
Abstract
Resolution of intervertebral disc (IVD) degeneration-associated inflammation is a prerequisite for tissue regeneration and could possibly be achieved by strategies ranging from pharmacological to cell-based therapies. In this study, a proinflammatory disc organ culture model was established. Bovine caudal disc punches were needle punctured and additionally stimulated with lipopolysaccharide (10 μg/mL) or interleukin-1β (IL-1β, 10–100 ng/mL) for 48 h. Two intradiscal therapeutic approaches were tested: (i) a nonsteroidal anti-inflammatory drug, diclofenac (Df) and (ii) human mesenchymal stem/stromal cells (MSCs) embedded in an albumin/hyaluronan hydrogel. IL-1β-treated disc organ cultures showed a statistically significant upregulation of proinflammatory markers (IL-6, IL-8, prostaglandin E2 [PGE2]) and metalloproteases (MMP1, MMP3) expression, while extracellular matrix (ECM) proteins (collagen II, aggrecan) were significantly downregulated. The injection of the anti-inflammatory drug, Df, was able to reduce the levels of proinflammatory cytokines and MMPs and surprisingly increase ECM protein levels. These results point the intradiscal application of anti-inflammatory drugs as promising therapeutics for disc degeneration. In parallel, the immunomodulatory role of MSCs on this model was also evaluated. Although a slight downregulation of IL-6 and IL-8 expression could be found, the variability among the five donors tested was high, suggesting that the beneficial effect of these cells on disc degeneration needs to be further evaluated. The proinflammatory/degenerative IVD organ culture model established can be considered a suitable approach for testing novel therapeutic drugs, thus reducing the number of animals in in vivo experimentation. Moreover, this model can be used to address the cellular and molecular mechanisms that regulate inflammation in the IVD and their implications in tissue degeneration.
Introduction
D
The degenerated IVD is characterized by cell death and changes in matrix composition, being the nucleus pulposus (NP) subjected to high mechanical and osmotic pressures, severe hypoxia, and limited nutrient supply.7–9 In this process, increased levels of nitric oxide and prostaglandin E2 (PGE2), as well as upregulation of metalloproteases (MMPs) and a wide number of inflammatory mediators (e.g., tumor necrosis factor-α [TNF-α], interleukin-1β [IL-1β], IL-6), have also been observed.3,5,10–13 Ultimately, structural damage of the outer annulus provides an opportunity for blood vessels and nerves to invade the disc and cause pain.10,14
The high impact on population health and the lack of adequate solutions in the clinics stimulate the development of novel IVD biological therapies, 15 with the goal to promote IVD regeneration and/or control inflammation-associated pain.8,16,17 However, the question of how inflammation can be related with IVD degeneration is still controversial and has been described differently from in vitro experiments to in vivo animal models. 18
Nevertheless, there is a lack of adequate models to study inflammation within IVD degeneration. IVD cells in vitro lose their ability to produce IVD native extracellular matrix (ECM). 19 In vivo models of disc injury by puncture alone (Punct) or NP digestion do not mimic the natural process of human IVD degeneration.20,21 Thus, ex vivo organ culture models using disc explants from different species have been established for studying disc degeneration in a more physiologically relevant environment. 21 Explant cultures of bovine discs have the advantage of easy availability and they are assumed to be suitable ex vivo models for studying therapies of disc degeneration as they allow well-controlled environmental conditions21,22 and show high similarities with human samples regarding the induction of a degenerative environment. 23
The aim of the present study was to establish a bovine organ culture model in a proinflammatory environment.
The control of inflammation by intradiscal injection of a nonsteroidal anti-inflammatory drug (NSAID), (diclofenac [Df]), was then tested and used to validate this model. NSAIDs are known as the most effective anti-inflammatory agents in the market, 24 but their systemic administration presents drawbacks such as short biological half-life, rapid metabolism, and high percentage of protein binding. These characteristics lead to the use of high doses in patients, which causes side effects in the gastrointestinal, hepatorenal, and cardiac systems.25,26 Intradiscal administration of these drugs could overcome some of the drug systemic side effects.
Furthermore, to analyze if mesenchymal stem/stromal cells (MSCs)-based therapy could have an impact in the reduction of inflammation in degenerated IVD, MSCs embedded in a human serum albumin/hyaluronan (HSA-HA) hydrogel 27 were injected in the same model. MSCs have been suggested to be an adequate cell source, being capable of differentiating toward an NP-like phenotype in coculture with NP cells28–31 and promoting the production of healthy ECM in NP in vivo.32,33 However, although animal model studies have reported that MSCs can promote IVD regeneration,20,34 less is known about the possible beneficial effect of MSCs in the modulation/resolution of inflammation in IVD. The expression of Fas ligand (FasL), a protein found in immune-privileged sites, has been restored in the IVD upon implantation of MSCs. 35 Human MSCs were able to downregulate the gene expression of proinflammatory cytokines (IL-3, IL-6, IL-11, IL-15, TNF-α) and MMPs produced by rat NP cells 36 and also reduce IgG production from human NP fragments. 14 In human trials, patients referred reduction of pain,37,38 but the influence of MSCs on inflammation was poorly investigated in the context of disc degeneration. Therefore, we investigated in our organ culture approach if MSCs injection into a proinflammatory disc environment has the capacity to control the inflammation response.
Materials and Methods
Establishment of a bovine organ culture model
Bovine IVD tissue was isolated from tails of young adult animals (age <48 months old) from a local slaughterhouse (in Germany by the local abattoir Ulmer Fleisch GmbH and in Portugal with the ethical approval of the Portuguese National Authority for Animal Health). Within 3 h after slaughter, up to six caudal discs from each specimen were isolated by removal of skin, muscles, and ligaments. Discs were dissected from the adjacent vertebral bodies as close as possible to the upper and lower cartilaginous end plates (Fig. 1A, B). 39 Standardized punches (diameter of 13 mm) were prepared from each disc with the NP in the center and few surrounding annulus lamellae (Fig. 1C, D).
