistical analysis was carried out using SAS software (Statistical Analysis System, Seoul, Korea). The mean values of the developmental data and nuclei counts were compared by the Student's t-test or analysis of variance, as appropriate. Data are presented as mean ± standard error of mean. Differences at p < 0.05 were considered statistically significant.
Results
Isolation of cells from adult human scar tissue
Several small, round outgrowing cells became visible around the explants after 24 h of incubation. These cells increased, turned into fibroblast-like cells, and developed on the bottom within 60–72 h. Cells (Fig. 1A, left) were harvested after 7–10 days of primary culture, and a sufficient number of cells were obtained after one to two subcultures. A total of four types of cells were established.
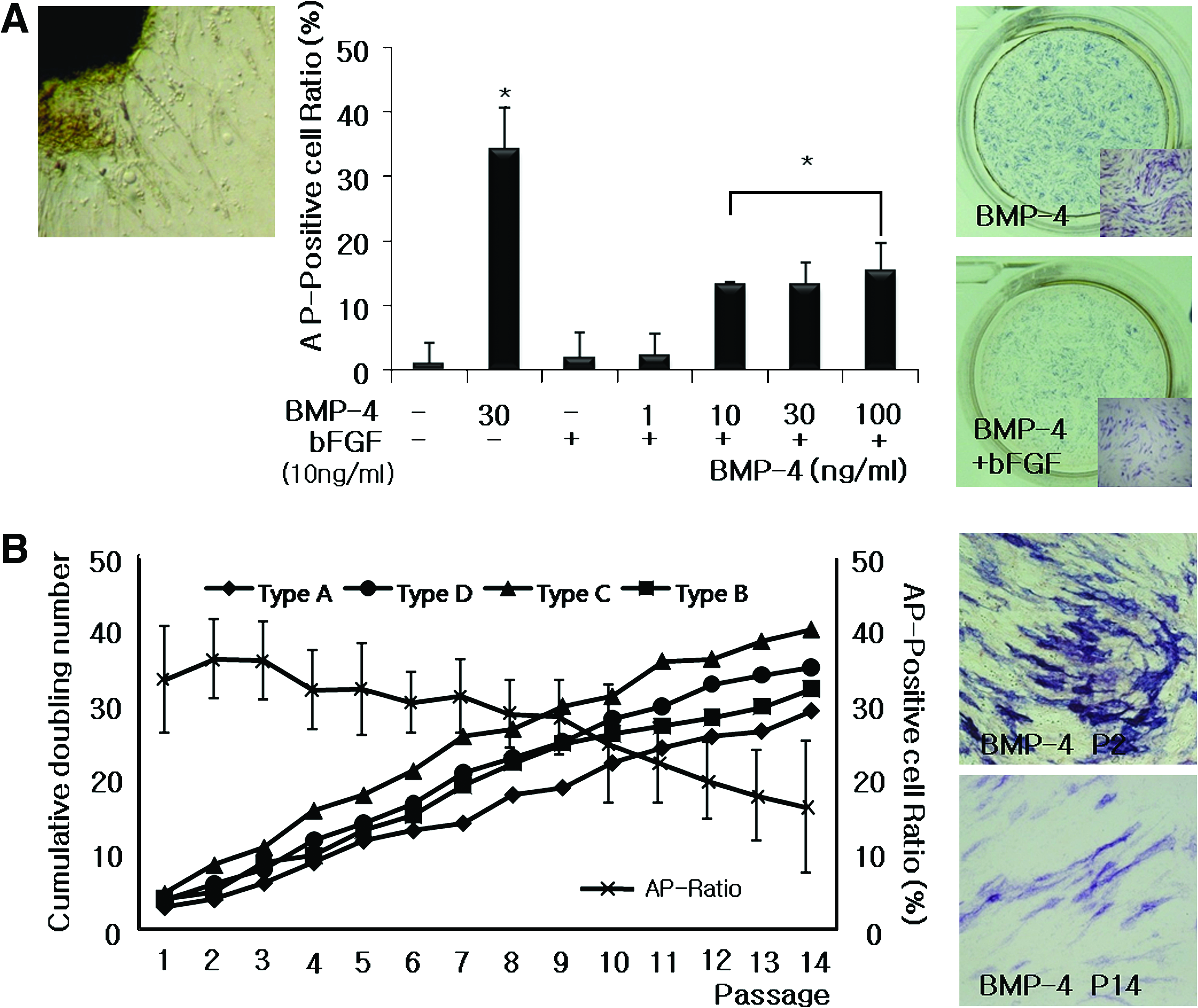
Stem-cell properties of hSTSCs. (
Measurement of AP activity with different concentrations of BMP-4 treatments of isolated cells
To determine the optimal dose of BMP-4, the cells were cultured in a BMP-4 dose-dependent manner, and AP staining was employed to evaluate the effect of BMP-4. AP-positive cells increased with BMP-4 dose increment (Fig. 1A, middle). AP-positive cells further increased when the cells were cultured in high density (data not shown). We decided that a BMP-4 concentration of 30 ng/mL was the optimal dose for subsequent experiments because there was no significant difference between the treatment concentrations of 30 and 100 ng/mL of BMP-4 (data not shown). However, it was found