
Abstract
Cartilage healing by tissue engineering is an alternative strategy to reconstitute functional tissue after trauma or age-related degeneration. However, chondrocytes, the major player in cartilage homeostasis, do not self-regenerate efficiently and lose their phenotype during osteoarthritis. This process is called dedifferentiation and also occurs during the first expansion step of autologous chondrocyte implantation (ACI). To ensure successful ACI therapy, chondrocytes must be differentiated and capable of synthesizing hyaline cartilage matrix molecules. We therefore developed a safe procedure for redifferentiating human chondrocytes by combining appropriate physicochemical factors: hypoxic conditions, collagen scaffolds, chondrogenic factors (bone morphogenetic protein-2 [BMP-2], and insulin-like growth factor I [IGF-I]) and RNA interference targeting the COL1A1 gene. Redifferentiation of dedifferentiated chondrocytes was evaluated using gene/protein analyses to identify the chondrocyte phenotypic profile. In our conditions, under BMP-2 treatment, redifferentiated and metabolically active chondrocytes synthesized a hyaline-like cartilage matrix characterized by type IIB collagen and aggrecan molecules without any sign of hypertrophy or osteogenesis. In contrast, IGF-I increased both specific and noncharacteristic markers (collagens I and X) of chondrocytes. The specific increase in COL2A1 gene expression observed in the BMP-2 treatment was shown to involve the specific enhancer region of COL2A1 that binds the trans-activators Sox9/L-Sox5/Sox6 and Sp1, which are associated with a decrease in the trans-inhibitors of COL2A1, c-Krox, and p65 subunit of NF-kappaB. Our procedure in which BMP-2 treatment under hypoxia is associated with a COL1A1 siRNA, significantly increased the differentiation index of chondrocytes, and should offer the opportunity to develop new ACI-based therapies in humans.
Introduction
Among the various scaffolds, the chondrogenic properties of type I/III collagen sponges have already been demonstrated.4,5 This collagen sponge scaffold was prepared with type I atelocollagen derived from calf skin and was chemically crosslinked to obtain a three-dimensional (3D) scaffold with good mechanical and thermal stability. 6 Furthermore, chondrocytes cultured in these collagen sponge scaffolds stably express metalloproteinases MMP-1 and MMP-13 in comparison to cells embedded in collagen gels, suggesting that this biomaterial is in favor of a stabilized cartilage matrix. 4 AC is avascular with an oxygen concentration varying from 1% to 7% and hypoxia had been demonstrated to be essential for the differentiation of chondrocytes7,8 or mesenchymal stem cells (MSCs). 9 In this sense, the transcriptional complex hypoxia-inducible factor (HIF)-1 when binding to specific hypoxia responsive elements, initiates the transcription of aggrecan and Sox9, 8 a transcription factor essential for chondrocyte differentiation and cartilage formation. 10
Several growth and differentiation factors involved in cartilage development and homeostasis have been identified and used for cartilage repair. Among them, the insulin-like growth factor I (IGF-I) and bone morphogenetic protein-2 (BMP-2) are of particular interest for cartilage engineering. IGF-I, one of the most important anabolic growth factors for AC, plays a key role in cartilage homeostasis, balancing proteoglycans synthesis and breakdown. 11 IGF-I is not only capable of extending the chondrogenic potential of dedifferentiated chondrocytes in vitro, 12 but it also enhances the chondrogenesis of bovine articular chondrocytes seeded into polymer scaffolds. 13 However, others factors, such as BMP-2, seem to have a better redifferentiation potential. 14 BMPs are mainly related to bone and cartilage formation. 15 In particular, BMP-2 is involved in chondrocyte differentiation by enhancing type II collagen and aggrecan expression, and in matrix maturation, preventing dedifferentiation of chondrocytes cultured in monolayer.16–19 Type II collagen is an homotrimer composed of α1(II) chains encoded by the COL2A1 gene. Two forms of type II collagen can be generated by alternative COL2A1 mRNA splicing 20 : the long type IIA form is synthesized by chondroprogenitor cells and noncartilaginous epithelial and mesenchymal cells, whereas the short type IIB form is synthesized by mature chondrocytes.
At the transcriptional level, although IGF-I stimulatory effects on COL2A1 transcription have been already analyzed, 21 those of BMP-2 in chondrocytes have not. In rabbit articular chondrocytes (RACs), IGF-I stimulatory effects on COL2A1 transcription are mediated by an increase in the DNA-binding activity of Sox-9/Sox-6/L-Sox-5 and Sp1 in the intronic enhancer of the gene. These trans-activators are known to be implicated in the regulation of this gene.22–24 These studies were performed with chondrocytes cultured in monolayer, but no study has been performed with chondrocytes cultured in 3D scaffolds.
In this study, we aimed at improving a procedure to reinitiate the differentiation of dedifferentiated human articular chondrocytes (HACs) from OA cartilage, but also at deciphering the molecular mechanisms associated with a functional cartilage matrix synthesis. Chondrocytes were cultured, after expansion, in type I/III collagen sponge scaffolds, under low oxygen tension, with two chondrogenic factors: BMP-2 and/or IGF-I. In our 3D scaffold cultures, we demonstrated that (1) hypoxia can potentiate chondrocyte redifferentiation; (2) BMP-2 was a better candidate than IGF-I for chondrocyte redifferentiation, without any signs of hypertrophy; and (3) that siRNA targeting COL1A1 mRNA is essential for stabilizing the chondrocyte phenotype. Moreover, we identified the trans-activators and the trans-inhibitors involved in the regulation of COL2A1 gene expression by BMP-2 and IGF-I in our 3D scaffold. Overall, this study provides new insights for the development of innovative and clinically relevant methods for repairing AC in humans.
Materials and Methods
Chondrocyte cultures
HACs were prepared from macroscopically healthy zones of femoral heads obtained from patients undergoing joint arthroplasty (age range: 52–83 years) as previously described. 8 All patients signed an informed consent agreement form, which was approved by the local Ethics Committee. Chondrocytes were seeded at 4×104 cells/cm2 in plastic dishes, in an atmosphere of 5% CO2, with a medium consisting of the Dulbecco's modified Eagle's medium (DMEM; Gibco, Invitrogen) supplemented with 10% fetal calf serum (FCS; Invitrogen), 100 IU/mL of penicillin, 100 μg/mL of erythromycin, and 0.25 μg/mL of fungizone. After one passage, trypsinized chondrocytes were recovered and used for redifferentiation experiments in 3D collagen scaffolds manufactured by Symatèse Biomatériaux. These collagen sponge scaffolds were composed of native type I collagen (90%–95%) and type III collagen (5%–10%) from calf skin. Collagen sponges, 2 mm thick and 5 mm in diameter, were crosslinked using glutaraldehyde to increase their stability and sterilized with β radiations. 6 HACs were seeded onto the sponges (4×104 cells/sponge i.e., 10×106 cells/cm3) in 96-well culture plates and incubated at 37°C and 5% CO2 for 16 h with the DMEM+2% FCS. The following day, cells were incubated in the DMEM+2% FCS, with or without 50 ng/mL of BMP-2 (R&D Systems), 10 ng/mL of IGF-I (Sigma-Aldrich), or both. This point established day 0 and HACs were incubated in normoxia (21% O2) or in hypoxia (3% O2) for 3, 7, and 14 days. Hypoxic cultures were performed in a sealed chamber as previously described. 25 The medium was changed and collected on days 0, 3, 5, 7, 10, 12, and 14, and sponges were harvested on days 0, 3, 7, and 14. Each set of samples had the same medium replacement protocol.
RACs were isolated from AC slices of shoulders and knees of 3-week-old male rabbits. The cells were seeded at 3.5×105 cells per 9.6 cm2 for transient transfection experiments (P0) as previously described. 21 They were also subcultured as monolayers until passage 3 and seeded at 3.5×105 cells per 9.6 cm2 for transient transfection experiments on dedifferentiated chondrocytes (P3).
Electron microscopy
Chondrocytes cultured in collagen sponges were fixed for 24 h with 2.5% glutaraldehyde in phosphate-buffered saline (PBS) at 4°C, and washed in a 0.1 M phosphate buffer, pH 7.4. Samples were postfixed with 1% osmic acid for 2 h, washed, and dehydrated in a graded series of ethanol. Then, they were critical point-dried, coated with a platinum layer for scanning electron microscopy (SEM) observations (JEOL 6400F). For transmission electron microscopy (TEM), after dehydration, samples were embedded in an epoxy resin (Epon 812; EMS). Ultrathin sections were prepared with an ultramicrotome (Ultracut R LEICA), mounted on grids and contrasted with uranyl acetate/lead citrate and visualized by TEM (JEOL 1011).
Reverse transcription-polymerase chain reaction analysis
Total RNA was extracted and reversed transcribed into cDNA before real-time polymerase chain reaction (PCR) amplification as previously described.
26
The ribosomal protein L13a (RPL13a) was used as an endogenous reference gene. The sequences of the primers are given in references26,27 or were newly designed from sequences publically available in databanks using Primer Express Software (Supplementary Table S1; Supplementary Data are available online at
Western blotting
Following treatment, cells cultured in collagen sponges were rinsed once with ice-cold PBS, crushed, and lysed in the RIPA buffer to prepare cellular extracts. 29 Cellular extracts underwent sodium dodecyl sulfate–polyacrylamide gel electrophoresis and were electrotransferred to a polyvinylidene difluoride transfer membrane (Millipore). Membranes were probed with type X collagen (Sigma Aldrich), Sox9 (Santa Cruz Biotechnology, Inc.), and GAPDH (Santa Cruz Biotechnology, Inc.) antibodies, and then washed and incubated with anti-goat secondary peroxidase-conjugated antibodies (Santa Cruz Biotechnology, Inc.). The signals were revealed with ECL plus western blotting detection reagent+(Santa Cruz Biotechnology, Inc.).
Protein quantification
Enzyme-linked immunosorbant assays (ELISA; TECOmedical) were performed on the culture media of HAC cultures. The amount of procollagen II C-propeptide and aggrecan chondroitin sulfate 846 epitopes were measured in a competitive enzyme immunoassay, whereas the C-terminal propeptide of type I collagen was measured in a sandwich enzyme immunoassay. The experiments were carried out according to the manufacturer's recommendations. In the case of redifferentiation experiments in collagen sponges, for each set of cultures, the concentrations were calculated in ng/mL, on days 0, 3, 5, 7, 10, 12, 14, and converted into ng. Values were expressed as percentages, relative to day 0 and cumulated. Therefore, results were expressed as percentages of induction, relative to day 0. For the gene silencing experiments, the concentrations of type I or II collagens were calculated in ng/mL and normalized to the total protein content of each time point. Then, results were expressed as mg of collagen per gram of the total protein content.