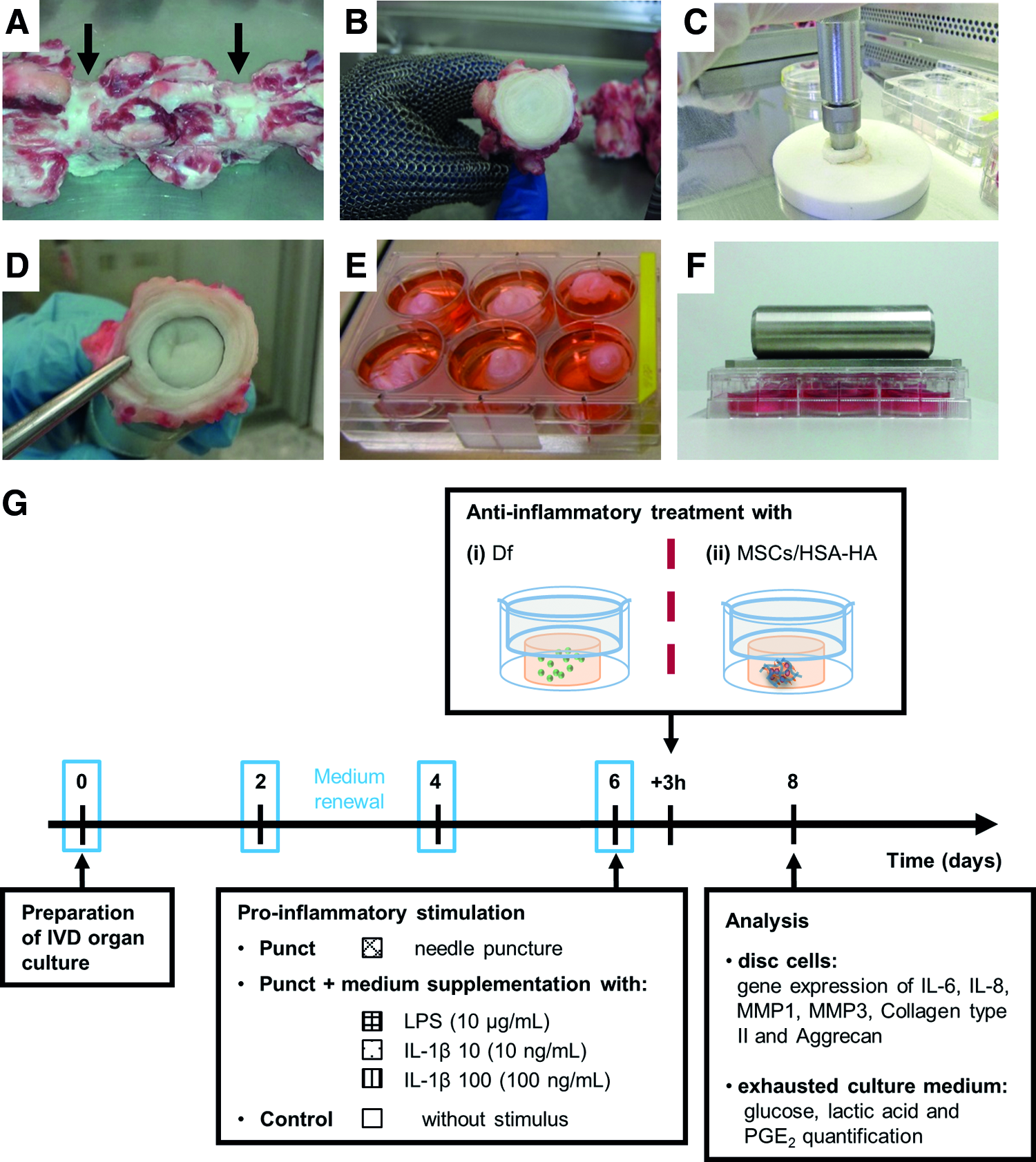
Organ culture preparation.
In pre-experiments, constrained and unconstrained conditions were compared (Supplementary Materials and Methods; Supplementary data are available online at
To compare constrained and unconstrained conditions, macroscopic parameters of disc punches were determined by measurement of disc height, diameter, and wet weight after 14 days of culture in BM. Biochemical characterization was performed by quantification of glycosaminoglycans (GAG) release over a culture period of 5 weeks. In addition, alcian blue staining of IVD sections was performed at different time points. For all the following experiments, constrained conditions were used.
Simulation of proinflammatory environment
For simulation of the proinflammatory/degenerative environment, after 6 days of culture in BM, IVD organ cultures were injured by needle Punct 40 or also stimulated with proinflammatory factors: lipopolysaccharide (LPS)41,42 or IL-1β.43,44 Briefly, samples were punctured with a sterile 21G needle; discs were oriented with sterile forceps, while the needle was inserted laterally throughout the remaining annulus fibrosus into the NP and rotated clockwise for 30 s. Discs were cultured in BM supplemented with bacterial (Escherichia coli) LPS (10 μg/mL; Sigma-Aldrich) or recombinant human IL-1β (10 or 100 ng/mL; R&D Systems) according to Figure 1G. Nonpunctured untreated discs (only cultured in BM) were used as controls. Tissue samples were collected after 48 h of proinflammatory treatment for cell viability and gene expression analysis. The metabolic activity analysis and PGE2 quantification were performed in culture supernatants.
Evaluation of Df injection in the proinflammatory IVD organ culture
Three hours after proinflammatory stimulus, discs were treated with an injection of 500 μL of Df in solution (19 μM; Sigma-Aldrich) using a microsyringe and a 33G needle (Hamilton). The time point for Df treatment was selected based on our previous work with in vitro studies of an anti-inflammatory treatment of activated macrophages. 45 Nonmanipulated samples cultured only in BM were used as controls. The effects were evaluated 2 days later by gene expression and by PGE2 production. The experimental scheme is represented in Figure 1G.