that AP-positive cells significantly decreased when the basic fibroblast growth factor (bFGF) (FGF2), which is generally known to facilitate self-renewal in human ESCs (Fig. 1A, middle and right), was added. The proportion of AP-positive cells was compared at the 30 ng/mL concentration of BMP-4 with and without bFGF treatment. The results were unquestionably different: 34.125 ± 6.325% of AP-positive cells were detected in BMP-4 alone, whereas 13.258 ± 3.251% of AP-positive cells were counted in the treatment of BMP-4 + bFGF. Although AP-positive cells increased along with the BMP-4 dose increment, they did not increase more than 20 ± 5.851% in the presence of bFGF (Fig. 1A, middle and right). Although samples from four different donors showed similar cell growth characteristics, overall AP intensity gradually declined to passage increment (Fig. 1B). Among them, type C (41.892 ± 4.587%) showed an excellent proliferation tendency and stable AP-positive cell ratio. For this reason, type C hSTSCs were used for subsequent experiments.
Marker expressions in the isolated cells
To identify the characteristics of the AP-positive cells, reverse transcriptase (RT)-PCR was performed using several stem-cell markers: Oct-4, c-kit, Sox2, Nestin, and AP in the cells with or without BMP-4 treatment. The results showed that all tested markers were expressed clearly in the BMP-4–treated group (Fig. 2A). However, the immunostaining results showed that only Oct-4 and CD90 were weakly expressed and that no other stem-cell markers were expressed (Fig. 2B).
