
Abstract
Background:
The production of iodine-containing thyroid hormones necessary for brain development in the fetus depends not only on maternal dietary intake but also on placental iodine transport. The optimum level of iodine nutrition during pregnancy and the proportion of the pregnant population reaching this level have previously been evaluated. Little information exists on the ability of the placenta to either accumulate or store iodine. This study aims to investigate iodine uptake and tissue iodine content within placental tissue obtained from women delivering at term.
Methods:
Samples (∼1 cm3) obtained from placental cotyledons (n = 19), thyroid (n = 4), and uterine myometrial (n = 4) tissue were incubated for 6 hours with 125I in the presence and absence of potassium perchlorate. To account for variation in tissue composition, results were expressed in cpm 125I/μg DNA.
Results:
Placental uptake of 125I (375 cpm/μg DNA) was significantly higher than that of control myometrial tissue (226 cpm/μg DNA) (p < 0.05) and was ∼25% that of thyroid tissue (1702 cpm/μg DNA). Uptake of 125I could be partially blocked in the thyroid and placenta, respectively, by potassium perchlorate (100 μM), which had no effect on uptake by myometrial tissue. Iodine content of tissue samples measured using an alkaline ashing technique with Sandell-Kolthoff colorimetry gave a mean value for total iodine of 30.4 ng/g placental tissue (range 21–50 ng/g), 1.74 ng/g myometrial tissue, and 1037 ng/g thyroid tissue.
Conclusions:
Placental iodine content was only ∼3% that of the thyroid, but on the basis that neonatal iodine stores are very low and highly sensitive to fluctuations in maternal iodine supply, we postulate that placental iodine bioavailability makes a significant contribution to protection against neonatal hypothyroidism. These findings suggest that the placenta has a role not only in uptake but also in storing iodine as a possible means of protecting the fetus from inadequacies in maternal dietary iodine intake.
Introduction
Can the placenta store iodine?
Is such storage sufficient to compensate for inadequate iodine intake?
In this study, we aimed to investigate iodine uptake and tissue content in placental tissue at term (>37 weeks) gestation, to determine the relative rate of placental iodide uptake and the ability of the placenta to store a supply of iodine.
Materials and Methods
Tissue collection
Placentas (n = 9) were obtained from a random group of euthyroid women undergoing prelabor caesarean section at term. All sections were carried our after 38–40 weeks and were elective with no associated pathology and none were in labor. Ages ranged from 26 to 39, and eight were multiparous and one primiparous. There was one twin pregnancy, a dichorionic twin from which only one placenta was harvested.
Women being treated with anti-thyroid drugs were excluded from the study. Mean/median age of the women was 30.7/31.4 years (range 21.7–38.7) and mean/median parity was 2 (range 1–5).
To establish optimum 125I uptake times, three segments were cut from three placentas (n = 9) and incubated for 7 hours at 37°C. A 4-hour incubation time was subsequently selected for uptake experiments. Similarly, the optimum concentration of potassium perchlorate, a known inhibitor of the NIS, was established as 100 μM. When optimum conditions had been established, nine placentas were used for tissue iodine uptake and studies. Segments ∼1 cm3 were cut from 2 to 3 cotyledons per placenta (n = 19). Due to the complexity of the procedures only three placentas were chosen for 125I efflux studies. To enable comparisons, the same number of thyroid (n = 3) and myometrium (n = 3) served as controls. A total of nine segments (three for each time point) were taken from each placenta with the same number from thyroid and myometrium.
The same placentas were used for studying tissue iodine content. Six cotyledons were selected from different locations within each placenta for tissue iodine content studies (n = 54 in total).
Thyroid tissue for iodide uptake studies (n = 4) and for iodine content (n = 6) (mean/median age = 49.4/51.2) was obtained from euthyroid patients undergoing partial or total thyroidectomy due to nodular goiter, and myometrial samples serving as controls (n = 4) were obtained from premenopausal women undergoing hysterectomy for benign disease (mean/median age = 42.6/41.4). Tissue iodine content was also measured in other extrathyroidal control samples [fat (n = 3) and placental amniotic (n = 2) and chorionic (n = 2) membranes]. Fat samples were obtained from healthy women undergoing reduction mammoplasty. Amniotic and chorionic membranes were obtained from the same subjects as the placental cotyledons.