Culture of human MSCs
Human MSCs harvested from bone marrow were obtained from five different donors who underwent knee joint surgery with informed consent and according to the rules of the ethical commission of the University of Ulm (Ulm, Germany). One patient was a healthy young donor for bone marrow transplantation. MSCs phenotype was tested immunohistochemically by CD9, CD90, CD105, CD44, and Stro-1 staining. In addition, the cells were confirmed to be able to differentiate into osteogenic, chondrogenic, and adipogenic lineages as described. 46 All cells were expanded in DMEM (Biochrom) supplemented with 10% v/v FBS (HyClone; Thermo Scientific), 1% v/v penicillin/streptomycin (10,000 U/mL–10,000 μg/mL; Biochrom), and 0.5% v/v fungizone (Biochrom). Cells were seeded at a concentration of 3000 cells/cm2 and expanded in T-flasks at 37°C, under a humidified atmosphere of 5% v/v CO2 in air, with the culture medium being changed twice a week and trypsinized when reaching 70% confluence. Experiments were performed with MSCs in passage 2.
MSCs injection in the proinflammatory IVD organ culture
A hydrogel of albumin–hyaluronan was selected as a carrier system for MSCs.27,47 MSCs/HSA-HA (500 μL) with a cell concentration of 2 × 106 MSCs per milliliter of hydrogel were injected in the IVD in culture using a double-chamber syringe (Medmix Systems AG) and a 21G needle. The cell-containing gel mixture was filled in the larger chamber of a two-chamber syringe and the cross-linker in the smaller one. The gel was polymerized in situ.
Sample preparation for quantitative real-time reverse transcription–polymerase chain reaction
Gene expression levels were determined by quantitative real-time reverse transcription–polymerase chain reaction (qRT-PCR) performed in triplicate on cDNA derived from disc samples. NP samples were digested enzymatically, cell pellets were recovered, and total RNA was isolated, quantified, and transcribed into cDNA.
Briefly, each tissue sample was dissected into 2–3 mm3 fragments and enzymatically digested for 2 h in 0.8 mg/mL collagenase type I (Sigma-Aldrich) in DMEM, under agitation (50 rpm), and reduced oxygen atmosphere (37°C, 6% O2 and 8.5% CO2) and saturated humidity. Supernatant was passed through a 40-μm filter (BD Falcon) to remove tissue debris. Cells were collected by centrifugation at 400 g for 7 min. Total RNA was extracted from disc punch cells, using ReliaPrep RNA Cell Miniprep System (Promega), according to manufacturer's instructions. RNA was quantified using a NanoQuant spectrophotometer (Infinite M200; Tecan). Quality was checked by means of an RNA ratio, pooled from two disc samples for each condition. Of each RNA pool, 2 μg was reverse transcribed into cDNA using the Omniscript RT Kit (Qiagen) completed with oligodeoxythymidine primers (5 μM), random hexamer primers (50 μM), and RNase inhibitor (10 U) in a total volume of 20 μL. The obtained cDNA was diluted at a ratio of 1:4 in RNase-free water (Qiagen) and used for qRT-PCR.
Quantitative real-time reverse transcription–polymerase chain reaction
Specific primer pairs were designed using published gene sequences (PubMed, NCBI Entrez Nucleotide Database) and Primer 3 software 48 for bovine IL-6, IL-8, MMP1, MMP3, collagen II (Coll II), aggrecan (Agg), and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (Table 1) and synthesized by Thermo Fisher Scientific. The analysis was carried out using SYBR® Green method. Reactions were conducted on StepOnePlus Real-Time PCR System (Applied Biosystems), in triplicate, in PCR 96-well TW-MT-Plates (Biozym Scientific), under standard conditions. Reaction mixes contained 12.5 μL of Platinum SYBR Green qPCR SuperMix-UDG (Invitrogen) master mix, 0.25 μL of ROX Reference Dye (Invitrogen), 1 μL (0.4 μM) of forward primer, 1 μL (0.4 μM) of reverse primer, 8.25 μL of RNase-free water, and 2 μL of cDNA. For the analysis of the mRNA expression, cloned amplification products were provided and used as standards for qRT-PCR. Statistical analysis was performed on ΔCt values according to a modified method described by MacLean et al. 49 Fold changes in gene expression were presented as 2−(averageΔΔCt). The average Ct value of each triplicate measurement of each sample was normalized to the house-keeping gene GAPDH in each sample [ΔCt = Ct(gene of interest) − Ct(GAPDH)]. The ΔCt of each stimulated sample was related to the respective ΔCt of each control sample. Normalized values of samples collected at the end of the experiments were compared with the control and between the different experimental groups.
Bovine oligonucleotide primers used for qRT-PCR.
GAPDH, glyceraldehyde 3-phosphate dehydrogenase; IL, interleukin; MMP, metalloprotease; qRT-PCR, quantitative real-time reverse transcription–polymerase chain reaction.
Statistical analysis
Since the data follow a nonparametric distribution, for qRT-PCR, Mann–Whitney and Kruskal–Wallis tests were used to compare two or several groups, respectively. Statistical analysis was performed using GraphPad Prism vs. 6.0 for Windows (vs. 6.01), with values of p < 0.05 considered significant.
Results
Cell viability and metabolic activity in a bovine IVD organ culture model in proinflammatory conditions
Disc punches were prepared as described and cultured at 5 mM glucose, 400 mOsm, 8.5% CO2, and 6% O2 under constrained conditions. Discs were constrained with membrane inserts posed on top of each disc punch covered by a weight on the top of the culture plate (simulating 0.5 MPa static loading, which corresponds to physiological loads during the standing phase 50 ) and allowed the prevention of disc swelling and tissue deformation (Supplementary Results) that occurred without end plates (Supplementary Fig. S1). In these conditions, tissue GAG slightly decrease (Supplementary Fig. S2).
To induce a degenerative and proinflammatory environment, cultures were stimulated at day 6 after isolation with Punct or puncture with supplements: LPS, 10 or 100 ng/mL IL-1β (IL-1β 10 or IL-1β 100, respectively). A microscopic evaluation of cell viability by LIVE/DEAD assay showed that cells remained viable at day 8 of culture, with no apparent differences between the tested conditions, indicating that neither the low oxygen tension nor the puncture and proinflammatory stimulus leads to a significant loss of cell viability (Fig. 2A). In addition, glucose and lactic acid levels were quantified in the collected supernatants during culture (Fig. 2B, C). Results showed similar glucose consumption and lactic acid production for all stimulated groups and the control, corroborating cell viability results.