Immunochemistry
Following treatment, cells cultured in collagen sponges were rinsed once with a 0.1 M phosphate buffer, pH 7.4, fixed with buffered 4% paraformaldehyde for 16 h at 4°C, and rinsed with the 0.1 M phosphate buffer. They were then embedded in 5% agar (Super LM) in the 0.1 M phosphate buffer. Then, 50-μm floating sections were cut with a vibratome (MICROM HM 650V) and mounted on microscope slides. They were incubated for 5 min in PBS/0.5% triton (Sigma Aldrich), rinsed in PBS, incubated for 30 min at 37°C in PBS/0.2% hyaluronidase (Sigma Aldrich), rinsed in PBS, and treated with PBS/10% bovine serum albumin (Sigma Aldrich). Immunohistochemical staining was carried out using polyclonal-specific antibodies against type I and II collagens diluted to 1:100 (Novotec). These primary antibodies were revealed using a goat anti-rabbit immunoglobulin G conjugated to Alexa Fluor 546 diluted to 1:2000 from a 2 mg/mL solution (Invitrogen). As controls, primary antibodies were omitted. Slides were then treated with the UltraCruzTM Mounting Medium with 4′,6-Diamidino-2-phenyindole,dilactate (Santa Cruz Biotechnology, Inc.). The observations were made on a confocal laser scanning microscope (Olympus FV1000). The same parameters were used for all acquisitions and representative images are shown in the figures.
Nuclear extracts and DNA-binding analysis
Alternatively, after washing with PBS, HACs cultured in sponges were also lysed in hypotonic and hypertonic buffers to prepare nuclear extracts as previously described. 29 Electrophoretic mobility shift assays (EMSAs) were performed with the oligonucleotides presented in Supplementary Table S1 and with their specific binding buffer.21,27,30
Transient transfection experiments
Eighty percent confluent RACs in six-well plates (P0 or P3) were transiently transfected using the calcium phosphate precipitation method as described previously. 22 Confluent HACs in P0 were collected and were transiently transfected as previously described. 31 Luciferase reporter plasmids containing various deletions of COL2A1 promoter/first intron were cotransfected with a pSV-40 β-galactosidase expression vector as an internal control of transfection efficiency. The reporter constructs pGL2—3.374 kb, pGL2—3.316 kb, pGL2—3.059 kb, pGL2—2.367 kb, pGL2—1.167 kb, pGL2—0.387 kb, and pGL2—0.110 kb cover, respectively, the −932/+2842 bp, −932/+2384 bp, −932/+2127 bp, −1744/+623 bp, −544/+623 bp, −266/+121 bp, and −63/+47 bp of the promoter and the first intron of the COL2A1 gene.32,33 After 16 or 5 h of transfection for RACs or for HACs, respectively, the medium was changed to the DMEM+2% FCS and chondrocytes were treated with or without 50 ng/mL of BMP-2. After 24 h, samples were harvested, the protein content and the luciferase and β-galactosidase activities were assayed. Luciferase activities were normalized for transfection efficiency and protein content. Transcription activities were calculated as relative luciferase units (RLUs). Due to the differences in the extent of transcriptional activities for each set of cultures, RLU was adjusted to the control sample that did not receive BMP-2. Percentages of RLU relative to the control were expressed as means±standard deviation from a minimum of three independent experiments performed in triplicate.
Gene-silencing experiments
For the gene-silencing experiments, 2×106 HACs (P1) were nucleofected, using the Amaxa apparatus, with 1 μg siRNA against COL1A1 (target sequence: 5′-ACCAATCACC TGCGTACAGAA-3′; Qiagen) or with a negative control siRNA (target sequence: 5′-AATTCTCCGAACGTGTCA CGT-3′; Qiagen). HACs were incubated in a medium (DMEM+20% FCS) for 1 h at 37°C, centrifuged, and seeded onto type I collagen sponges as described above with slight modifications: the HACs were included in sponges and after a 1-h incubation at 37°C and 5% CO2, the culture medium (DMEM+2% FCS) was added to each well, with or without BMP-2 (50 ng/mL). Then, sponges were transferred to hypoxia for 24 and 48 h. The culture media were collected for ELISA on type I and II collagens, whereas the cells cultured in collagen sponges were processed for RT-PCR analysis.
Statistical analysis
For RT-PCR analysis, data are presented as box plots, representative of four experiments performed in triplicate. For protein quantification, box plots were used to present data from three or four experiments with the minimum value the 25th, 50th (median), 75th percentiles, and the maximum value. Means were shown as lozenges. The Mann–Whitney U test was used to determine if there were significant differences between the control and treated groups. p Values less than 0.05 were considered significant: ***p<0.001, **p<0.01, and *p<0.05.
Results
Chondrocytes cultured in collagen sponges show sustained metabolic activity associated with focal adhesion and primary cilia interactions
SEM images of the collagen I/III sponges used in this study showed, as expected, a porous matrix with a large pore size (about 100 μm of diameter) (Fig. 1A), leaving large spaces between the collagen walls for cells to grow. When scaffolds were seeded with 10×106 cells/cm3, the distribution throughout the sponge was homogeneous with rounded cells in the superficial (Fig. 1B) and internal areas (Fig. 1C) of the scaffold. The cells had microvilli and vesicle-like protrusions on their surfaces. They wrapped around the collagen network. To complete the outer cell morphology of chondrocytes, we next examined subcellular features of HACs seeded in sponge by TEM. TEM images showed that chondrocytes displayed a chondrocyte-like morphology rather than a fibroblast-like morphology with an ovoid cell shape and numerous microvilli on their surface (Fig. 2A). Cells also showed an ovoid nucleus, an extensive rough endoplasmic reticulum, and many vesicles. These morphological features were previously described with chondrocytes grown on the type I/III collagen scaffold.34,35 They had close contacts with the collagen scaffold and released granular material, attesting to processes of extracellular matrix (ECM) synthesis. Otherwise, primary cilia, involved in the signaling processes related to the synthesis and maintenance of the ECM,36,37 were detected in contact either with neo-synthesized collagen lattices or with the collagen scaffold (Fig. 2B). The micrographs showed that the collagen scaffolds support cell adhesion and metabolic activity of chondrocytes. Furthermore, the ratio of COL2A1/COL1A1 mRNA levels, which monitors the differentiation status of chondrocytes increased first, as early as they were cultured 3 days in the 3D scaffolds, and then decreased. On the contrary, the same ratio decreased during all the monolayer culture. This scaffold reversed chondrocytes dedifferentiation, but was not sufficient, by itself, to stabilize the chondrocyte phenotype during 14 days of culture.
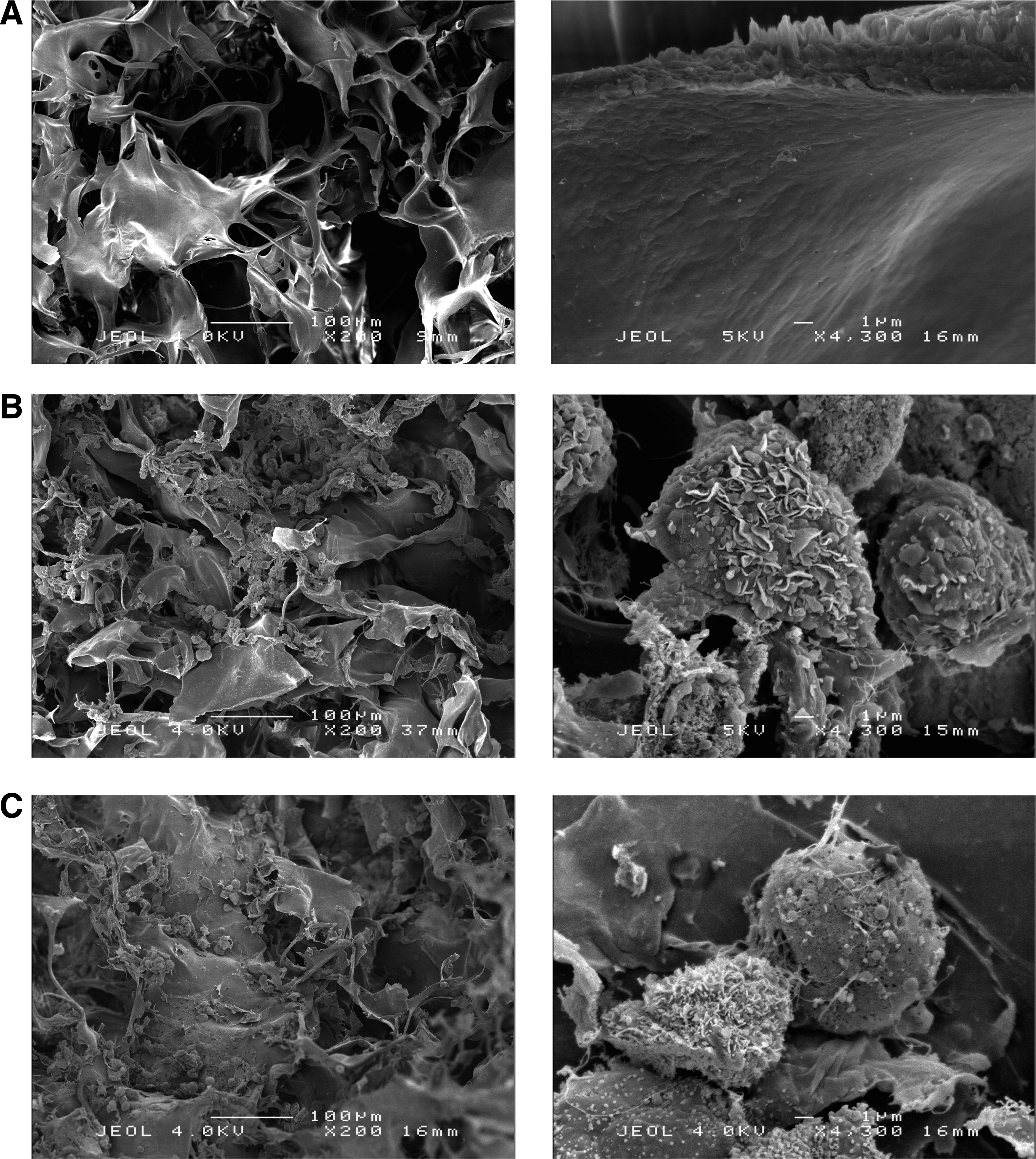
Scanning electron micrograph (SEM) of human articular chondrocytes (HACs) cultured in a type I collagen sponge. HACs were cultured in a type I collagen sponge for 3 days in normoxia and SEM observations were then realized. Left panels, original magnification ×200 and scale bar: 100 μm. Right panels, original magnification×4300 and scale bar: 1 μm.