Tissue iodine uptake studies
Approximately 1 cm3 portions of each tissue type were washed in buffered Hanks balanced salt solution before incubation. Incubation times with 125I were ranged from 1 to 7 hours to establish the time at which optimum iodide uptake occurs. Uptake studies were carried out in the presence or absence of potassium perchlorate (100 μm), a known inhibitor of the NIS. This concentration was selected as that which produced a plateau in blocking 125I uptake (see Fig. 2). After the incubation period, surface adhering radioactive iodide was then removed by rinsing twice with ice-cold phosphate-buffered saline (PBS), and 125I activity in tissue recorded as cpm was counted on a γ counter. To account for variation in tissue composition, results were expressed in cpm/μg DNA with DNA content being measured using Quaigen DNA Extraction kit.
Tissue efflux studies
After preincubation with 125I, samples were counted and then placed in PBS. The tissue was moved to fresh PBS every 5 minutes and the original PBS was counted on a γ counter to determine the rate of efflux from the different tissue types.
Tissue iodine content
Tissue iodine content was determined in 54 cotyledon samples from 9 placentas, 6 thyroid, 4 myometrium, 4 placental membranes, and 3 samples of fat. Samples were washed with 0.9% NaCl solution and weighed. Approximately three volumes of 50 mM KH2PO4 were then added to each sample. Samples were then dried overnight and incinerated for 3 hours at 600°C before colorimetric analysis in the Sandell-Kolthoff reaction. Results were expressed as μg I/g tissue.