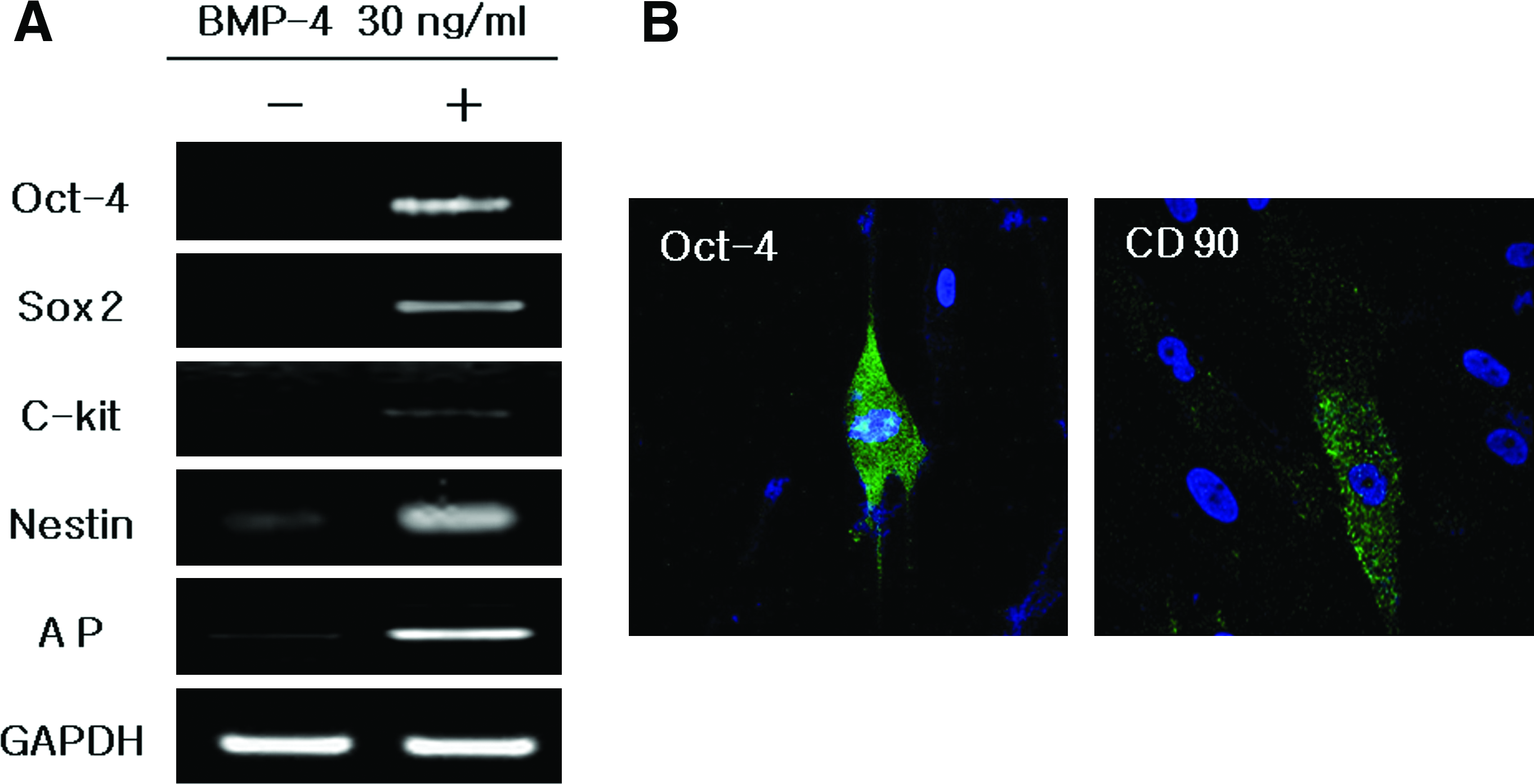
Gene expression patterns of hSTSCs with or without BMP-4 treatment in culture. Immunocytochemistry analysis of hSTSC with Oct-4 and CD90 antibodies (
Generation of OB
To isolate multipotent precursor cells, we modified a conventional technique devised to generate neural stem cells from the brain and skin.10,41,42 A number of free-floating, sphere-like OBs were observed after 24 h culture in SPM (DMEM/F12 + 20% KSR) containing 30 ng/mL of BMP-4. Various sizes (<10–150 μm) of free-floating, sphere-like OBs were distinctly generated and observed at this point during the culture (Fig. 3A, supplemental video; supplemental video available online at

Sphere-like OBs formation and classification and characterization of hSTSCs. (
Characteristics of hSTSCs in OBs
Immunocytochemistry was at first employed to determine the characteristics of free-floating OBs. OBs cultured for 24–36 h were collected and examined. In OBs of 50–100 μm, Oct-4 (ESC marker), SSEA-4, Nestin, TuJ1, and CD 90 (mesenchymal marker) were expressed (Fig. 4). However, OBs of over 100 μm in size did not express any markers at all. Unlike the normal cell culture result (Fig. 2B), a greater variety of stem-cell markers were expressed in the OBs with significantly higher expression rates (Fig. 4).