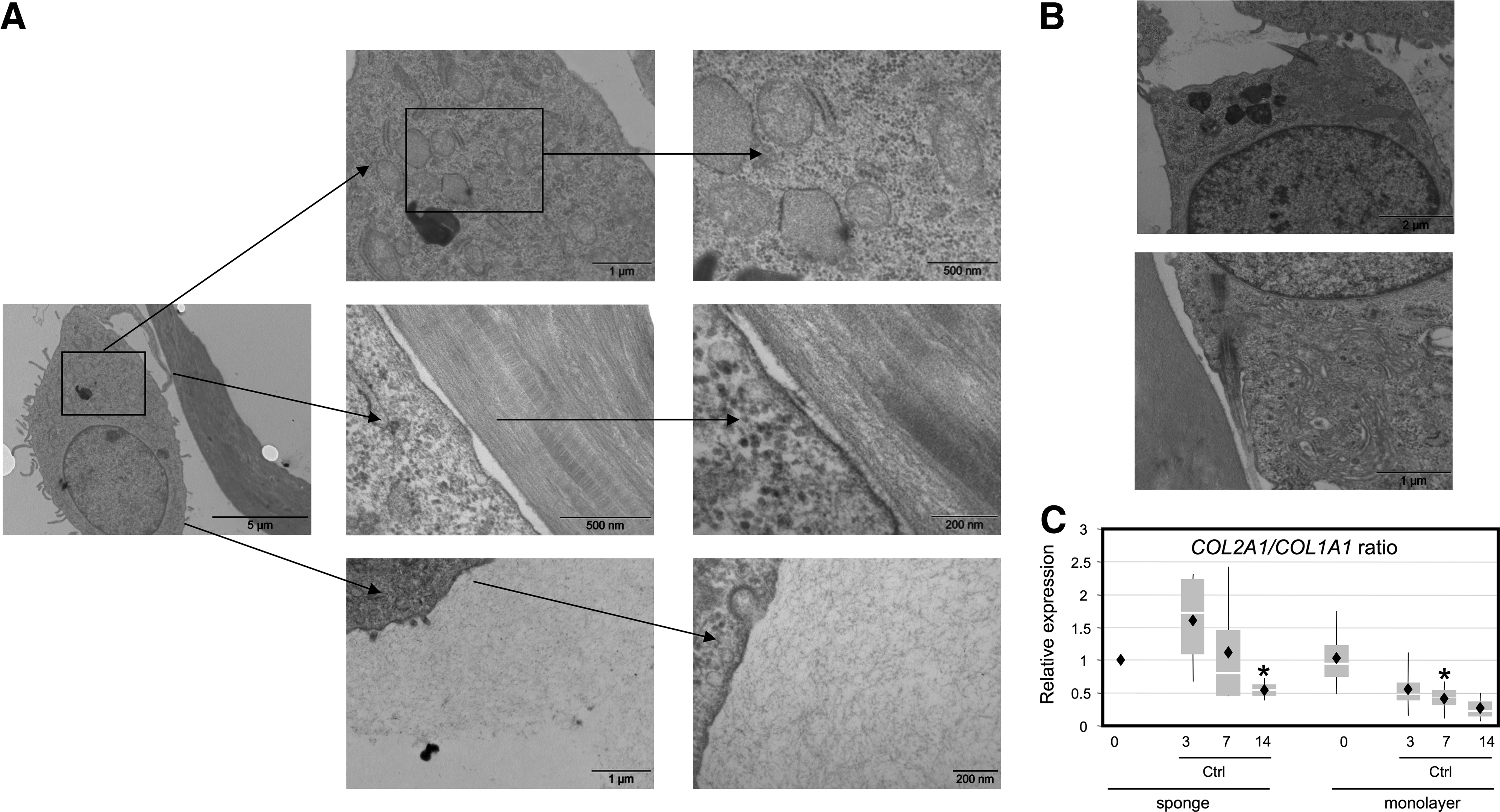
Metabolic activity of HACs cultured in a type I collagen sponge.
Phenotypic profiles of chondrocytes cultured in collagen scaffolds with chondrogenic factors
To determine the differentiation status of HACs, we analyzed the relative abundance of transcripts encoding eight proteins associated with differentiated, dedifferentiated and hypertrophic chondrocytes. In addition to the two well-known chondrocyte phenotype markers, type II collagen and aggrecan, and SOX9, which positively regulates their expression, the steady-state mRNA levels of two type II collagen isoforms, prechondrogenic type IIA collagen and mature type IIB collagen, were evaluated. Among molecules unsuitable for cartilage engineering (e.g., type X collagen, an indicator of chondrocyte hypertrophy and osteocalcin, a bone-specific marker), type I collagen was used to calculate a differentiation index corresponding to the ratio of type II collagen mRNA to type I collagen mRNA. Finally, the phenotypic profile was completed by the ratio of type IIB collagen mRNA to type IIA collagen mRNA, where type IIB collagen characterizes well-differentiated chondrocytes and type IIA is representative of cells more likely to be undergoing chondrogenesis. 38
The expression of type II collagen and its spliced variants gradually increased during 3D scaffold culture in hypoxia or remained stabilized in normoxia (Fig. 3). Moreover, the collagen sponge culture favored the ratio of COL2B/COL2A mRNA levels. The addition of BMP-2 or IGF-I improved the expression of the different type II collagen mRNAs (from 2- to 15-fold), with a greater increase in the BMP-2 treatment compared to IGF-I, whatever the oxygen tension conditions. Furthermore, in hypoxia, BMP-2 induced a relative transcript abundance 3–7 times higher than in normoxia for COL2A1, COL2A, and COL2B mRNA at day 7. Therefore, the BMP-2 chondrogenic effect is heightened in hypoxia, and the same conclusions also hold for IGF-I, although the expression of type II collagen mRNA at day 14 returned to day 3 levels. By day 7, in hypoxia, COL2A1 mRNA levels were higher with BMP-2-treated cells than with IGF-I treatment. Furthermore, BMP-2 was a better inducer of type IIB collagen mRNA than IGF-I, which favored type IIA mRNA expression. The combination of both BMP-2 and IGF-I had an additive effect only in hypoxia and on all forms of type II collagen, but the ratio of COL2B/COL2A mRNA was significantly higher for BMP-2 alone, indicating more differentiated chondrocytes in the BMP-2 treatment.
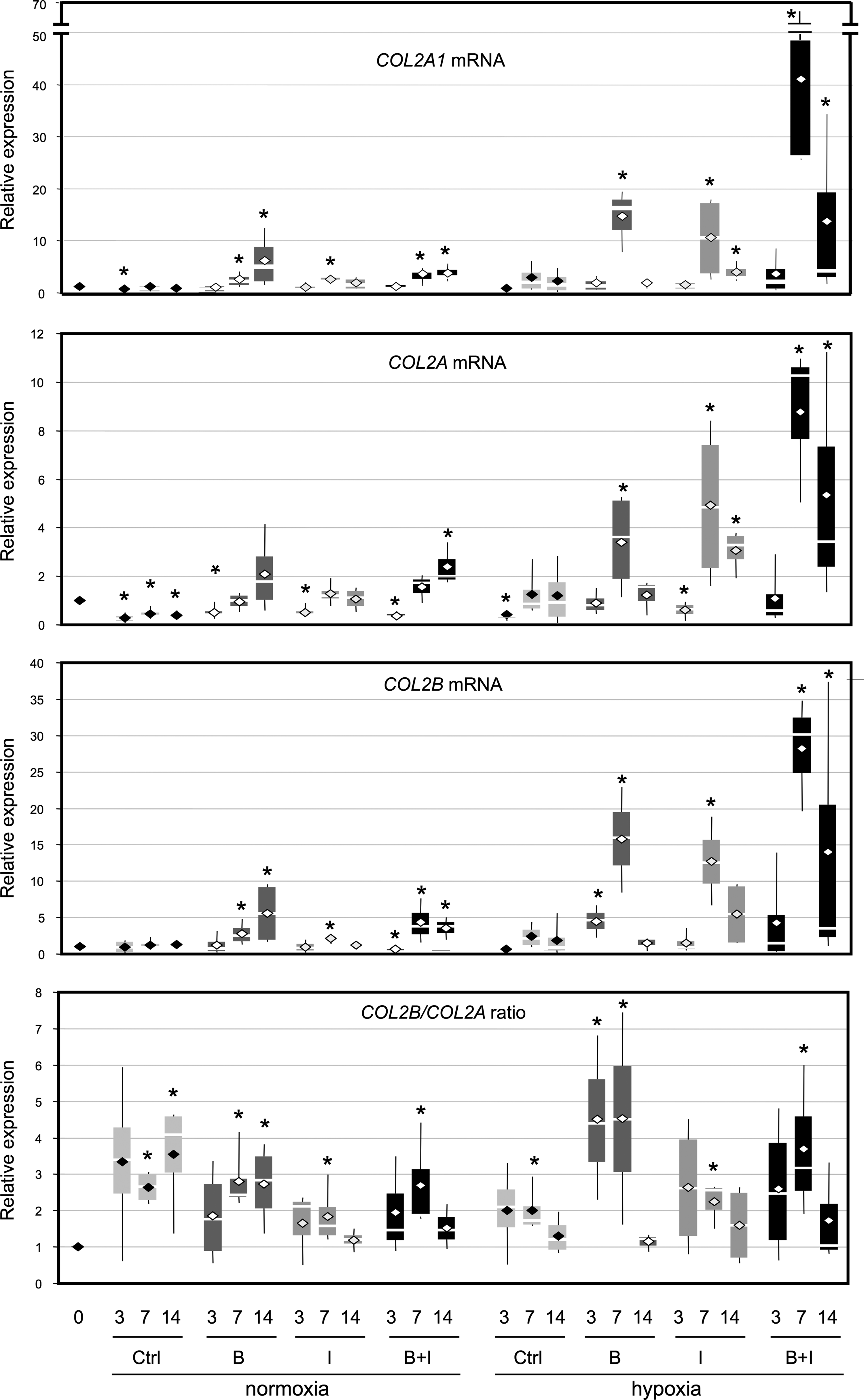
Effect of culture conditions on type II collagen mRNA expression. HACs were cultured in type I collagen sponges for 3, 7, and 14 days in normoxia or in hypoxia, in the absence (Ctrl), or in the presence of 50 ng/mL of bone morphogenetic protein-2 (BMP-2) (B), or 10 ng/mL of insulin-like growth factor I (IGF-I) (I), or both BMP-2 and IGF-I (B+I). The steady-state mRNA levels for type II collagen were measured using real-time RT-PCR with primers or probes specific to type IIA collagen (COL2A mRNA), type IIB collagen (COL2B mRNA), or the total amount of type IIA and IIB collagen (COL2A1 mRNA). All results were normalized to RPL13a mRNA, compared to untreated sponge-cultured cells at day 0, and presented as the relative expression of each gene. For each set of experiments, we also calculated the COL2B/COL2A ratio. Box plots represent four independent experiments in triplicate. Statistically significant differences between the untreated cells in sponge at day 0 and treated cells were determined using the Mann–Whitney U test (*p<0.05).
Regarding aggrecan, the collagen sponge culture without growth factors did not significantly increase ACAN expression, whereas the addition of BMP-2 and IGF-I had a beneficial effect mainly in hypoxia (Fig. 4). In normoxia, the best effect was observed with BMP-2 (four times greater at day 14), which nevertheless, remained lower than in hypoxia at day 7. In hypoxia, BMP-2 displayed also a stronger effect than IGF-I and only the cotreatment with IGF-I and BMP-2 at day 14 had an additive effect. In collagen sponge cultures in normoxia, SOX9 mRNA levels remained relatively stable whatever the growth factor treatment. In hypoxia and in 3D scaffold culture, chondrocytes increased their SOX9 mRNA levels, which were enhanced in the BMP-2 treatment (from 2- to 3-fold), but less so in the IGF-I treatment and only transiently in the combined treatment with both BMP-2 and IGF-I.