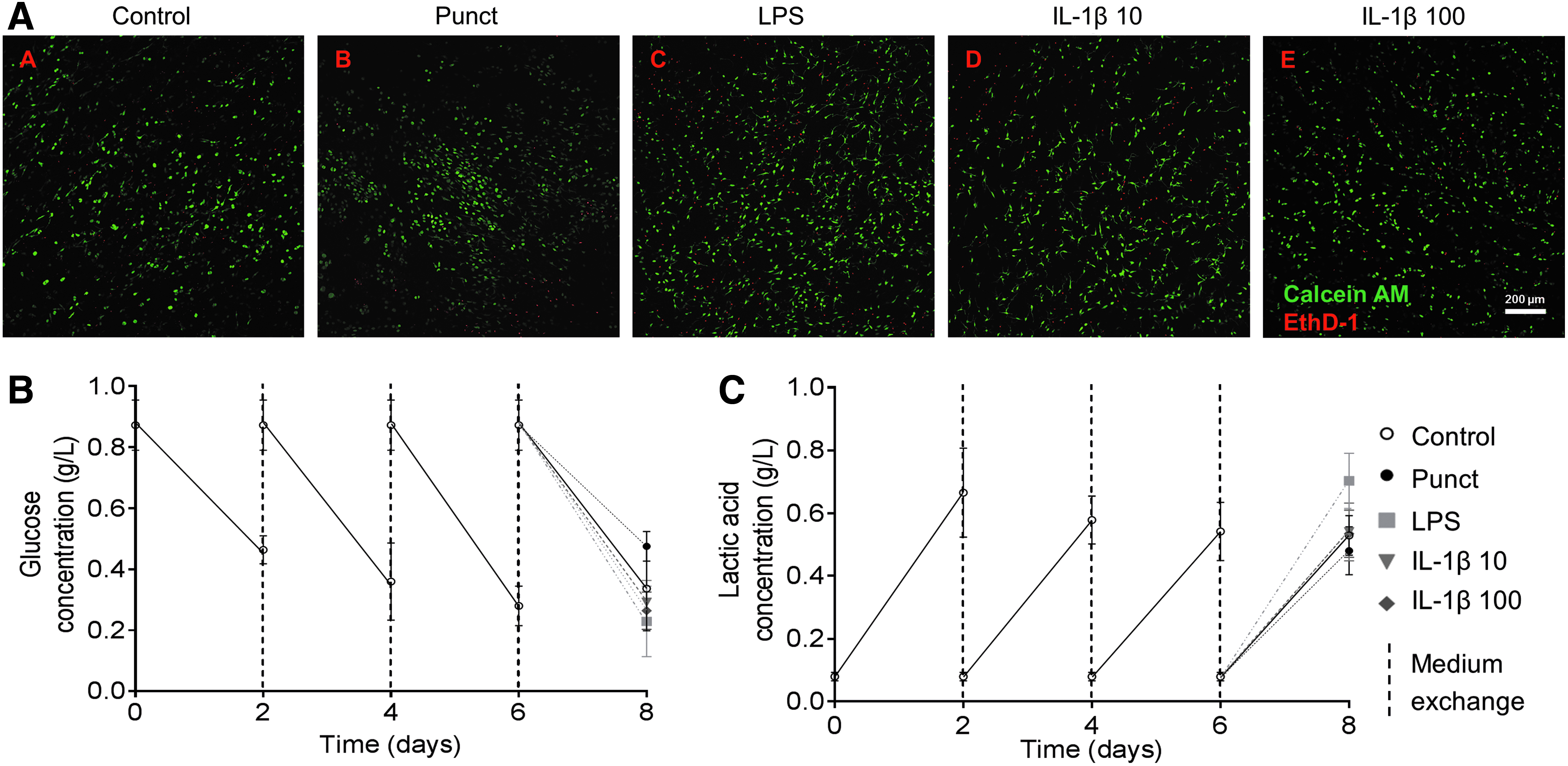
Cell viability and metabolic activity over the culture period of IVD punches cultured under constrained conditions.
Analysis of proinflammatory markers, MMPs, and ECM proteins of the proinflammatory IVD organ culture model
PGE2 production was quantified in culture supernatants over time. No statistically significant differences were found between time points during culture in BM. After 2 days of stimulation with different conditions (day 8), PGE2 production significantly increased for all the groups tested, in comparison to the control (Fig. 3A). LPS group showed the highest PGE2 fold increase (11.5 ± 1.2-fold, p < 0.0001), while Punct presented only 1.8 ± 1.2-fold (p < 0.01). IL-1β 10 and IL-1β 100-stimulated groups achieved, respectively, 3.7 ± 1.2- and 5.8 ± 1.2-fold (both with p < 0.0001). Thus, IL-1β 100 increased 1.6 ± 0.7-fold (p < 0.05) compared with IL-1β 10.