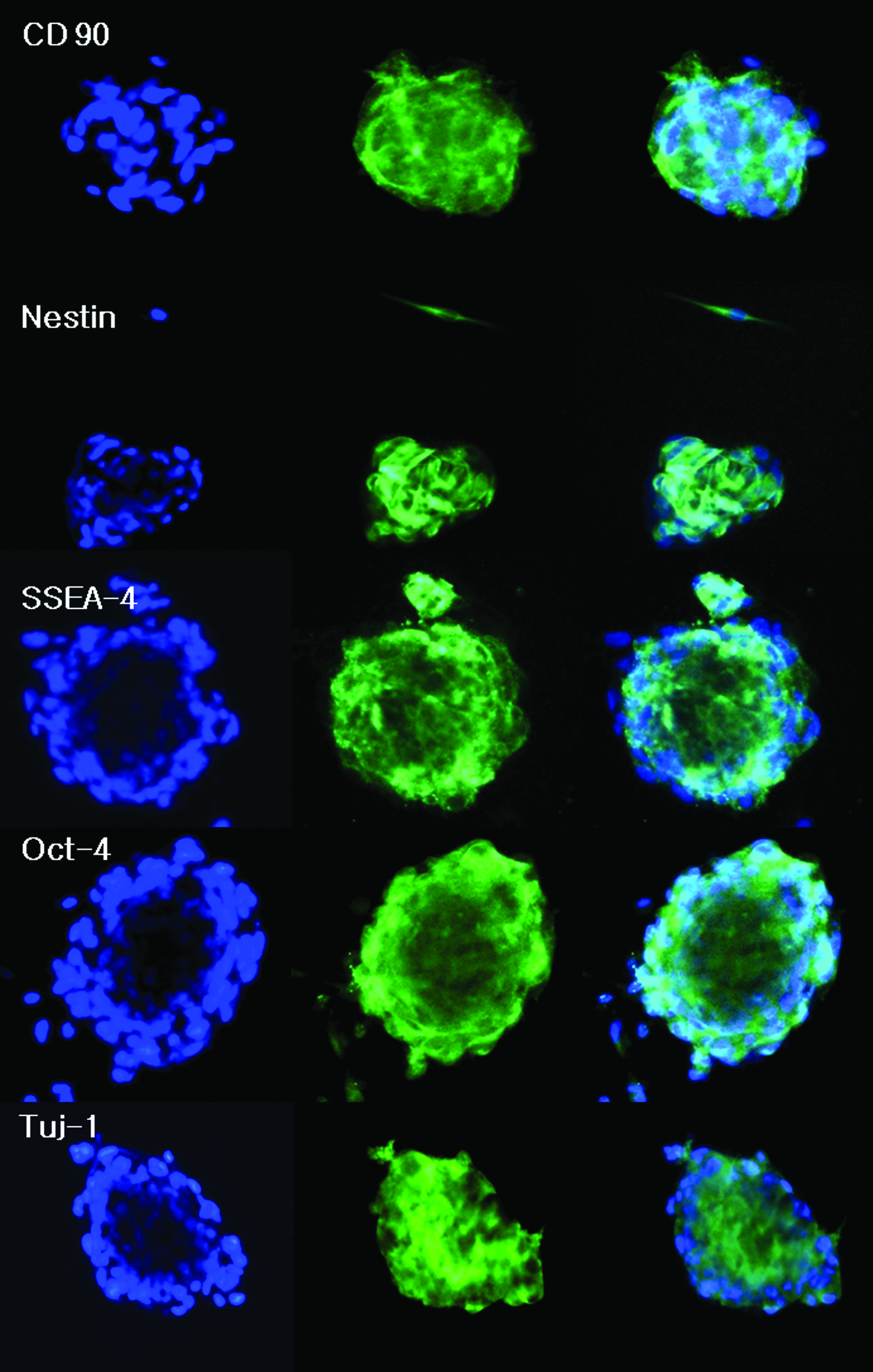
Immunofluorescence evaluation of stem-cell markers and neuronal lineage markers in hSTSCs (blue: 4′,6-diamidino-2-phenylindole, green: CD90, Nestin, SSEA-4, Oct-4, and TUJ1). SSEA-4, stage-specific embryonic antigen-4. Color images available online at
To confirm the immunocytochemistry results, mRNA expression and western blot analysis were also performed. Genes associated with pluripotent and multipotent stem cells were examined in both hSTSCs and hSTSCs in OBs. Transcription factors, such as Oct-4, c-kit, AP, Nestin, Sox 2, and Rex-1, were expressed in both cells and OB as indicated by RT-PCR (Fig. 5A, Table 1). Western blot analysis results showed that OB generated more proteins related to multipotent stem cells than normal cell cultures (Fig. 5B). All these results imply that continuous treatment with BMP-4 may result in transcriptional changes of human epidermal cells.