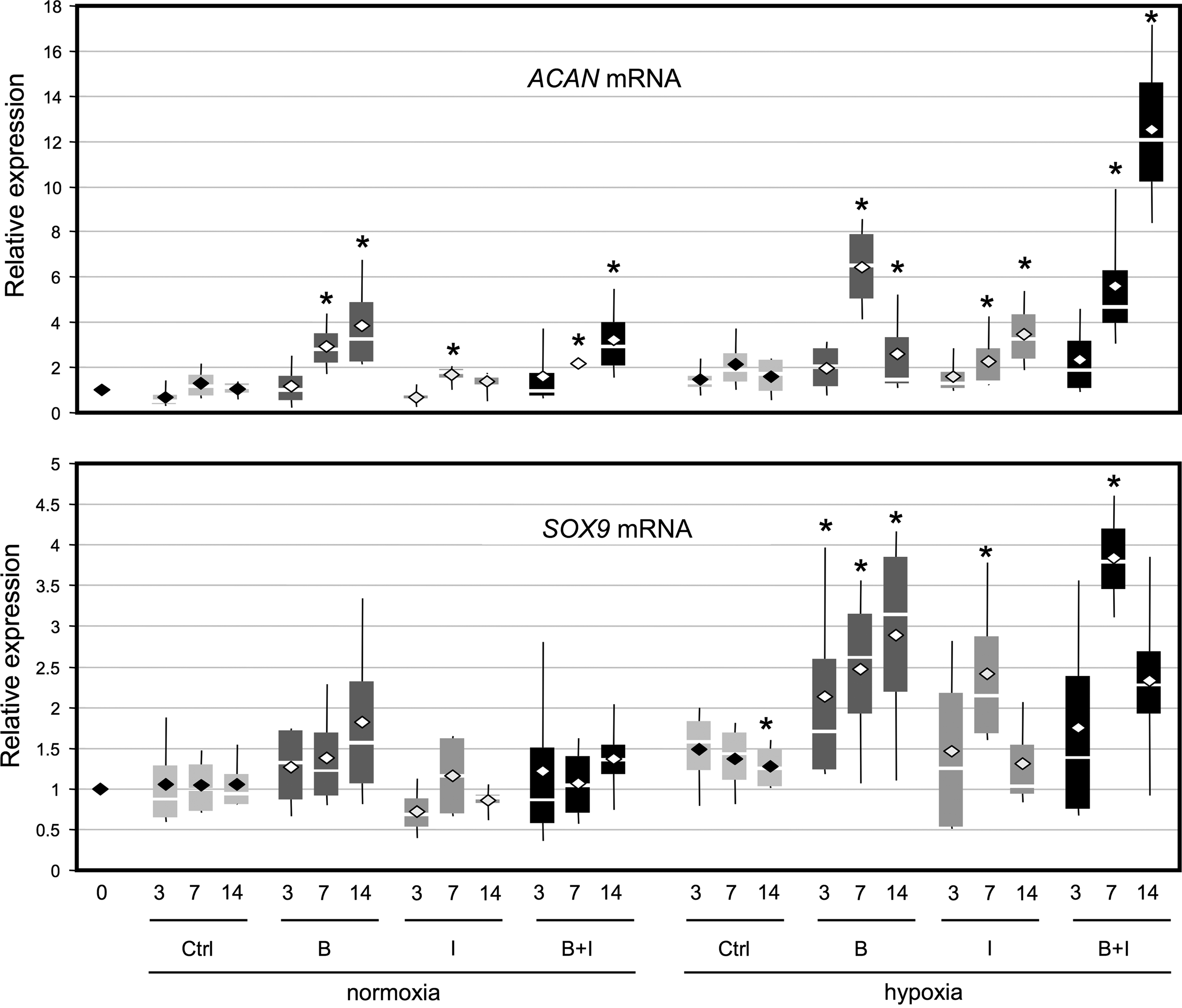
Effect of culture conditions on aggrecan and Sox9 mRNA expression. HACs were cultured in type I collagen sponges for 3, 7, and 14 days as described in Fig. 3. The levels of mRNA encoding aggrecan (ACAN mRNA) and Sox9 (SOX9 mRNA) were measured using real-time RT-PCR with specific primers. All the results were normalized to RPL13a mRNA, compared to untreated sponge-cultured cells at day 0, and are presented as the relative expression of each gene. Box plots represent four independent experiments performed in triplicate. Statistically significant differences between the untreated sponge-cultured cells at day 0 and treated cells were determined using the Mann–Whitney U test (*p<0.05).
Collagen sponge culture decreased COL1A1 mRNA steady-state levels during the first 3 days in normoxia as well as in hypoxia, but did not prevent their slight expression rise (Fig. 5). This expression profile was similar when chondrocytes were incubated with BMP-2 or BMP-2/IGF-I, whereas IGF-I accentuated this rise (with a significant increase at day 7 in normoxia and at day 14 in hypoxia). As a result, the COL2A1/COL1A1 ratio was improved in hypoxia compared to normoxia, with the best results obtained with the BMP-2/IGF-I combination or BMP-2 alone, 7 days after the beginning of the sponge culture (Fig. 5).
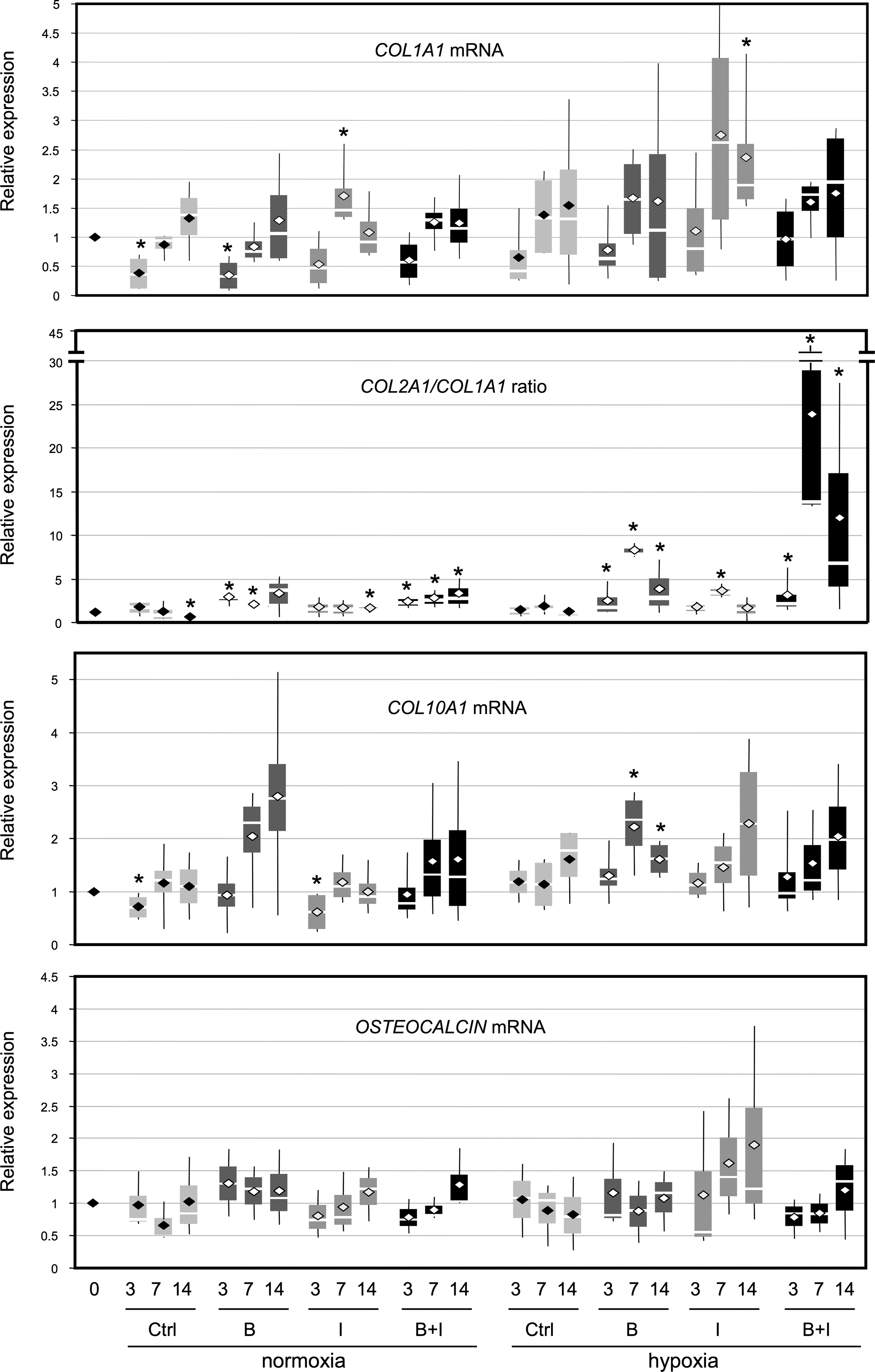
Effect of culture conditions on mRNA expression of nonspecific cartilage markers. HACs were cultured in type I collagen sponges for 3, 7, and 14 days as described in Fig. 3. The levels of mRNA encoding type I collagen (COL1A1 mRNA), type X collagen (COL10A1 mRNA), and osteocalcin (OSTEOCALCIN mRNA) were measured using real-time RT-PCR with specific primers. We also determined the COL2A1/COL1A1 ratio. Box plots represent four independent experiments performed in triplicate. Statistically significant differences between the untreated sponge-cultured cells at day 0 and treated cells were determined using the Mann–Whitney U test (*p<0.05).
Regarding steady-state levels of COL10A1 mRNA, oxygen tension had no influence on its expression when chondrocytes were cultured in sponge (Fig. 5). BMP-2, IGF-I, or both showed slightly increased COL10A1 mRNA levels, except in normoxia, where IGF-I had no significant effect between 7 and 14 days. However, chondrocytes cultured in normoxia or in hypoxia, did not initiate an osteoblastic maturation when seeded in collagen sponges: osteocalcin mRNA levels remained stable, even in the presence of BMP-2 or both BMP-2 and IGF-I (Fig. 5).
These results provide evidence that dedifferentiated chondrocytes, cultured in a collagen scaffold under low oxygen tension, become metabolically active when they were stimulated with BMP-2 or both BMP-2 and IGF-I and showed the best differentiated phenotypic profile after 7 days of culture, with synthesis of chondrocyte phenotype markers: type IIB collagen, aggrecan, and SOX9. Treatment with BMP-2 alone gave the best results: it specifically favored COL2B expression compared to COL2A, indicating that redifferentiated chondrocytes are obtained. However, low levels of COL1A1 mRNA expression persist, and COL10A1 is also enhanced by BMP-2, IGF-I, or both.
Furthermore, in the collagen sponge culture, in hypoxia or in normoxia, BMP-2 and IGF-I did not induce hypertrophy or osteogenesis.
Newly synthesized ECM proteins by chondrocytes cultured in type I/III scaffolds
We then monitored the chondrogenic effects of BMP-2 and IGF-I within the sponge culture at the protein level.
ELISA analysis of the sponge culture showed that the percentage of induction of type II collagen was higher in hypoxia than in normoxia (Fig. 6A). In contrast, the opposite effect was detected for type I collagen and, consequently, the induction ratio of type II collagen to type I collagen was enhanced in hypoxia compared to normoxia. In normoxia or in hypoxia, IGF-I was the strongest inducer of type I and type II collagen synthesis and the combination of IGF-I and BMP-2 had less of an effect than IGF-I alone. In normoxia, the induction ratio of type II collagen to type I collagen appeared to be better with IGF-I alone (but remained below 1), whereas in hypoxia, the best ratio was observed with BMP-2 alone (ratio >1.5). Regarding aggrecan, its percentage of induction was similar in normoxia and in hypoxia in basal conditions, whereas the addition of growth factors had a strong effect in hypoxia. In normoxia, only the combination of IGF-I and BMP-2 had inducing effects. In hypoxia, BMP-2 seemed to have the greater effect on aggrecan synthesis.