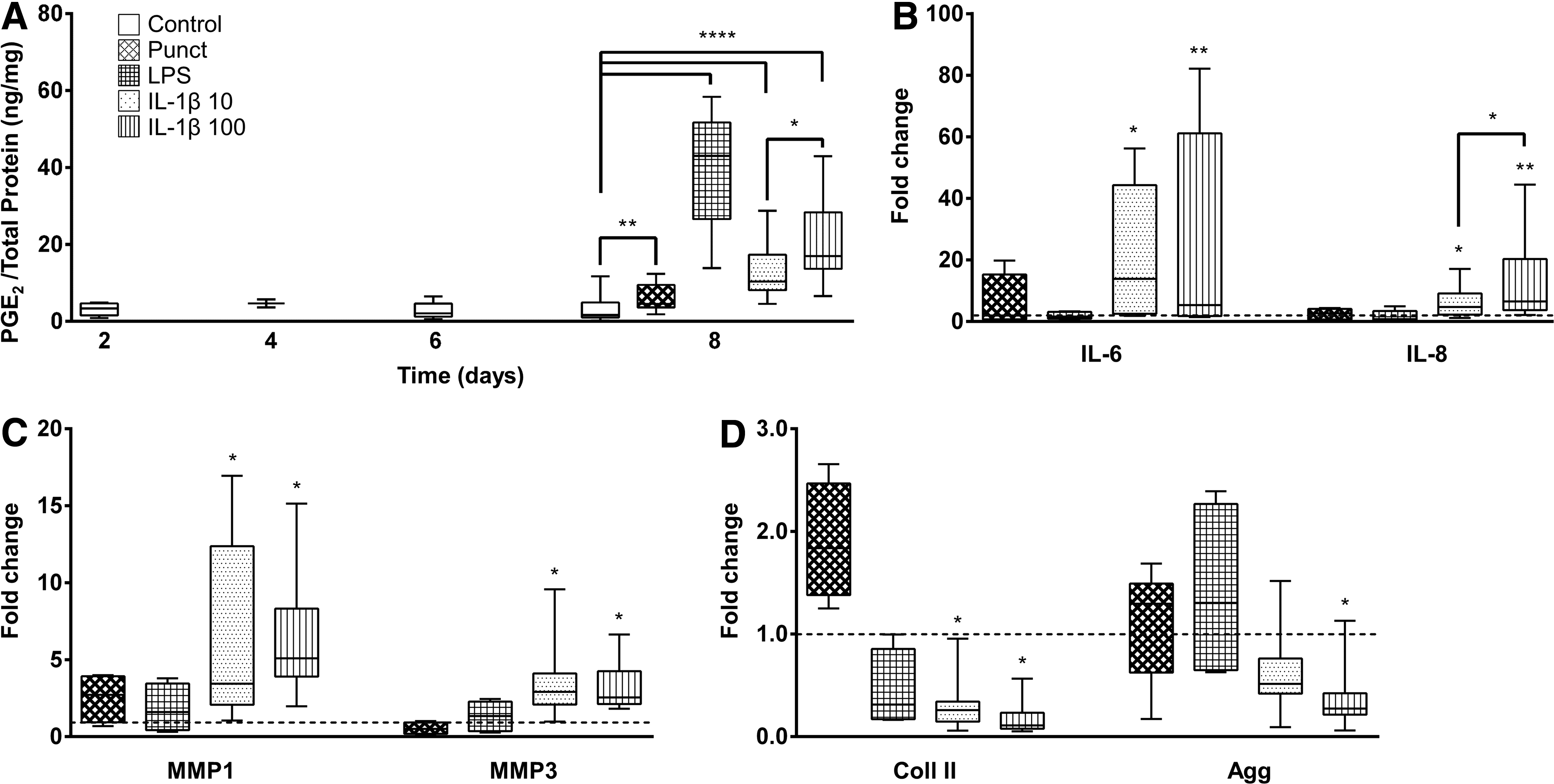
Quantitative analysis of proinflammatory markers, matrix degrading enzymes, and extracellular matrix (ECM) components in IVD organ cultures. The IVD organ cultures were stimulated with Punct, puncture plus culture medium supplementation with 10 μg/mL LPS (LPS), 10 and 100 ng/mL IL-1β (IL-1β 10 and IL-1β 100, respectively), and compared with unstimulated control.
Gene expression of proinflammatory markers and MMPs was also analyzed at day 8 of culture. IL-1β 10 and IL-1β 100 groups showed statistically significant upregulation of IL-6, IL-8, MMP1, and MMP3 expression compared with unstimulated discs (p < 0.05, Fig. 3B, C). For IL-8, IL-1β 100 was also significantly upregulated in comparison to IL-1β 10 (p < 0.05).
Concerning gene expression of ECM proteins, Coll II and Agg was downregulated in the presence of IL-1β (Fig. 3D). While a statistically significant downregulation was observed in Col II expression for IL-1β 10 and 100 groups, Agg expression was only downregulated for IL-1β 100 (p < 0.05).
Since IL-1β stimulation, in addition to puncture, appears to induce a proinflammatory/degenerated IVD environment, with an upregulation of proinflammatory markers, as well as a downregulation of ECM proteins, and moreover, since it is a more physiological method to induce this inflammatory milieu than LPS, the proinflammatory model with IL-1β was selected for the following experiments.
Evaluation of an anti-inflammatory (Df) injection in the proinflammatory IVD organ culture
To validate the proinflammatory IVD organ culture model, an injection of a commonly used anti-inflammatory drug (Df) was tested. For that, soluble Df (19 μM) was injected in the NP, 3 h after proinflammatory stimulus. The concentration and time point of addition were selected based on previous work from our group with Df treatment in human macrophages. 45
Two days after injection, PGE2 production significantly decreased in the groups supplemented by Df (IL-1β 10 + Df and IL-1β 100 + Df, p < 0.05) in comparison to the respective IL-1β-stimulated group alone (IL-1β 10 and IL-1β 100, Fig. 4A). Expression of IL-6 was significantly downregulated, and MMP3 was slightly downregulated with Df injection for both IL-1β concentrations (p < 0.05, Fig. 4B, C). For the higher IL-1β concentration used, IL-8 and MMP1 were significantly downregulated after Df injection (p < 0.05, Fig. 4B, C). Concerning the ECM proteins, Coll II did not show significant differences after Df injection. In contrast, an upregulation of Agg was observed after Df injection for both IL-1β concentrations (p < 0.05, Fig. 4D).