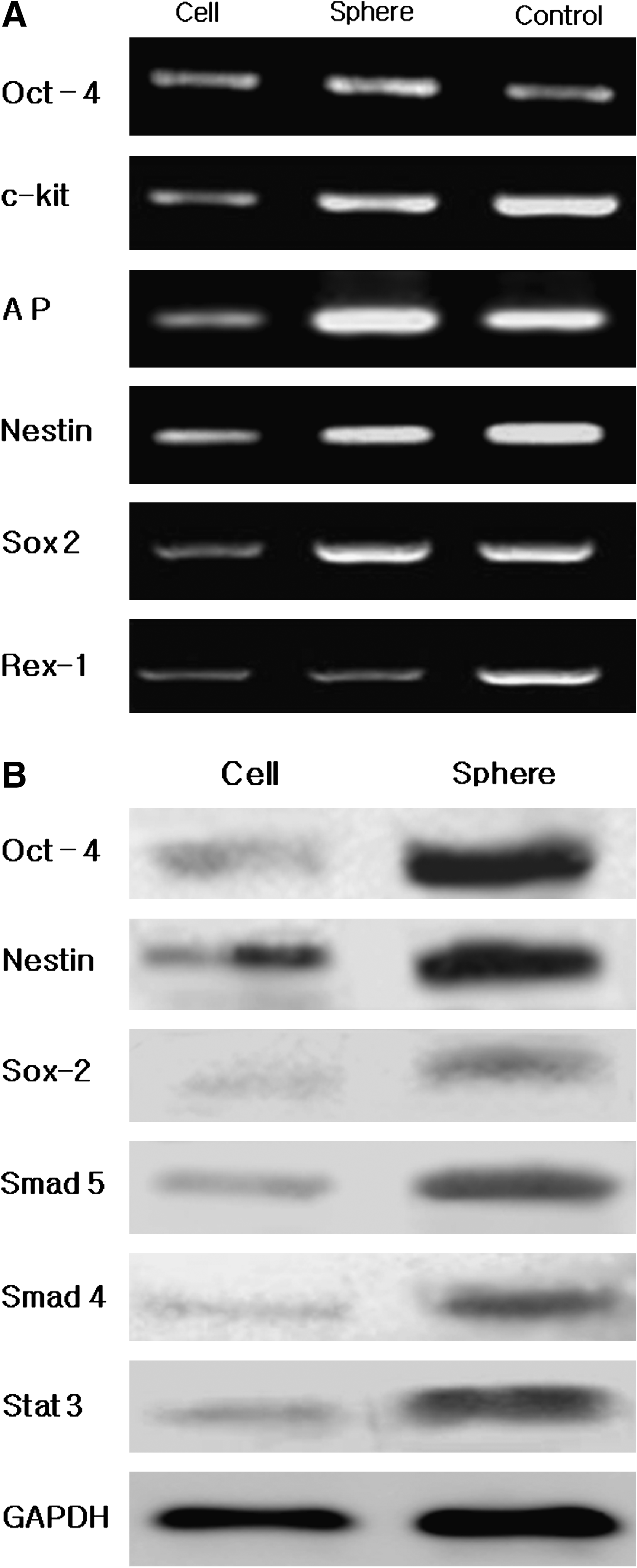
Molecular analysis of expression of stem-cell markers in hSTSCs. RT-PCR (
SMA, smooth muscle actin; NF, neuron filament; GFAP, glial fibrillary acidic protein; PGP, protein gene product; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Additionally, expressions of three germ layer–specific markers were also examined on the basis of the pluripotent gene expression results in cells and OBs (Fig. 6, Table 1). Interestingly, the obtained results showed that all tested markers were expressed in both normal cells and OBs and that the expression degree was generally higher in the OBs, except for a few genes. The ectodermal markers, protein gene product (PGP) 9.5 (a marker for mature neuronal cells) and GFAP, were highly expressed in OBs. Considerable amounts of integrin a6 mRNA was expressed in both hSTSCs and OBs. With respect to the mesodermal markers, smooth muscle actin and troponin T2 were expressed in both hSTSCs and OBs. Endodermal markers were also detected; amylase and albumin expression was stimulated by our new culture system.
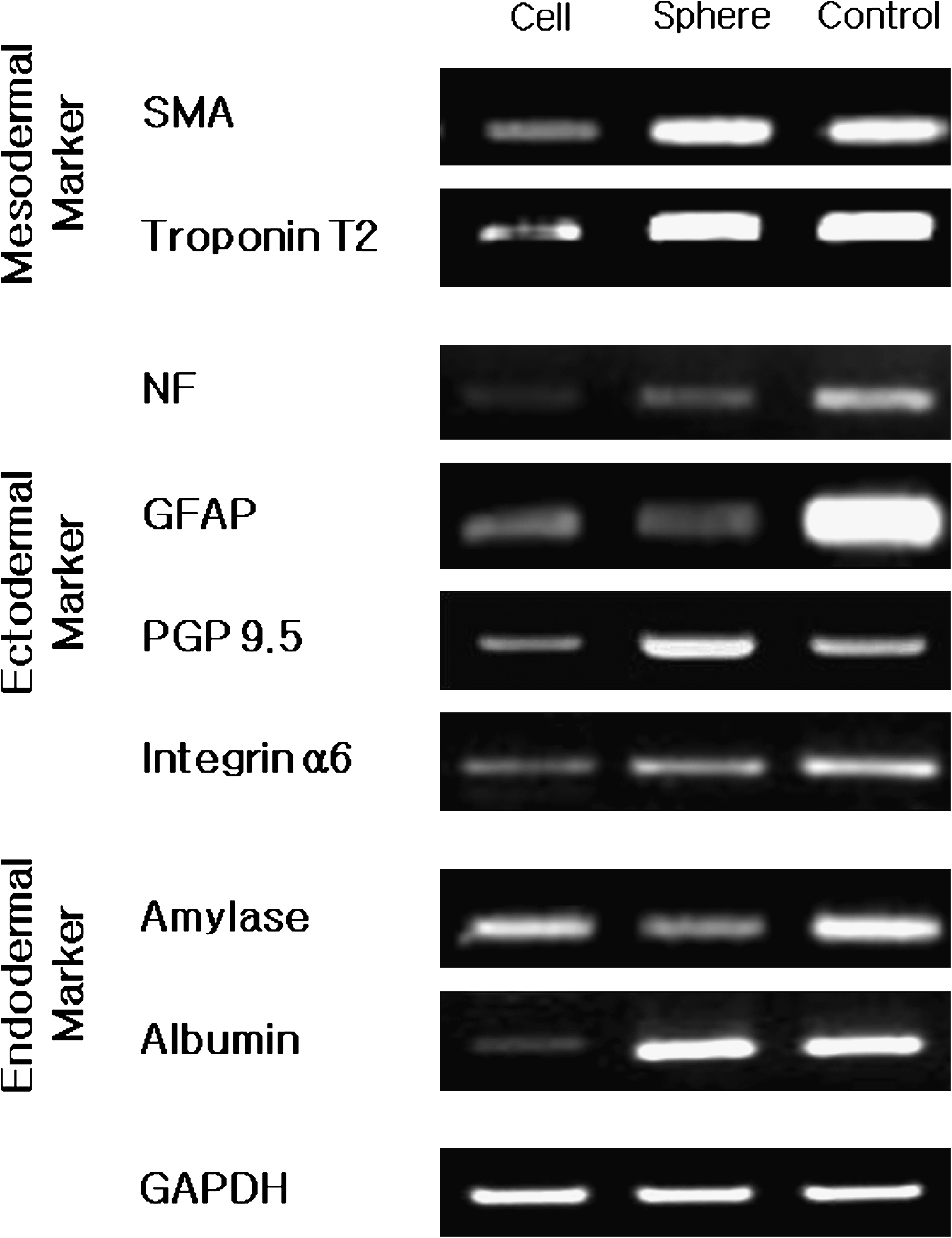
RT-PCR analysis of expression of various markers in hSTSCs. Cell: hSTSC cultured in the presence of BMP-4. Sphere: hSTSCs in OBs cultured in the sphere-producing medium. Control: human universal total RNA; GAPDH was used as a positive control.
Neuronal differentiation potential of hSTSCs