Statistical analysis
Statistical analysis was performed using GraphPad Prism software (version 4.0; GraphPad Software Inc., San Diego, CA). Paired Student's t-tests were used to compare differences between the perchlorate blocked and untreated tissue samples. Results are expressed as cpm/μg DNA and p < 0.05 was considered significant. Wilcoxon's Rank sum test was used to compare differences in iodine content between different tissue types.
Results
The rate of 125I uptake in placental tissue samples over a period of 7 hours in the presence and absence of perchlorate (100 μM) is shown in Figure 1. Placental tissue exhibited saturable uptake of radioiodide, which increased steadily for the first 4 hours of incubation to a peak of 6210 cpm in the test sample but only 3940 cpm in the perchlorate blocked sample. Thereafter, further uptake was minimal remaining relatively constant in both test (6286–6234 cpm) and perchlorate blocked samples (4080–4121 cpm). At a maximum uptake of 125I (6234 cpm) perchlorate blockade was 34%.
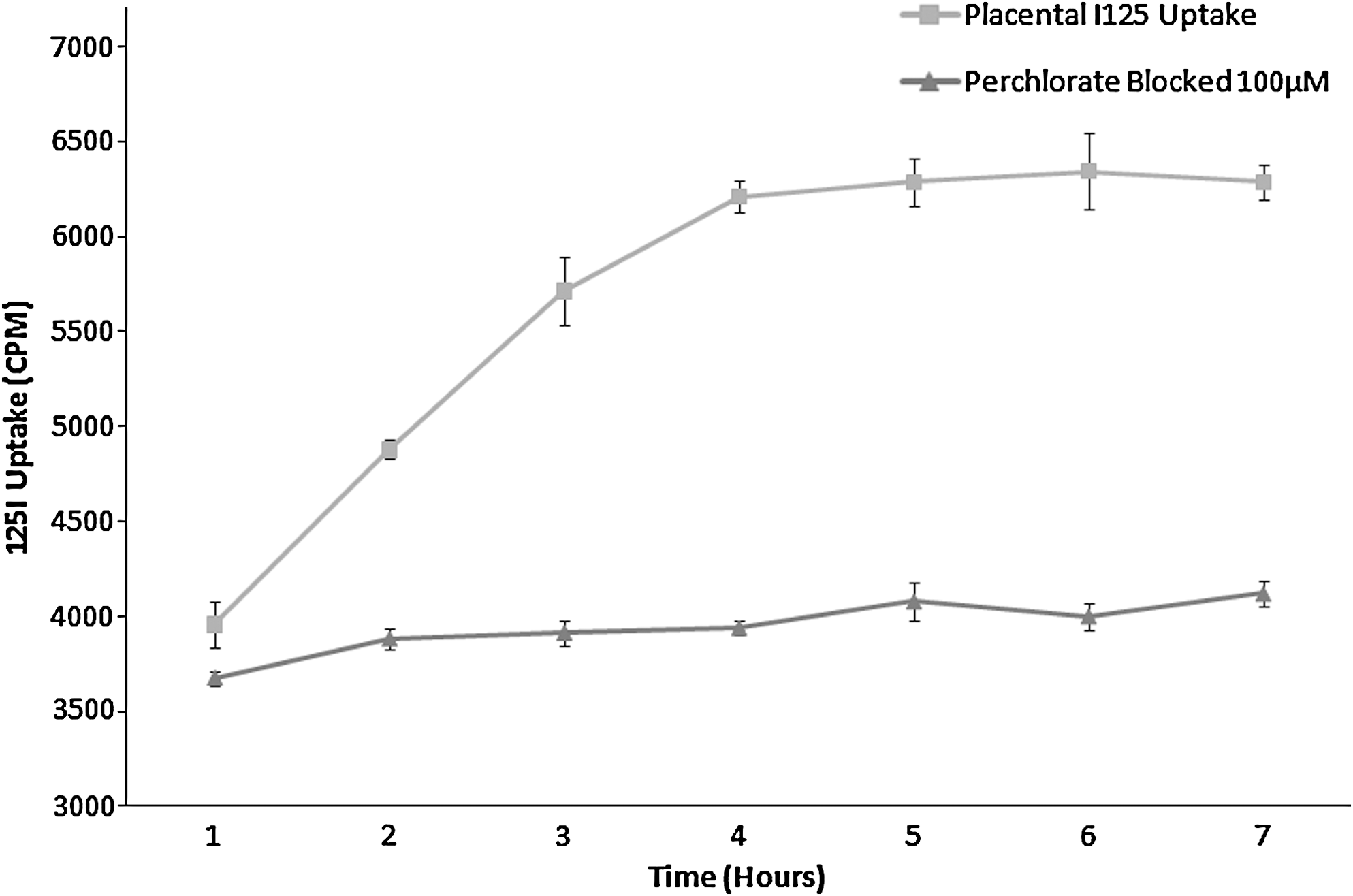
Rate of in vitro placental 125I uptake in placental cotyledons (n = 3) in the presence and absence of potassium perchlorate. Perchlorate (100 μM)-treated tissue showed no increase in uptake over the time course of 7 hours, whereas untreated cotyledons exhibited saturable uptake of iodide reaching a peak after ∼4 hours time course. Error bars represent the SD from the mean. SD, standard deviation.