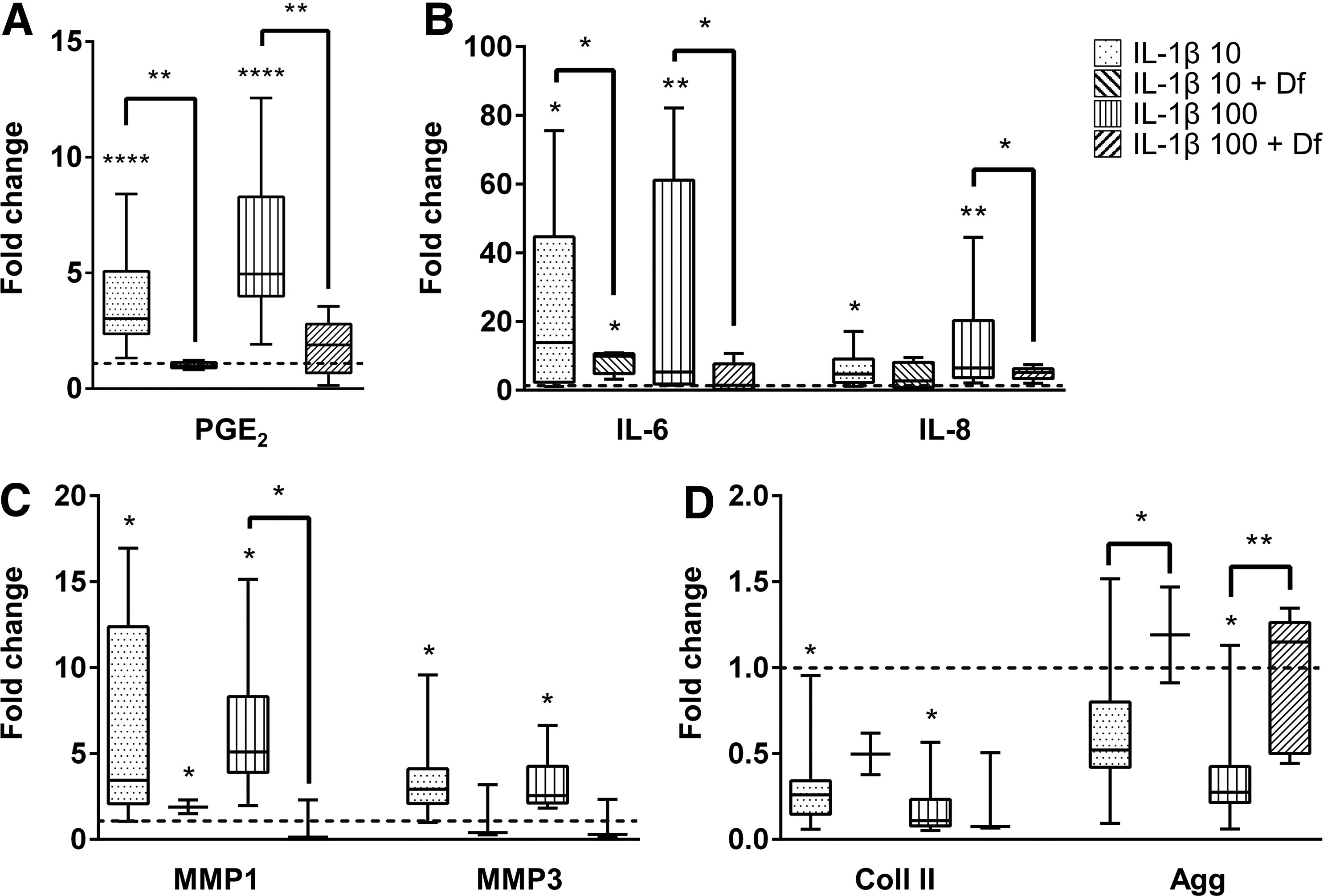
Effect of diclofenac (Df) injection in IVD organ cultures. Quantitative analysis of proinflammatory markers, matrix degrading enzymes, and ECM components in IVD organ cultures after proinflammatory stimulus with puncture plus culture medium supplementation with 10 and 100 ng/mL IL-1β (IL-1β 10 and IL-1β 100, respectively), and treatment with 19 μM Df for 2 days (IL-1β 10 + Df and IL-1β 100 + Df).
Evaluation of MSCs injection in the proinflammatory IVD organ culture
MSCs anti-inflammatory effect was evaluated in the IVD proinflammatory model stimulated with IL-1β 10. For this, MSCs were injected in the IVD using a HSA-HA hydrogel as a vehicle. Injection of either MSCs embedded in HSA-HA (MSCs/HSA-HA) or HSA-HA alone did not alter PGE2 production (Fig. 5A). Regarding the proinflammatory cytokines, there was an apparent slight downregulation of IL-6 and IL-8 expression in the MSCs/HSA-HA injection group compared to IL-1β 10 stimulus and to the injection of HSA-HA alone (Fig. 5B). MMP1 and MMP3 did not appear to have been differently expressed between injections of MSCs/HSA-HA or HSA-HA alone, however, they seem to be upregulated in those two groups in comparison to IL-1β 10 (Fig. 5C). Gene expression of Coll II and Agg was downregulated at all conditions with proinflammatory stimulation compared to the unstimulated controls, and HSA-HA scaffold alone or combined with the MSCs further decreased matrix protein expression (p < 0.05, Fig. 5D).