During the sphere-like OB-producing process, most hSTSC populations (more than 95%) possessed Nestin-positive characteristics, and these cell populations proliferated geometrically within 72 h. After that time, the number of Nestin-positive cells gradually reduced (Fig. 7A, Table 1). These OBs were used in neuronal differentiation. After 7 days of the differentiation process, typical neuronal filamentous structures were detected, and expression of neuronal cell–specific markers (GFAP, microtubule-associated protein 2, TuJ1, neural cell adhesion molecules, and neuron filament-M) was identified by immunocytochemistry and RT-PCR analysis (Fig. 7B, C, Table 1).

Neuronal differentiation potential of hSTSCs. (
Discussion
In this study, we established a novel innovative culture system to isolate and proliferate adult stem cells isolated from adult human tissue using SPM with BMP-4. Most previous studies had derived cells from the target tissue using conventional enzymatic dissociation. However, we employed explant culture of small pieces of the tissue to rule out any possibility of putative stem cell loss and damage by enzymes. The results verify that small quantities of hSTSCs were successfully cultured (Fig. 1).
We used SPM and BMP-4 for generating the sphere-like OBs. Conventionally, cells were cultured in DMEM/F12 containing B-27, epidermal growth factor, and bFGF for 3–7 days to generate neurosphere or sphere-like OBs.10,41,42 In this study, however, KSR in SPM and BMP-4 were used instead of B-27, epidermal growth factor, and bFGF. It is known that KSR does not contain the serum element. Therefore, we expected that (1) KSR might be appropriate for anchorage-independent culture, (2) KSR might prevent BMP-like factor that might be present in serum, and (3) the exact function of BMP-4 could be tested. Unlike previous reports, the results showed that our sphere culture system