The concentration-dependent nature of perchlorate blockade of 125I uptake by placental tissue (n = 3) is shown in Figure 2, which correlates iodide uptake with a range of perchlorate concentrations. Uptake in the placenta could be inhibited by the presence of perchlorate in a dose-dependent manner with uptake inhibited 4%, 13%, 24%, and 31% by 1, 10, 50, and 100 μm K2ClO4, respectively.
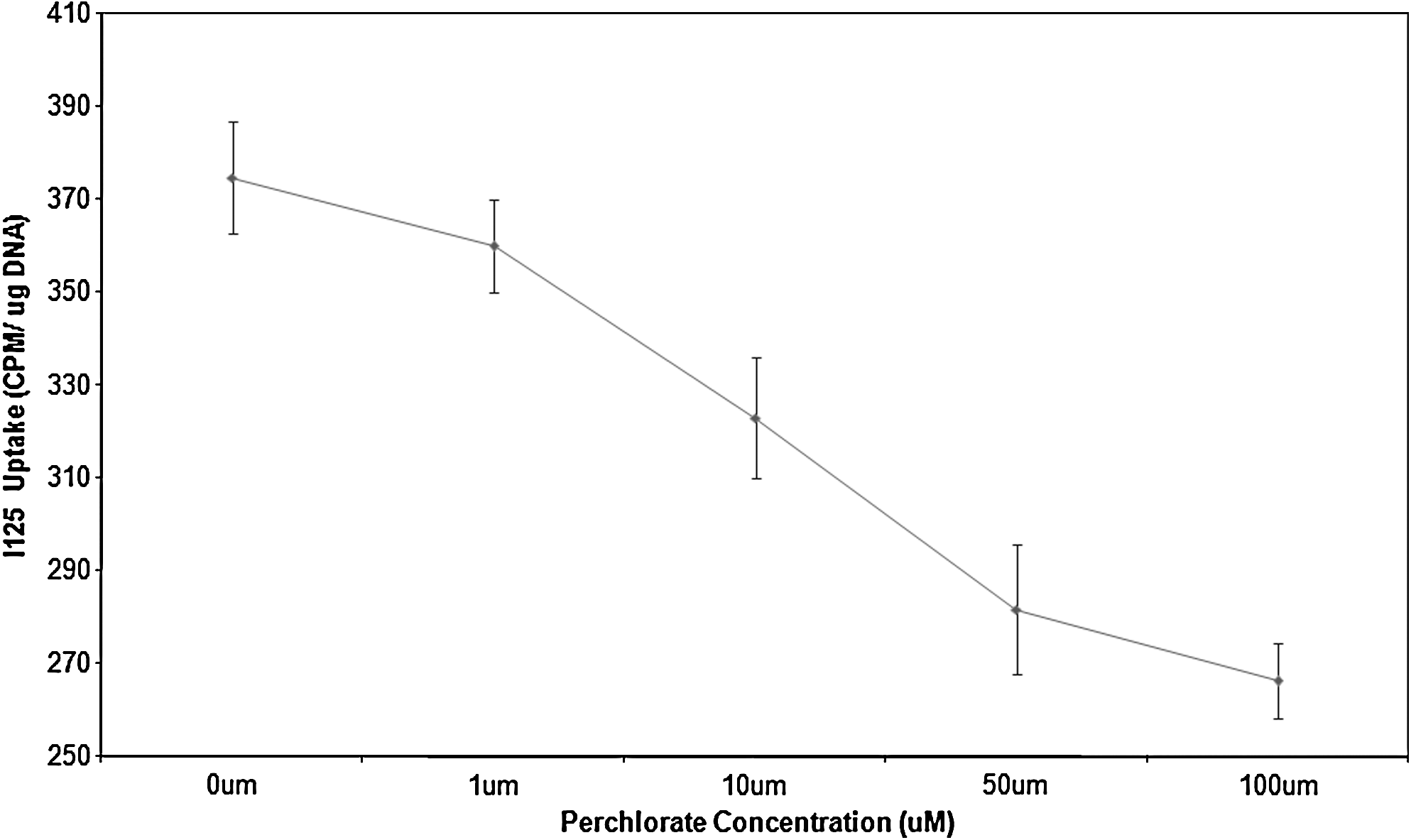
Inhibition of iodide uptake within placentas (n = 3) by varying concentrations of potassium perchlorate. Uptake is measured as cpm/μg DNA on the y-axis, whereas the range of perchlorate concentrations from 0 to 100 μM is shown on the x-axis. Data are displayed as average uptake, with error bars representing the range of values obtained.
Figure 3 shows the median 125I uptake for placental, thyroidal, and myometrial. Tissues were incubated with 125I for a total of 4 hours. The test placental samples exhibited a higher mean uptake than myometrium but a much lower uptake than that of thyroid (∼20% of thyroid uptake). Mean placental uptake was 375 cpm/μg DNA in the placental samples of which 30% of this uptake could be blocked by 100 μM perchlorate (a similar degree of blockade to that of thyroid) (p < 0.005). Thyroid tissue exhibited the highest rate of uptake of the different tissue types studied. The test thyroid samples had a mean uptake of 1702 cpm/μg DNA of which 38% could be blocked in the perchlorate-treated sample. This difference in uptake between test and perchlorate-treated samples was highly significant (p < 0.005). Mean myometrial tissue uptake was 226 cpm/μg DNA in the test samples and could not be blocked with perchlorate, the mean for the perchlorate-treated samples being identical (231 cpm/μg DNA).