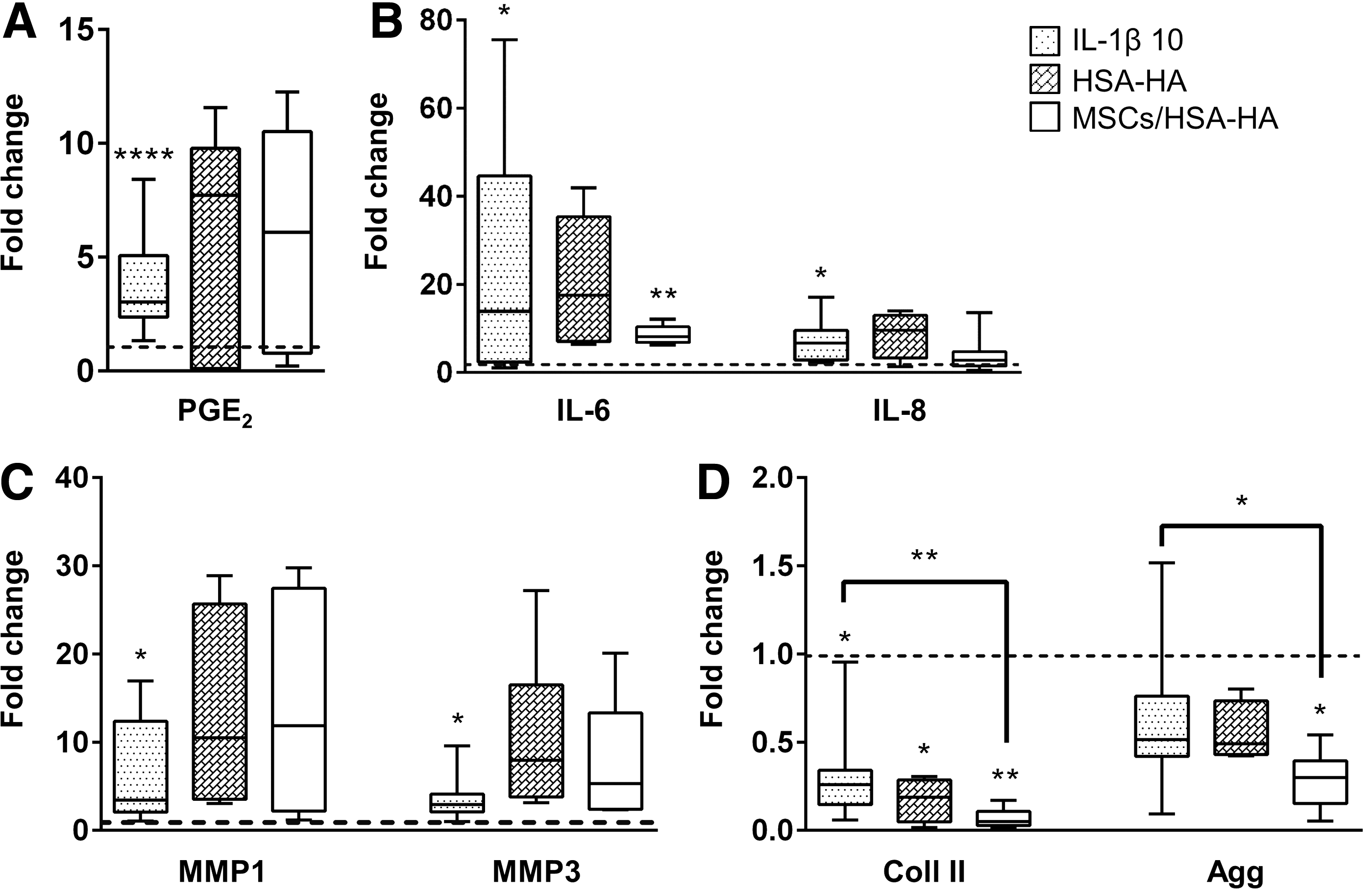
Effect of mesenchymal stem/stromal cells (MSCs) injection in human serum albumin/hyaluronan (HSA-HA) hydrogel in IVD organ cultures. Quantitative analysis of proinflammatory markers, matrix degrading enzymes, and ECM components in IVD organ cultures after proinflammatory stimulus with puncture plus culture medium supplementation with 10 ng/mL IL-1β (IL-1β 10), and treatment with HSA-HA or MSCs/HSA-HA for 2 days.
Discussion
In the present study, we established a standardized bovine organ culture system that simulates proinflammatory conditions. The approach used tissue punches that allowed the standardization of explant size. The physical culture conditions in culture plates constrained by inserts and compressed by a constant static loading prevented tissue swelling and tissue deformation that usually occurs without end plates, as reported in the literature.23,51–53 Moreover, the tissue is more easily collected using a less complex protocol (e.g., it does not require sawing the bone or jet lavage).52,54,55 In our approach, we have used constrained conditions in all experimental groups for standardization of the samples (prevention of swelling), but we did not simulate complex loading protocols, although we are aware of the importance of complex loading on the initiation and progression of disc degeneration.23,56 Nevertheless, GAG loss in these conditions was already reported to naturally occur in culture. 52 Although organ cultures are accepted as more reliable models than 2D in vitro cultures, since they constitute a step further in complexity before in vivo studies, there also some limitations that all should be aware. Namely, an organ culture model cannot simulate complex interactions with adjacent tissues and can only be kept for a limited time in culture14,40,57 to guarantee tissue viability. Another limitation is the lack of vascularization and immune cells. In our approach, tissue viability was assured by controlling metabolite production/consumption levels during culture time and the culture period was rather short. Concerning vascularization, since mature native IVD lacks vasculature, this limitation is of minor relevance in disc organ culture models.
To promote a proinflammatory environment, we tested different approaches. Some commonly methods in the literature use mechanical injury by needle puncture,58,59 to stimulate disc degeneration, or medium supplementation with LPS. 60 However, although LPS is a known stimulus of inflammation,60,61 it is not really a physiological approach. On the contrary, IL-1β is an interesting candidate, since it is expressed at high levels in the pathogenesis of disc disease13,62,63 and it has been demonstrated that exposure of NP cells to IL-1β leads to altered mechanical function, primarily due to loss of GAG. 64
In general, none of the conditions that induce a proinflammatory environment impaired cell viability or altered glucose consumption/lactic acid production, thus, they did not substantially alter cell metabolism in our approach. This is in accordance with other studies that observed a cell survival rate (after 14 days of rabbit IVD organ culture) higher than 90% in proinflammatory conditions (LPS and IL-1β). 60 However, Korecki et al. showed that at the needle insertion site cell damage may occur, 40 while results from Ponnappan et al. revealed an increase in apoptosis after IL-1β treatment. 44
LPS stimulation is known to induce COX-2 expression 65 and consequently generates different prostanoids, including PGE2. 66 PGE2 is excessively produced in response to proinflammatory cytokine signaling, particularly IL-1 and TNF-α.12,67 LPS was also shown to stimulate proinflammatory cytokine production (IL-1β, IL-6, and IL-8) by chondrocytes 68 and by murine disc cells. 69 In the present study, LPS stimulus significantly increased PGE2 production but no obvious effect was observed in other proinflammatory cytokines analyzed or in matrix remodeling.