produces, within a very short period of time, OBs that have the same characteristics and potential as normal hSTSCs treated with BMP-4. Moreover, this system enables the proliferation of hSTSCs while maintaining the stem-cell potency (Figs. 3, 4, and 5A). This result suggests that KSR presumably plays a role in preserving stem-cell potency during culture along with BMP-4. KSR, especially the albumin-associated lipids in KSR, is already known to maintain and stimulate human ESC self-renewal.36,37
It is also worthy of mention that we used BMP-4 as a key factor for improving cell growth and endowing cells with multipotent stem-cell characteristics. BMP-4 has been known to suppress mouse ESC differentiation by induction of Id genes through the Smad pathway, 15 and mouse ESC-conditioned medium or BMP-4–pretreated skin keratinocytes alter not only the lineage-specific potential but also activate embryonic gene expression through MAPK and Smad pathways.38,39 This implies that continuous epidermal cell treatment with BMP-4 would result in transcriptional changes, such as the inhibition or downregulation of the lineage-committed differentiation of epidermal cells, and stimulate the transcription of multi- and even pluripotent genes (Figs. 2, 4, and 5A). 40 Western blot results supported that the Smad pathway was probably activated and, to some extent, participated in the transcriptional changes of the cells due to the BMP-4 treatment. Additionally, this signal pathway was supposed to prevent committed differentiation of the cells, to rapidly generate AP-positive cells, and to allow multi/pluripotent gene expression of the cells and OBs (Fig. 5B).