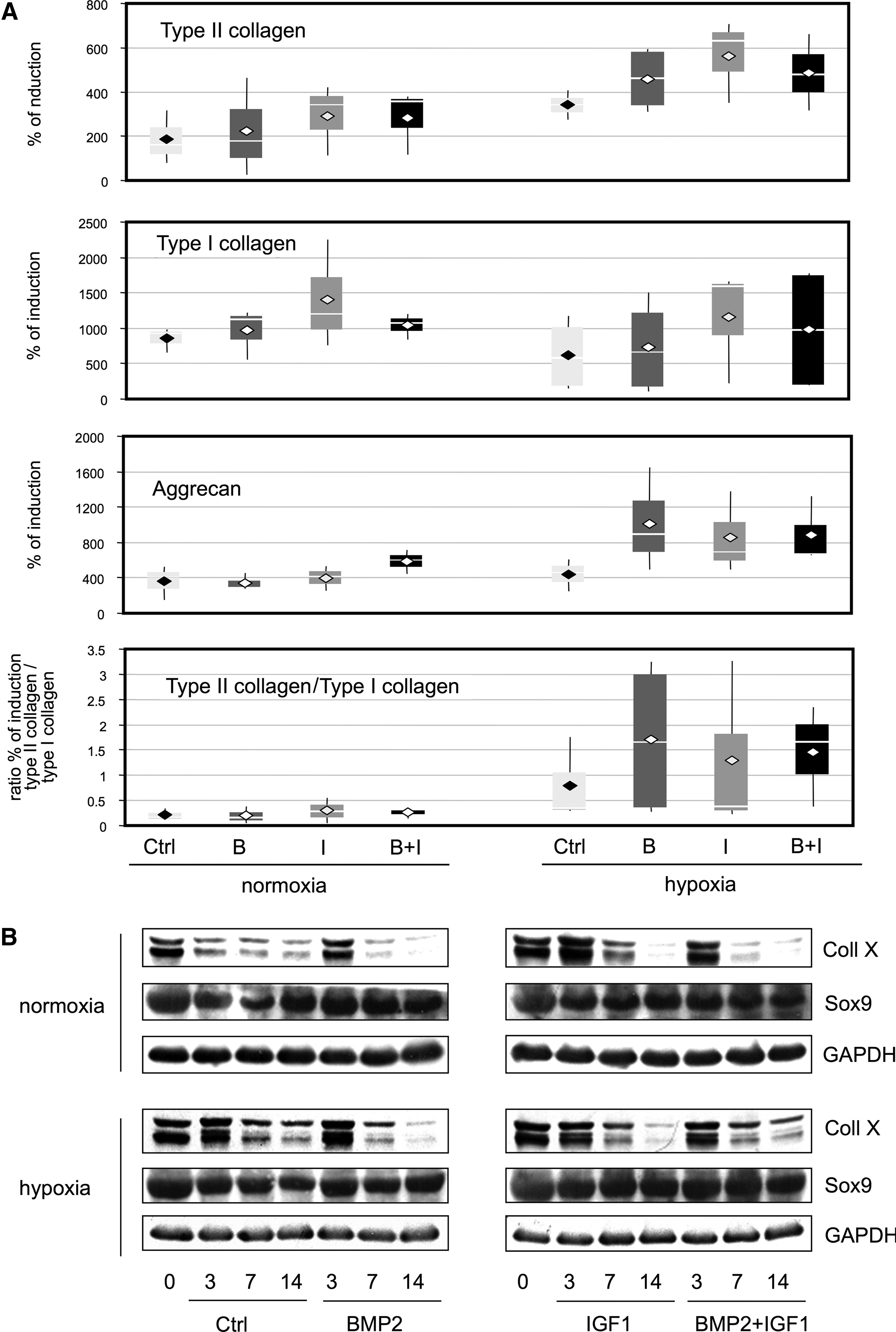
Effect of oxic conditions, collagen sponge culture, and BMP-2 and/or IGF-I treatments on the expression of collagenous proteins and the Sox9 protein. HACs were cultured in type I collagen sponges for 3, 7, and 14 days in normoxia or in hypoxia, in the absence (Ctrl), or in the presence of 50 ng/mL of BMP-2 (B), 10 ng/mL of IGF-I (I), or both BMP-2 and IGF-I (B+I).
The Western blot analysis showed that the two isoforms of type X collagen were regulated in the same way (Fig. 6B). Type X collagen levels from collagen sponge cultures gradually decreased over the 14 days of culture. In normoxia, BMP-2 and, especially, IGF-I treatments delayed this decrease. However, by day 7, the amount of type X collagen was decreased to below basal levels. In hypoxia, the BMP-2 and IGF-I treatments reduced the type X collagen expression as early as 7 days. The combination of BMP-2 and IGF-I had less effect than IGF-I alone. Furthermore, Sox9 expression increased in the BMP-2 treatment in normoxia, whereas it increased to a greater extent in the IGF-I treatment and the combination of two (IGF-I+BMP-2) in hypoxia.
These results confirm our analysis performed at the gene expression level and demonstrated that hypoxia greatly helped to actively redifferentiate dedifferentiated HACs. There was better induction of type II collagen in hypoxia and lowered type I collagen expression levels compared to those in normoxia. Both IGF-I and BMP-2 treatments induced aggrecan expression, but not type X collagen expression. Given that IGF-I was a stronger inducer of type II collagen and Sox9 expression, but also of type I collagen expression, BMP-2 appears to be the best candidate for redifferentiation experiments, and does not induce chondrocyte hypertrophy.
Type II collagen synthesis by newly redifferentiated chondrocytes
Immunofluorescence experiments were carried out to visualize collagen synthesis in the sponge cultures when chondrocytes presented a differentiated phenotype (Fig. 7). Beyond the slight background noise associated with type I collagen sponge culture autofluorescence, type I collagen was detected as early as dedifferentiated chondrocytes were seeded in the collagen sponge scaffold (day 0) until day 7 of culture, with or without BMP-2. In parallel, type II collagen was just perceptible at day 0 and at day 7, with stronger expression observed in the BMP-2 treatment. Given that type I collagen synthesis is a critical point in cartilage engineering, we adopted a strategy targeting type I collagen expression with a COL1A1 siRNA to abrogate its expression in our 3D experimental model (collagen sponge scaffold, hypoxia conditions, and BMP-2 treatment) to stabilize the differentiated chondrocyte phenotype.
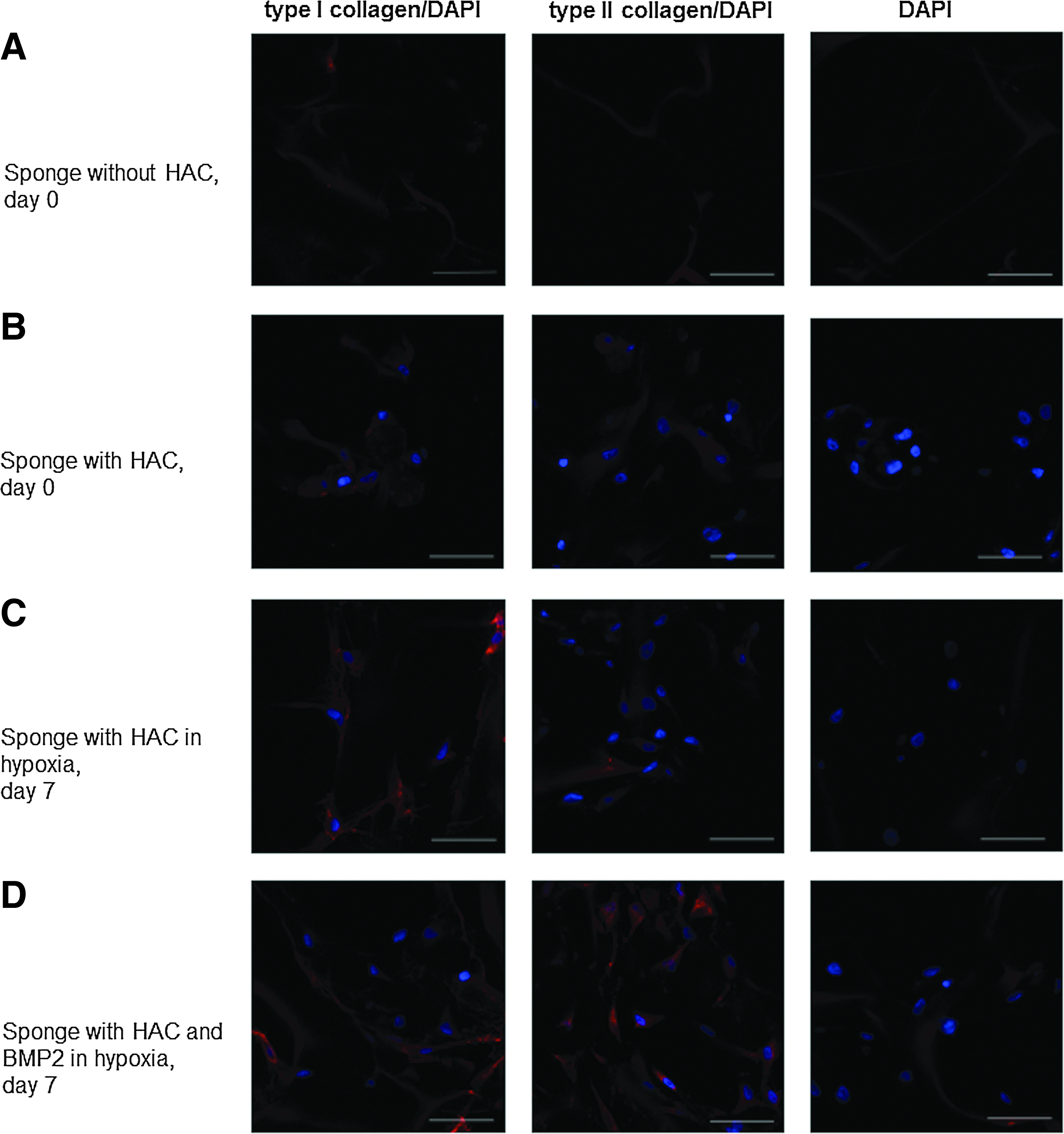
Immunostaining of type I and type II collagens secreted by HACs seeded onto sponges and incubated with BMP-2. HACs were cultured in type I collagen sponges for 7 days in hypoxia, with or without 50 ng/mL BMP-2. Samples were fixed on day 0
Improvement of the chondrocyte phenotype with COL1A1 siRNAs in the 3D model of culture
Nucleofected HACs with COL1A1 siRNA were seeded onto the collagen sponge scaffold and cultured for 24 or 48 h, in hypoxia, with or without BMP-2 (Fig. 8). The COL1A1 siRNA strongly decreased the type I collagen mRNA and protein amounts after 24 or 48 h of culture within collagen sponge scaffolds (from 50% to 80%). These results were also corroborated by Western blot analyses (data not shown). Moreover, the COL1A1 siRNA/BMP-2 combination enhanced the index of chondrocyte differentiation, evaluated by COL2A1/COL1A1 mRNA and type II/type I collagen protein ratios. Therefore, the use of COL1A1 siRNA substantially improves the differentiated chondrocyte phenotype essential for the synthesis of a hyaline cartilage-like matrix.