In contrast, only (100 ng/mL) IL-1β-treated discs showed a significant upregulation of proinflammatory cytokines and MMP expression accompanied by a significant downregulation of ECM proteins, in accordance to disc degeneration description.43,44,63 The other study by Burke et al. reported that disc cells from patients with scoliosis or degenerated IVDs respond to an exogenous proinflammatory stimulus with an increased secretion of proinflammatory markers IL-6, IL-8, and PGE2.41,67 Therefore, a proinflammatory IVD organ culture under loaded conditions, punctured and stimulated with IL-1β (100 ng/mL), was selected as the most reliable model for further studies.
An anti-inflammatory drug (Df) was injected in discs stimulated by IL-1β and its effect on proinflammatory/degenerative IVD was evaluated. As expected, Df injection was able to decrease PGE2 production, since Df is a COX-2 inhibitor. However, besides this known effect, Df also downregulated IL-6, IL-8, and MMP1 expression, while stimulating Agg synthesis, suggesting that this treatment might not only reduce inflammation but could also delay matrix protein degradation and/or increase ECM protein levels. Intradiscal injection of steroids or glucocorticoids to control inflammation in IVD has been used in clinics. However, their influence on deregulation of matrix turnover leading to further disc degeneration is also known. 70 Df direct injection into the IVD may have a limited clinical use as it has a short biologic half-life and may require repeated administrations. Therefore, a sustained delivery system that prolongs Df release in the disc may be a promising alternative. In fact, Df intraperitoneal injection in a rat lumbar disc herniation model showed a reduced analgesic effect with time. 71 Nevertheless, in a pig model of NP-induced nerve root injury, Df reduced NP-induced nerve root dysfunction, showing good prognosis after Df treatment. 72 Nonetheless, most of these studies are focused on pain by analysis of change in disability and pain scores and not biological effects on matrix turnover and inflammation.73–75
In parallel, the effect of MSCs transplantation on the proinflammatory/degenerated IVD organ culture was here investigated. MSCs are capable to differentiate into NP-like cells, 38 increasing expression of nonspecific markers as Agg or Coll II, 30 and were reported to induce less pain in IVD degeneration human clinical trials. 37 The cell density was used based on the work by Serigano et al., which showed that 106 MSCs per transplanted disc were the ideal number of cells, since less viable cells were detected when 105 MSCs were transplanted and more apoptotic cells were found in 107 MSCs transplanted discs. 76
The vehicle used to inject MSCs was based on a HSA-HA hydrogel. This hydrogel was shown to be adequate for MSCs differentiation in chondrogenic lineage, in vitro, 27 and to enhance disc endogenous repair after 6 months, in an in vivo nucleotomized sheep model. 47 In vivo studies using this hydrogel showed good integration with the host without reporting associated inflammatory response.77,78 Only low levels of the proinflammatory cytokine IL-1β were reported in the nucleotomized sheep model, 6 months after implantation. 47 Nevertheless, the results from our model demonstrate that this carrier by itself activates the production of proinflammatory mediators PGE2, IL-6, and IL-8 by disc cells, when compared to control discs. This suggests a foreign-body-type reaction, similar to what happens with biomaterial recognition by immune cells. Interestingly, upon implantation, MSCs were able to reduce this activation status, demonstrating their immunomodulatory effects in this model also.
The literature reports that MSCs are able to increase the expression of the immunesuppressive ligand FasL in IVD 35 as well as downregulate gene expression of proinflammatory cytokines (IL-3, IL-6, IL-11, IL-15, TNF-α) and MMPs produced by rat NP cells. 36 Bertolo et al. also showed that MSCs are able to reduce IgG production by human NP fragments and slightly reduce TNF-α expression, although no influence was observed on IL-1β. 14 Nevertheless, in the present study, MSCs effect on proinflammatory markers appeared to be highly donor dependent. PGE2 revealed either an upregulation or a decrease, depending on the donor, and only a slight downregulation of IL-6 and IL-8 expression was observed after MSCs/HSA-HA injection. The preliminary short-term findings at gene expression level rather suggest impaired matrix formation and increased matrix degradation. Although 2 days after injection appears to be an adequate time point to address the inflammatory markers, we cannot exclude that it may be a too early time point to analyze matrix formation. The other aspect that we cannot exclude is the use of human MSCs on a bovine disc organ culture, although human MSCs are known by their immunomodulatory capacity79,80 and have frequently been used in animal studies from other species, without immune suppression.81,82 In those cases, no exacerbated immune response was observed. New experiments with MSCs are currently in progress to clarify the open question about the immunomodulatory role of MSCs in the degenerated IVD.
In conclusion, we have developed and validated a reproducible proinflammatory/degenerative organ culture model. This model is suitable to investigate inflammation-associated mechanisms and other possible pathways that cause disc degeneration. Moreover, this ex vivo model could be used to assess cytotoxic effects of novel pharmaceutical strategies for IVD before animal experimentation. Particularly, Df intradiscal injection seems to be a promising approach to control inflammation while delaying and/or decreasing matrix protein degradation. Regarding the suitability of MSCs injection to modulate the inflammatory response in the degenerated disc, although this approach could be tested in the model developed, our findings were not uniform among different MSCs donors, suggesting that MSCs-based therapy with regard to controlling the inflammatory response in the IVD requires further investigation.
Footnotes
Acknowledgments
The authors of the present study wish to thank Helga Bach and Iris Baum for their excellent technical assistance and the local abattoir Ulmer Fleisch GmbH for providing bovine tails. This work had the financial support of FCT/MEC through National Funds and, when applicable, was cofinanced by the FEDER through the PT2020 Partnership Agreement under the 4293 Unit I&D. G.Q.T. and C.L.P. Also acknowledge FCT for their PhD grants (SFRH/BD/88429/2012 and SFRH/BD/85779/2012, respectively). The authors also acknowledge the German Spine Foundation (Deutsche Wirbelsäulenstiftung), the German Academic Exchange Service (DAAD), and the Conselho de Reitores das Universidades Portuguesas (CRUP).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.