Taken together with all previous results, the three-germ layer marker analysis results also demonstrated that our hSTSCs and OBs possessed multilineage plasticity. As seen in Figure 6, expression of PGP 9.5 and glial fibrillary acidic protein (GFAP) reflects the possibility of differentiation to neuronal lineages, and the expression of integrin a6 signifies progenitor characteristic of the epithelial cell populations.21,22 It is known that smooth muscle actin and troponin T2, which serve as differentiation markers for smooth muscle cells and for heart and skeletal muscle, respectively, are expressed in hSTSCs and OBs.23,24 Most intriguingly, amylase and albumin, endodermal markers, were also detected. Taken together, these results strongly suggest that human adult stem/precursor cells and OBs with multilineage plasticity can be easily isolated from adult human cutaneous tissue using our culture system.11,25,26
Moreover, in vitro differentiation of these cells into neuronal progenitor cell lineages was successfully performed. Recent studies have reported that several kinds of skin-derived stem cells isolated and differentiated into specific cell types,11,27–30 but the isolated cell volume was very small and the cells took a long time to proliferate and differentiate26,31,32 with imperfect differentiation potential. In contrast, differentiation of our hSTSCs–OBs generated a large number of neurogenic progenitor cells, that is, Nestin-positive cells, within a very short period of time (Fig. 7A). These Nestin-positive cells demonstrated the diverse physiological status of neuronal cells (Fig. 7B, C). This neuronal-specific gene expression (GFAP, microtubule-associated protein, and TuJ1) was not observed in somatic cells.
Recently, induced pluripotent stem cells (iPSCs) have been spotlighted as a new stem cell to replace ESCs. This iPSCs have the advantages of preserving the properties of ESCs without the ethical issues and overcoming the low efficiency of adult stem-cell proliferation. Generally, iPSCs were created by introducing four transcription factors (Oct-4, Sox2, Klf4, and c-Myc) to the fibroblast. 33 However, this procedure might cause infection via the vector system and tumorigenesis for foreign genes. Consequently, iPSCs are deemed unsuitable for clinical applications, but can be used for research.33,34 For this reason, we cautiously claim that the work detailed in our study is valuable for the proliferation and amplification of adult stem cells, using useless human scar tissue, through environmental regulation, without genetic modifications.
In conclusion, we established a totally new culture system to isolate and proliferate adult stem cells from adult human tissue using SPM with BMP-4. This system allows the formation of multi/pluripotent stem cells without genetic modifications of adult somatic cells. Further, the hSTSCs cultured in this system yielded promising results for neuronal differentiation. It is considered that the results of this research may contribute to the development of new cell therapy systems.
Author Contributions
Ji Hoon Yang: conception and design, collection and assembly of data, data analysis and interpretation, and manuscript writing; Sang Woo Shim: conception and design, data analysis and interpretation, and manuscript writing; Bo Yon Lee: administrative support; Hoon Taek Lee: conception and design, analysis and interpretation, and final approval of manuscript.
Footnotes
Acknowledgments
This work was supported by grant from Konkuk University in 2009 (Code: 2009-A019-0041).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.