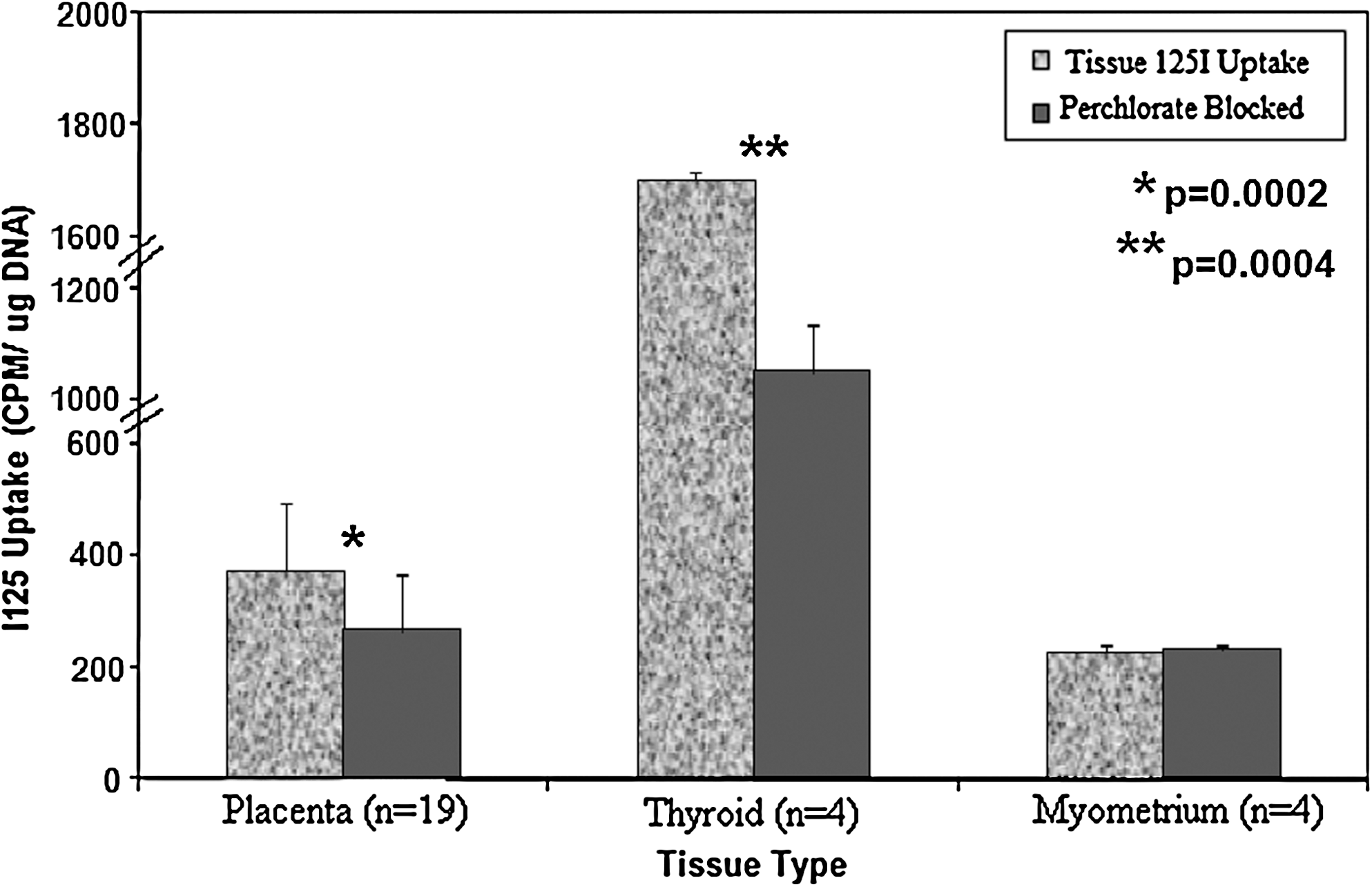
125I uptake in placenta, thyroid, and control myometrium in the presence and absence of potassium perchlorate. Different tissue types in the presence and absence of perchlorate (100 μM) are shown on the x-axis, with the uptake in cpm 125I/μg DNA on the y-axis. Data are displayed as mean uptake, with error bars representing the SD from the mean. There were significant differences in uptake between perchlorate-treated and untreated tissue in both thyroid (p = 0.004) and placental (p = 0.002) tissue.
Figure 4 illustrates the rate of iodide efflux from placental, thyroidal, and myometrial tissues. Tissues were preincubated with 125I before commencement of efflux studies, which lasted for 1 hour. Initially, all tissues had a fast rate of efflux with loss of ∼20% activity in the first 5 minutes. For the next 15 minutes, the rate of efflux from placenta and thyroid tissue was faster than that of myometrial tissue. Thyroid tissue initially showed slightly faster efflux than placenta, but after 25 minutes their efflux values were virtually superimposeable. Thereafter, the rate of efflux from all three tissue types remained relatively constant. After 1 hour of efflux, only 6%, 4%, and 13% of activity remained within placenta, thyroid, and myometrium, respectively. The t 1/2 for thyroid was fastest at 12.13 minutes; placenta had a t 1/2 of 14.28 minutes, whereas the t 1/2 for uterus was considerably slower at 21.80 minutes. Although overall efflux rates were not significantly different between the three tissues over the 60 minutes test period, placenta and thyroid showed significantly greater loss than myometrium over the first 30 minutes (p < 0.01).