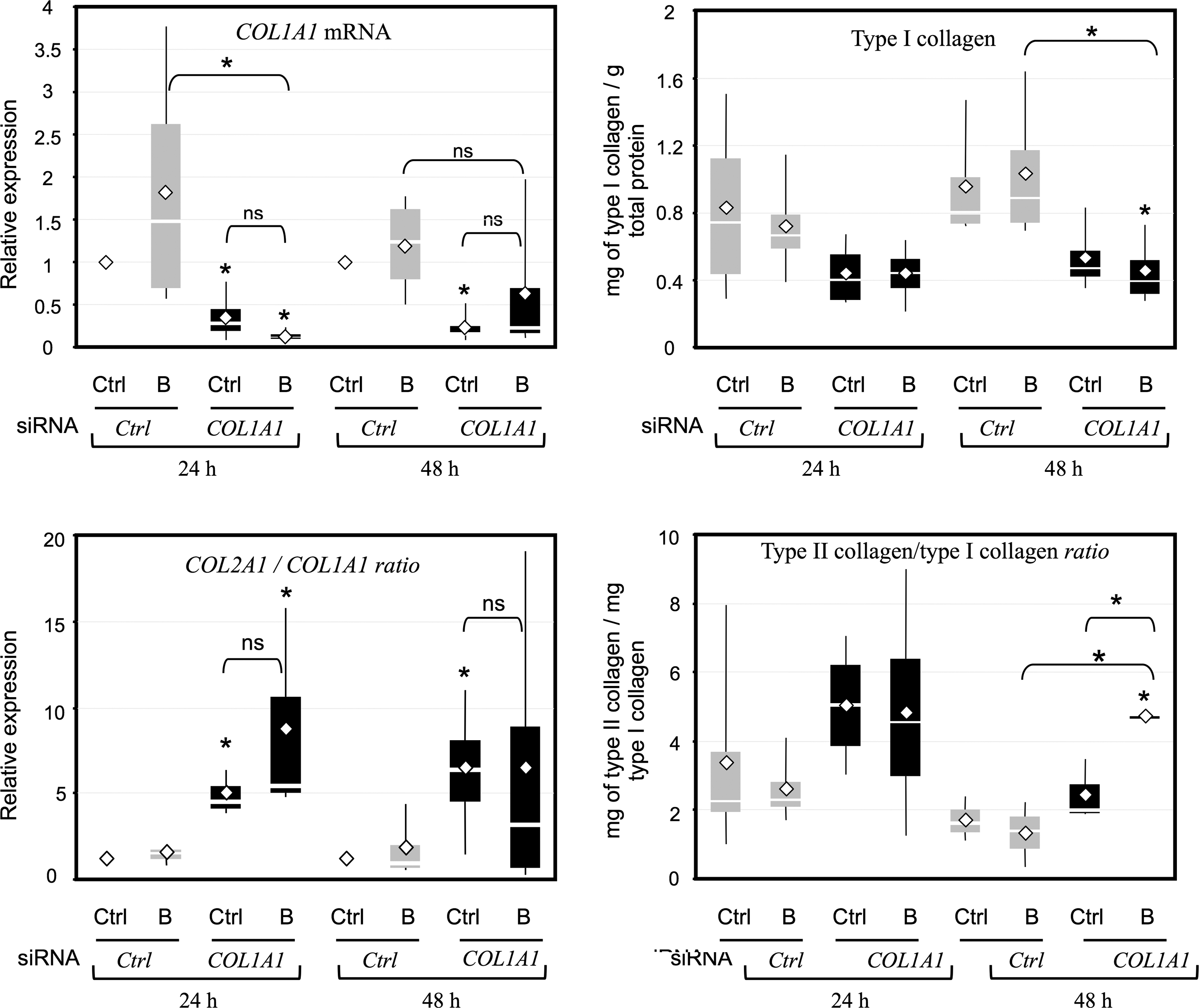
A COL1A1 siRNA enhances the differentiation index of HACs. HACs nucleofected with COL1A1 siRNA or control siRNA were seeded onto sponges and cultured for 24 or 48 h in hypoxia in the absence (Ctrl), or in the presence of 50 ng/mL of BMP-2 (B). Left panels: Steady-state levels of mRNA encoding type I collagen (COL1A1 mRNA) were measured using real-time RT-PCR with specific primers. Results were normalized to RPL13a mRNA, compared to control cells at 24 or 48 h and calculated as the relative expression of each gene. Then, for each set of experiments, we calculated the COL2A1/COL1A1 ratio. Results are presented as box plots of four independent experiments in triplicate. Right panels: Culture media were collected at 24 or 48 h. Protein quantification of type I collagen was performed using ELISA for each case of culture and normalized to total protein content. Results are expressed as milligram of collagen per gram of total protein content and are presented as box plots of four independent experiments. The ratio of type II collagen to type I collagen for each independent experiment was also calculated and is shown as box plots. Statistically significant differences between control cells at 24 or 48 h and treated cells were determined using the Mann–Whitney U test (*p<0.05), ns, not significant.
BMP-2 enhances the transcription activity of the human COL2A1 gene through its specific enhancer region
As transient transfection experiments require a substantial number of cells, they were first performed on RACs, and then validated on HACs, using human COL2A1 reporter gene constructs containing deletions in the promoter and/or first intron enhancer region. Experiments carried out first, on primary RACs showed that BMP-2 increased the transcription activity of the long construct pGL2—3.774 kb by 50% (Fig. 9A). For the other shorter constructs, transcriptional activation was lost, suggesting that BMP-2 trans-activating effects were mediated by a +2384/+2842-bp sequence. This region represents the greater part of the COL2A1 enhancer region located in the first intron (+2127/+2842). It includes the high-mobility group (HMG)-like binding site no. 2 (+2388/+2394) interacting with L-Sox-5 and Sox-6, the Sox-9-binding site (+2398/+2402), and the Sp1/Sp3/c-krox-binding sites (+2440/+2485 and +2817/+2842). BMP-2 treatments showed a decreased transcriptional activity in the pGL2-3.316 construct, which included the other part of this enhancer region (+2127/+2384), with the HMG-like binding sites no. 4 (+2361/+2367) and no. 3 (+2373/+2379) that bind L-Sox-5 and Sox-6. These data suggest that these two Sox binding sites are not sufficient by themselves to induce the BMP-2-trans-activation of COL2A1. The BMP-2 treatment also showed a reduced transcriptional activity of the pGL2-0.387 construct, including the Sp1/Sp3/c-krox-binding sites in the promoter. These cis-acting elements may be involved in BMP-2-negative regulation of COL2A1 mediated by the promoter. 22 In RACs after three passages, the BMP-2 treatment had no effect on the transcriptional activity of COL2A1, whatever the construct employed (Fig. 9B). This result suggests that BMP-2 transcriptional activation of COL2A1 was lost with the dedifferentiation of chondrocytes. The BMP-2 treatment induced a greater trans-activation of COL2A1 in HACs P1 than in RACs P0 (+74%), and this activation was also governed by the +2384/+2842-bp sequence of the intronic-specific enhancer (Fig. 9C). Sox9/L-Sox5/Sox6 and the zinc finger proteins c-Krox, Sp1, and Sp3 are known to be recruited on this region21,22,39,40 and are therefore involved in BMP-2 transcriptional regulation of the COL2A1 gene via this enhancer region.
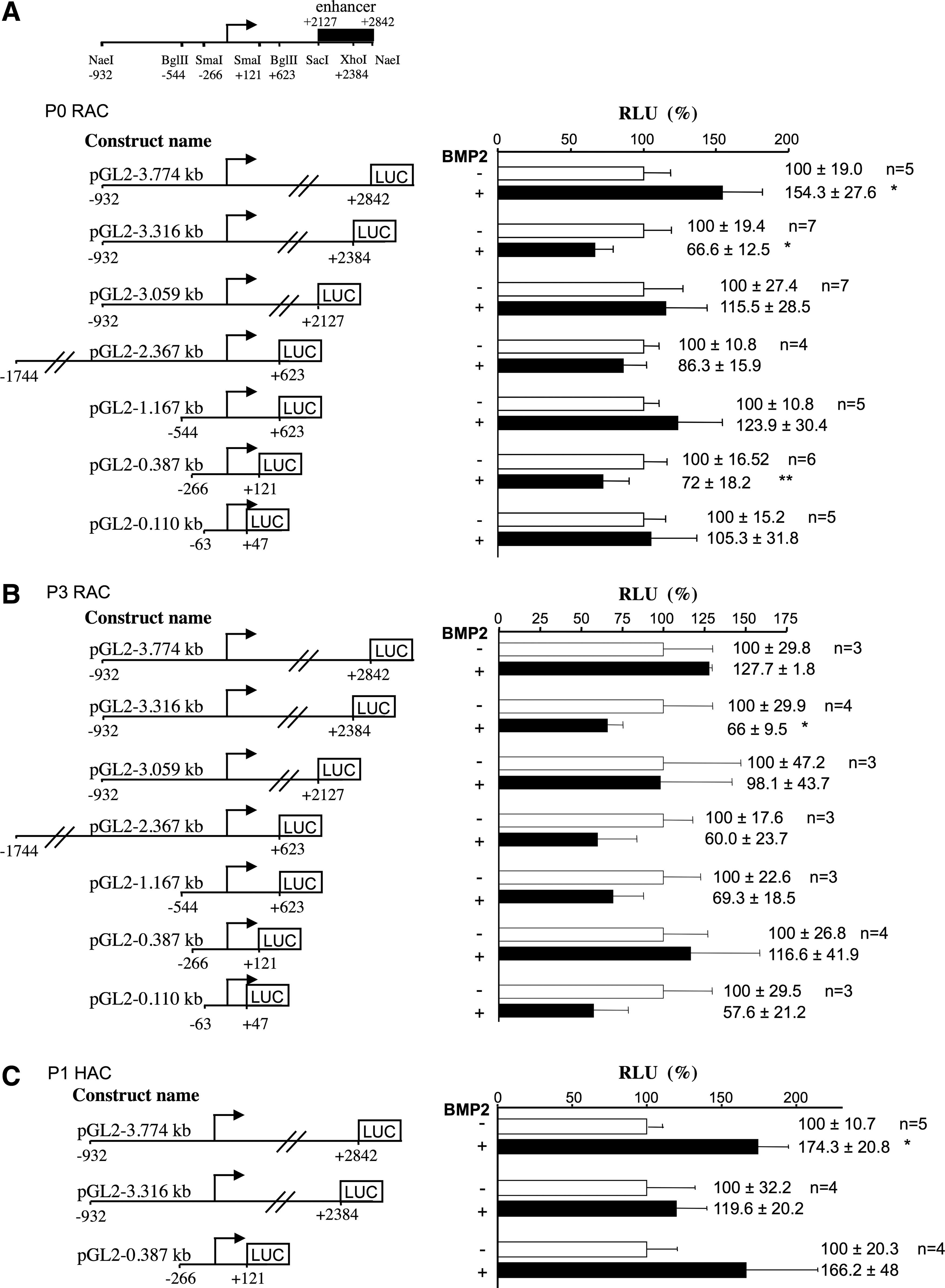
BMP-2-induced activation of human COL2A1 gene transcription is mediated by its first intron-specific enhancer region. Primary rabbit articular chondrocytes (RACs) [
In sponge-cultured chondrocytes in hypoxic conditions, BMP-2 increases the DNA-binding activity of the transactivators Sox9/Sox6/L-Sox5 and Sp1 on the specific enhancer of COL2A1, and decreased the binding of the transinhibitors of COL2A1, c-Krox, and NF-kappaB
We next examined the DNA-binding activity of Sox and Sp transcription factors on the COL2A1-specific enhancer in our 3D culture model (Fig. 10). First, IGF-I and BMP-2 had no effect on Sp1, Sp3, or c-krox gene expression in HACs cultured in a type I collagen sponge (Supplementary Fig. S1). Then, EMSA analyses were performed using probes for +2353/+2415α1(II) (four HMG cis-elements), +2383/ +2415α1(II) (HMG-like site no. 2 and Sox9 cis-element), and +2817/+2846α1(II) (Sp cis-elements) (Fig. 10). In normoxia, the BMP-2 treatment did not modulate the binding of Sox, particularly, Sox9 to the COL2A1 enhancer. In contrast, the IGF-I and the combined IGF-I and BMP-2 treatment increased the DNA binding of Sox9/L-Sox5/Sox6 and Sox 9 (Fig. 10A, B). In hypoxic cultures, BMP-2 and BMP-2 and IGF-I combined (at day 14), but not IGF-I alone, increased the DNA binding of Sox9/L-Sox5/Sox6 and Sox9 to their cis-acting elements in the COL2A1 enhancer. In normoxia, with the exception of IGF-I, neither BMP-2 nor BMP-2 combined with IGF-I, increased the DNA binding of Sp1 and Sp3 to the COL2A1 intronic enhancer (Fig. 10C). In hypoxia, at day 7, BMP-2 and/or IGF-I showed increased DNA binding of Sp1 and Sp3 to the COL2A1 enhancer.
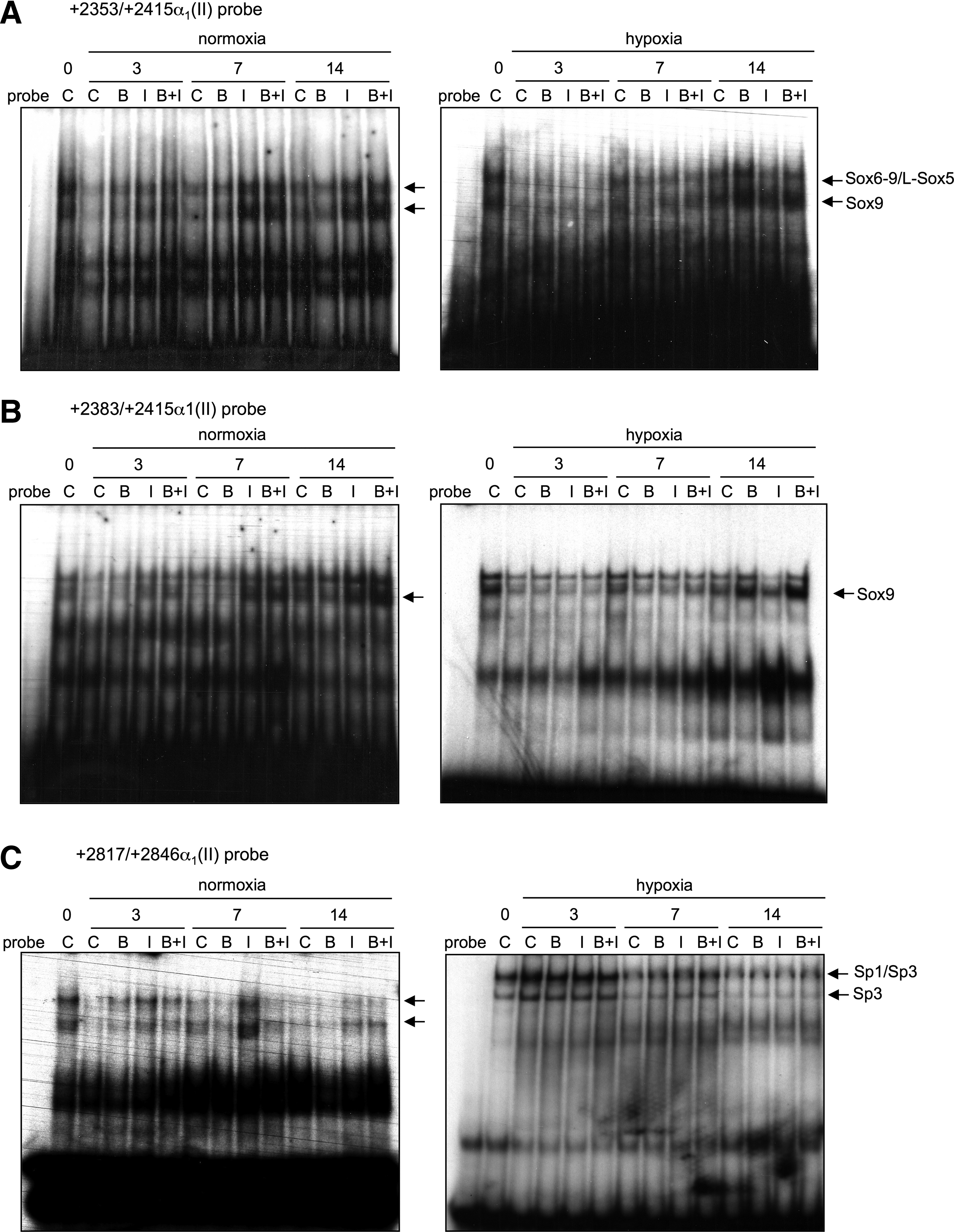
Characterization of the transcription factors implicated in IGF-I and BMP-2 transcriptional effects on COL2A1 gene in sponge-cultured HACs. HACs were cultured in type I collagen sponges for 14 days in normoxia or in hypoxia, in the absence (C), or in the presence of 50 ng/mL of BMP-2 (B), 10 ng/mL of IGF-I (I), or both BMP-2 and IGF-I (B+I). Nuclear extracts were prepared on days 0, 3, 7, and 14 and DNA binding was analyzed using electrophoretic mobility shift assays (EMSA). Labeled double-stranded +2353/+2415
Regarding the trans-inhibitors of COL2A1, c-Krox and NF-kappaB, their binding activity was evaluated using the −122/−61-bp probe of COL1A1 and a consensus probe, respectively. As shown in Figure 11A, whatever the oxic conditions and in the presence or absence of IGF-I, BMP-2 prevented the NF-kappaB DNA-binding activity. However, the decrease in binding was much more pronounced with BMP-2 alone. In contrast, IGF-I alone did not affect NF-kappaB binding in normoxia or in hypoxia except at day 7 in normoxia and at day 14 in hypoxia. Regarding c-Krox, which has been demonstrated to stimulate COL1A1 transcription in dermal fibroblasts 27 and to inhibit COL2A1 transcription in chondrocytes,21,41 its DNA binding was only weakly decreased by BMP-2 and was unaffected by IGF-I in normoxia (Fig. 11B). In hypoxia, there was a large decrease in the c-Krox-binding activity compared to normoxia, and BMP-2 further specifically reduced this binding at days 7 and 14. In contrast, as in normoxia, IGF-I did not affect the c-Krox-binding activity.
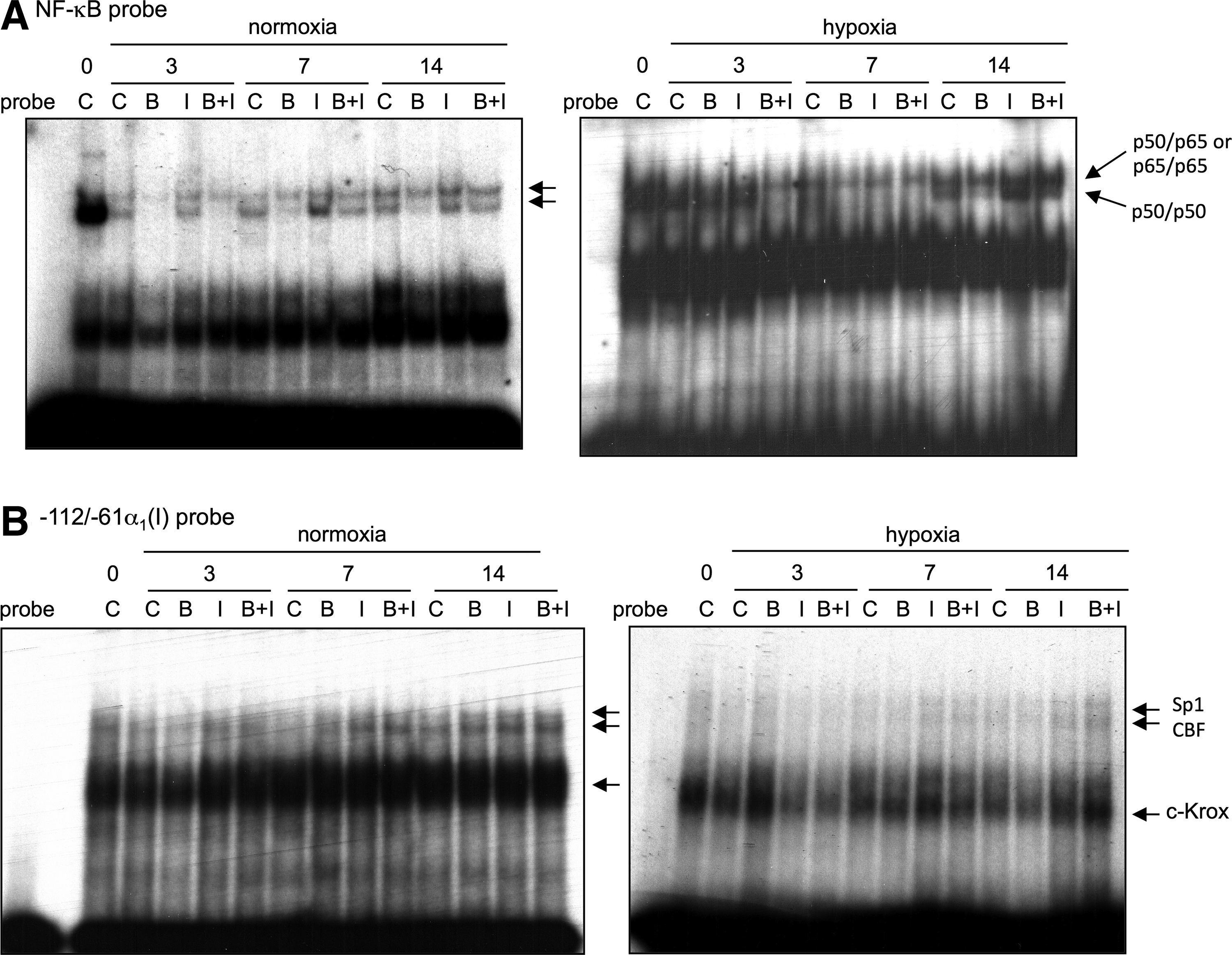
Effect of BMP-2 on the binding of trans-inhibitors of COL2A1 in sponge-cultured HACs. HACs were cultured in type I collagen sponges for 14 days in the same culture conditions as described in Fig. 10. Nuclear extracts were prepared on days 0, 3, 7, and 14 and DNA binding was analyzed using EMSA. A labeled double-stranded NF-kappaB probe
All together, these results suggest that BMP-2 induces upregulation of type II collagen in HACs cultured in hypoxia in type I collagen sponges, involve an increase in the binding and function of the trans-activators of the COL2A1 gene, that is, Sox9/L-Sox5/Sox6 and Sp1, recruited on its specific enhancer, and a concomitant decrease in the recruitment of the trans-inhibitors c-Krox and NF-kappaB on the COL2A1 gene.