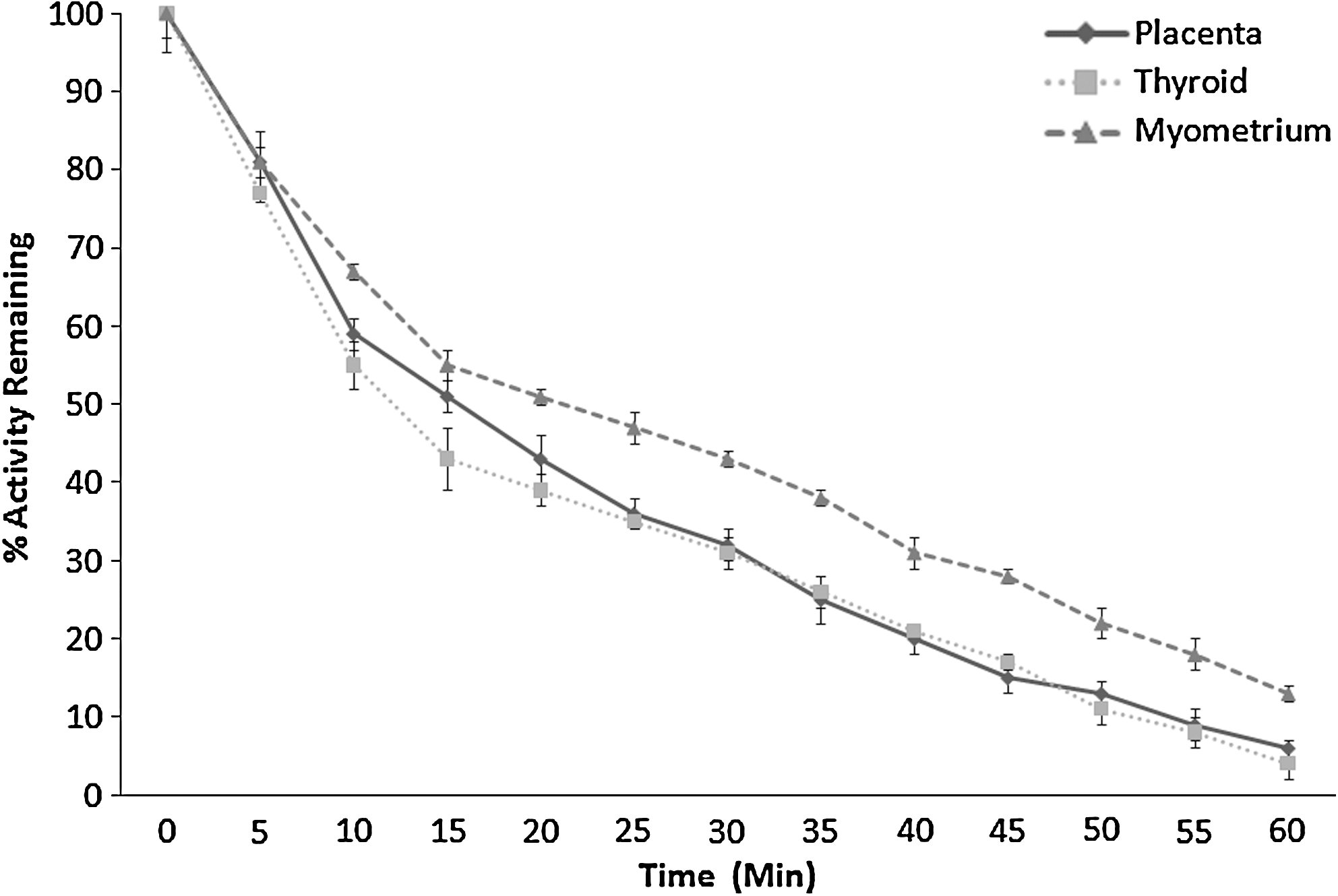
Efflux of iodide from placental (n = 3), thyroid (n = 3), and myometrial (n = 3) tissue over a period of 60 minutes. Activity in percentage of activity remaining is shown on the y-axis, with time measured in 5-minute intervals shown on the x-axis. Each time point was expressed as percentage activity remaining as compared to the initial activity present in each sample. Results are shown as mean ± SD. Error bars represent the SD.
The scattergraph in Figure 5 shows the variation in iodine content between the different tissue types but also between different placentas. The iodine content of the control tissue was the lowest of all tissue types, with median tissue iodine content of only 1.58 ng I/g tissue (range 0.38–4.12 ng I/g tissue). Thyroid tissue had the highest median tissue iodine content of 1031 ng I/g tissue (range 750–1299 ng I/g tissue), whereas placental iodine content was intermediate, but median placental iodine content was 27.0 ng I/g tissue (range 21–50 ngI/g tissue) and varied very little within an individual cotyledon (CV = 9.1%). However, the variation in iodine content between different cotyledons from the same placenta was greater with inter-cotyledon CV ranging from 4.5% (range 30.4–34.4 ng I/g tissue) to as much as 87% (range 0–81.0 ng I/g tissue). These data are shown in tabular form in Table 1. Whereas median placental cotyledon iodine content was only ∼3% that of the thyroid, it was significantly higher than the other tissue types examined (p < 0.005; Wilcoxon's Rank sum test).
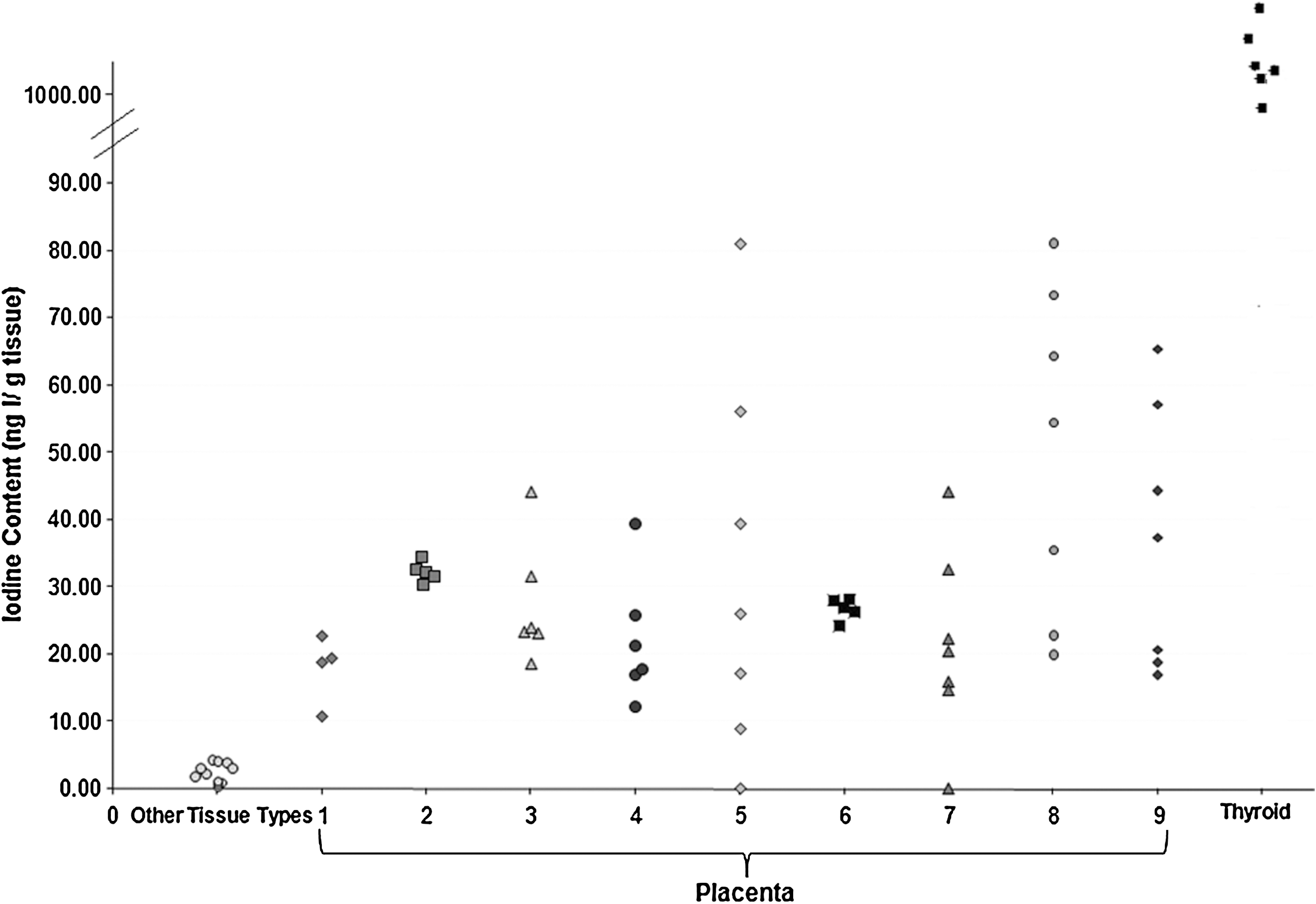
Tissue iodine content of placental, thyroid, and other control tissue types. (Other tissue types are uterine myometrium, fat, placental amnion, and chorion.) Numbers represent each different placenta, whereas each point represents either a single tissue or a cotyledon within a placenta.