Discussion
Cartilage matrix production in harvested chondrocytes is one of the paramount issues in cartilage engineering. In this study, we demonstrate that chondrocytes cultured in 3D collagen sponges under hypoxia with BMP-2 are metabolicaly active and synthesize a cartilage-like matrix improved with siRNA targeting COL1A1.
When trauma occurs or in OA, chondrocytes are in an inflammatory environment, with high cytokine (IL1-β, TNF-α, and IL-6) content. In our study, chondrocytes were derived from healthy femoral heads of patients undergoing total hip replacement, but nonetheless a proinflammatory cytokine environment. IL1-β plays a prominent role in cartilage catabolism, 42 concomitantly stimulating type I nonspecific collagen. This phenotype loss called dedifferentiation also occurs when primary chondrocytes are cultured as monolayers under 21% oxygen tension as soon as the first passage, a well-known mechanism previously described in our laboratory 8 and by others. 43
Although a great deal of interest has focused on growth and differentiation factors, such as the BMP family, we also monitored IGF-I, which has been demonstrated to stimulate COL2A1 transcription in RACs at 10 ng/mL. 21 In our study, we found that IGF-I induced COL2A1 and ACAN expression, like others,44,45 but also COL1A1, COL10A1, and Osteocalcin, which are matrix molecules inappropriate for cartilage engineering, and which indicates that IGF-I has a nonspecific effect. Then, the chondrocytes cultured with IGF-I are not well-differentiated. As TGF-β1 is also recognized as a potent inducer of redifferentiation of articular chondrocytes in hypoxia, the combinatory effect of TGF-β1 and IGF-I could be also an attractive issue as described recently by Jonitz et al. 46
In our study, BMP-2 appears to be a much better candidate for redifferentiation experiments, because unlike IGF-I, it favored type II collagen expression rather than type I collagen expression and especially type IIB collagen rather than the type IIA isoform. This cartilage-specific type IIB collagen re-expression was shown in mouse embryonic chondrocytes. 47 As in the study performed on human nasal chondrocytes, 17 our data confirm that BMP-2 (50 ng/mL) did not particularly enhance type I collagen expression. Likewise, BMP-2 does not affect the expression of another procollagen type I encoding gene (Col1a2) in equine articular chondrocytes. 19 This chondrogenic effect with low concentrations of BMP-2 was confirmed by the fact that BMP-2 did not upregulate chondrocyte hypertrophy. Since cells, used in this study, were obtained from surgically discarded tissues with various degrees of degeneration, HACs synthesized type X collagen at day 0. Nevertheless, collagen sponge culture drastically decreased type X collagen synthesis and the addition of BMP-2 and IGF-I accelerated this reduction at the protein level. Furthermore, BMP-2, used at a low concentration did not promote osteogenic phenotype in HACs, as showed in human intervertebral disc cells. 48
Several studies have used the combination of BMP-2 and IGF-I on chondrocytes and MSCs to promote cartilage formation.49,50 For instance, BMP-2, insulin, and IGF-I were identified as the most potent factors for GAG accumulation among 12 putative chondrocyte regulators, but the combination of BMP-2 and insulin was the most effective for the redifferentiation of dedifferentiated chondrocytes after repeated passages. 14 Here, the combination of BMP-2 and IGF-I did not improve the chondrocyte phenotype, insofar as it favored, once again, type II collagen rather than type I collagen, but it did not increase the COL2B/COL2A ratio or aggrecan expression as BMP-2 alone did.
Therefore, the best chondrocyte phenotype was observed when the chondrocytes were cultured in the biodegradable collagen sponge scaffold under hypoxia with BMP-2. It is well known that a 3D environment is beneficial for the restoration of the chondrocyte phenotype.35,51 Of the available scaffolds, type I collagen sponges were chosen for their chondrogenic potential5,52 and their compatibility with clinical practice. Compared to type I collagen gels, atelocollagen sponges are less contracted and are made of collagen from which the telopeptide-containing determinants have been removed. Thus, matrikine release is prevented and the risk of collagen-induced arthritis is avoided. Here, type I collagen sponges allow rounded cell attachment and chondrocyte-specific gene expression when chondrocytes are cultured in hypoxia with BMP-2. These scaffolds, already used for human skin regeneration, have now been demonstrated to be suitable for cartilage repair.
Low oxygen tension was another environmental parameter that exacerbated redifferentiation effects in the sponge cultures. Several research groups have used hypoxia as well as 3D cultures (alginate) to redifferentiate chondrocytes,53,54 but to our knowledge, no study has reported the combination of hypoxia and collagen sponge scaffolds. We showed that hypoxia increases type II collagen and aggrecan expression and decreases type I collagen expression. These effects have previously been described and are mediated by the HIF pathway and related to Sox9 induction.8,55 In our experimental conditions, the Sox9 activity appeared to be modulated by the addition of growth factors rather than by oxygen tension, and we did not explore the upstream mechanisms through which hypoxia improves the chondrocyte phenotype.
We set out to generate a functional cartilage-like tissue in vitro to test its potential as a biological substitute that can be implanted in joints. The association of HACs, BMP-2, hypoxia, and a type I/III collagen sponge scaffold is better suited to this objective, but a non-negligible background of type I collagen synthesis remains. We successfully employed a siRNA strategy directly targeting the COL1A1 gene of HACs, which were then cultured in sponges. Thus, the association of COL1A1 siRNA and BMP-2 favored chondrocyte differentiation in hypoxia. Others have used an adenoviral vector encoding both TGF-β3 and type I collagen shRNA to control the chondrogenesis of porcine chondrocytes. 56 This recombinant genetic technique (genetic therapy) leads to chondrocyte redifferentiation and corroborates the potential of RNA interference in cartilage engineering. However, it is not suitable for clinical applications and the transitory action of siRNA transient transfection allows the production of safe, competent therapeutic cells in a cell therapy strategy. Therefore, we first focused on the direct implantation of transfected cells into the scaffold. In a second step, in view of the in vivo implantation of our model, we also developed an effective strategy for the delivery of siRNA in sponge-cultured chondrocytes during 7 days, using an effective transfection reagent (data not shown).
We took our study one step further and explored the molecular mechanisms behind the BMP-2 enhancement of the chondrocyte phenotype. In our 3D culture, IGF-I increased the DNA binding of Sox9, Sox6-9/L-Sox5, Sp1, and Sp1/Sp3 to the COL2A1 intronic enhancer only in normoxia. These results are in accordance with our previous results obtained using RACs. 21 One study showed that BMP-2 transcriptional activation of proα1(II) collagen gene expression takes place through a 48-bp sequence located in the chondrocyte-specific enhancer of its first intron in the pluripotent mesenchymal cell line C3H10T1/2. 57 This sequence has already been characterized and is required for COL2A1 cartilage-specific expression in mice.24,40 A homolog enhancer sequence is present in the first intron of human genes (+2127/+2842). 32 In the present work, we demonstrated that BMP-2-trans-activation was mediated by the +2384/+2842-bp sequence of the COL2A1-specific enhancer in P0 RACs and in P1 HACs. Since Sox9/L-Sox5/Sox6 and the zinc finger proteins c-krox, Sp1, and Sp3 are recruited on this region,27,45,47 we hypothesized that they are involved in BMP-2 trancriptional regulation of the COL2A1 gene. More precisely, three HMG-like binding sites (+2361/+2367; +2373/+2379; +2388/+2394), a Sox-9-binding site (+2398/+2402), and Sp1/Sp3/c-krox-binding sites (+2440/+2485 and +2817/+2842) have previously been characterized in this region. The presence of the first two HMG-like binding sites is not sufficient to induce BMP-2 transciptional activity, but they can collaborate with other transcription factors in BMP-2 trans-activation. BMP-2 has no effect on the transcriptional activity of COL2A1 in P3 RACs, suggesting that BMP-2 transcriptional activation of COL2A1 is lost in dedifferentiated RACs, but not in dedifferentiated HACs: we used dedifferentiated HACs from arthritic patients. Furthermore, BMP-2 also maintains its effect on COL2A1 gene expression at passage 3 in human nasal chondrocytes. 17 Here, we demonstrated that BMP-2 increases the DNA binding of Sox9, Sox6-9/L-Sox5, Sp1, and Sp1/Sp3 to the COL2A1 intronic enhancer when chondrocytes are cultured in a collagen sponge scaffold, but only in hypoxia, suggesting that these transcription factors are involved in BMP-2 effects. In support of this finding, Sp1 has been shown to have a role in BMP-2 upregulation of the Erk2 gene 58 and several studies have implicated the three Sox transcription factors in nasal human chondrocytes, and particularly Sox6 in the C3H10T1/2 cell line.17,57 In hypoxia, we observed that BMP-2 induced Sox9 mRNA and upregulation of the three Sox transcription factors have previously been observed after BMP-2 treatment in various types of cells.59,60
Our studies developed on human dermal fibroblasts have shown that c-Krox, Sp1, and Sp3 are key trans-activators of COL1A1 transcription, whereas NF-kappaB, a factor involved in oxidative and aging/senescence processes, is a trans-inhibitor.27,30 These factors are also expressed in chondrocytes and we suspect that some of them are involved in COL1A1 trans-activation because dedifferentiation occurs in HACs grown in vitro. Moreover, chondrocyte dedifferentiation is characterized by a decrease in the expression and of the DNA-binding activity of the trans-activators of COL2A1, such as Sp1 and Sox9 and stabilization or an increase in the trans-inhibitors Sp3 and NF-kappaB, respectively. The p65 subunit of NF-kappaB was found to inhibit COL2A1 transcription in chondrocytes through the −63-bp promoter (data not shown). Our present data demonstrate that the RNA interference strategy targeting COL1A1, even though it is efficient for chondrocyte phenotype recovery, can be improved with the additive effects of BMP-2. BMP-2 can increase the binding and function of the transcription activators (the Sox transcription factors and Sp1) and concomitantly inhibit the trans-inhibitors of COL2A1 (c-Krox and NF-kappaB). Nevertheless, more investigations are necessary to better understand the molecular mechanisms behind the BMP-2 process in our experimental conditions and which cofactors are involved. In a previous work, we demonstrated that the BMP-2 effect is governed, at least in part, by an interaction of Sox9 with Sp1 and p300. 21
In conclusion, we demonstrated that dedifferentiated HACs can reactivate their redifferentiation program when cultured in a 3D scaffold in hypoxic conditions with BMP-2 and a COL1A1 siRNA. However, it will be necessary to culture the cells within a perfusion bioreactor, with or without physical stimulation, to standardize culture conditions to control cartilage regeneration. It would also be interesting to evaluate the behavior of this therapeutic model in competent cells in vivo. The subcutaneous implantation in the nude mouse model will help to determine if a hyaline-like or a fibro-cartilage is formed. This study therefore provides new insights into the molecular mechanism of stability and/or induction of the chondrocyte phenotype, an absolutely necessary step for developing new methods of ACI therapy in humans.
Footnotes
Acknowledgments
This work was supported by the Regional Council of Lower Normandy, the ANR (Agence Nationale de la Recherche) TecSan (Technologies de la Santé) program (PROMOCART 2006) and FEDER (“Fonds structurels européens”) grant HIPPOCART 1, n° 33535. F.L.: Post-doctoral fellow supported by a PROMOCART grant (Health Technology) from the French National Agency of Research (ANR). D.O. and L.M.: Fellowship from the French Ministry of Research and Technology. M.H.: Fellowship from the Regional Council of Lower Normandy.
Disclosure Statement
No conflict of interests is declared by all the authors.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.