Other tissue types are uterus, fat, and placental amniotic and chorionic membranes.
Discussion
Iodine supply to the fetus depends not only on maternal iodine intake but also on the ability of this iodine to cross the placenta and become available for incorporation into fetal thyroid hormones (4). Not only do these results strongly suggest a role for the placenta in uptake of iodine, but they also indicate the hitherto unreported possibility that the placenta may act as a storage site supplying iodine to the fetus.
Although placental iodine uptake was significantly less than that of the thyroid (by a factor of 4.5), the mechanism of this uptake appears similar to that in thyroid (21) in terms of both rate of uptake and known inhibitors of uptake. Our findings demonstrate that the placenta exhibits saturable uptake of iodine, a characteristic shared with the thyroid as shown in previous reports (21). The iodide transporter NIS has been demonstrated in placental tissue (8 –10), and our work supports these earlier findings, uptake of iodide being partially blocked by potassium perchlorate, a known inhibitor of NIS. Indeed, the level of blockade of iodide uptake in placenta was similar to that of thyroid (30% and 38%, respectively). Iodide efflux patterns also showed similarities between the two tissue types, with the control myometrial tissue showing a slower rate of efflux. Overall, whereas thyroid has a demonstrably greater capacity to take up iodide, the placenta and thyroid do appear to share mechanisms of uptake and efflux.
One of the main strengths of this study is that it was performed on fresh tissue from healthy term placentas sampled before labor onset to most closely represent normal physiological conditions in late pregnancy. The findings, nevertheless, do pose a number of further questions. All placentas used were from term pregnancies, and it remains to be determined if placentas at earlier gestations would have a similar iodine content. The variability in inter-cotyledon iodine content and between cotyledons within one placenta may represent differing proportions of trophoblastic and connective tissues at samples sites. We assume that where the predominant tissue samples was nonvillous, the iodine content would be lower. An alternative explanation might be differences in tissue NIS content, and although the distribution of NIS has been shown to vary within syncytioblasts [Manley et al. (13)], there is not, as far as the authors are aware, any data on NIS distribution within cotyledons. While it might be expected that more active cotyledons within the placenta might contain more iodine, no definite histological locations for increased iodine content could be identified within our results.
Our findings, therefore, suggest a possible homeostatic placental role in compensating for deficient maternal dietary iodine intake through maintenance of fetal iodine supply in women with inadequate dietary iodine intake. The thyroid has a well-established ability to store iodine, and our results show that the placenta also appears to share this capability, though with a much smaller cellular storage capacity than that of thyroid tissue, although balanced by a much greater tissue volume. Nonetheless, considering the relative size of the placenta (our findings demonstrate that an average placenta has the capacity to store between 12 and 50 μg iodine), such storage may play a significant role in supplying iodine to the fetal thyroid. Rapid turnover of iodine from a low volume fetal thyroid may result in the entire iodine content of the fetal thyroid having to be renewed daily (4). Considering that neonatal iodine stores are very low and highly sensitive to fluctuations in maternal iodine supply, the bioavailability of this placental iodine store could, nonetheless, make a significant contribution to protection against fetal and neonatal hypothyroidism (22). The authors are aware that the findings of 125I uptake and higher iodine content do not confirm the presence of a hitherto undescribed iodine storage organ. They do, however, offer the possibility of placental iodine content acting as a compensatory source of iodine to the fetus in the event of inadequate maternal iodine intake. The form in which iodine may be stored or if stored, the mechanism through which it might be made available to the fetus, remains to be elucidated.
Footnotes
Acknowledgment
The authors acknowledge the Health Research Board (Ireland) grant, which supported the research and writing of the article.
Disclosure Statement
The authors declare that they have no conflicting interests and that no competing financial interests exist.