
Abstract
Background:
Thyroid hormone is critical for normal brain development and acts in a spatial and temporal specific pattern. Thyroid hormone excess, or deficiency, can lead to irreversible impairment of brain and sensory development. Chicken ovalbumin upstream-transcription factor 1 (COUP-TF1), expressed early in neuronal development, is essential to achieve normal brain structure. Thyroid hormone stimulation of gene expression is inversely correlated with the level of COUP-TF1 expression.
Methods:
An in vitro method of differentiating mouse embryonic stem (mES) cells into cortical neurons was utilized to study the influence of COUP-TF1 on thyroid hormone signaling in brain development. mES cells were cultured and differentiated in specific conditioned media, and a high percentage of nestin-positive progenitor neurons in the first stage, and cortical neurons in the second stage, was obtained with characteristic neuronal firing.
Results:
The number of nestin-positive progenitors, as determined by fluorescence-activated cell sorting analysis, was significantly greater with triiodothyronine (T3) treatment compared to control (p < 0.05). T3 enhanced the expression of cortical neuron marker (Tbr1 and Rc3) mRNAs. After COUP-TF1 knockdown, the number of nestin-positive progenitors was reduced compared to control (p < 0.05), but the number increased with T3 treatment. The mRNA of cortical neuronal gene markers was measured after COUP-TF1 knockdown. In the presence of T3, the peak expression of neuron markers Emx1, Tbr1, Camkiv, and Rc3 mRNA was earlier, at day 18 of differentiation, compared to control cells, at day 22. Furthermore, after COUP-TF1 knockdown, T3 induction of Rc3 and Tbr1 mRNA was significantly enhanced compared to cells expressing COUP-TF1.
Conclusion:
These results indicate that COUP-TF1 plays an important role in modulating the timing and magnitude of T3-stimulated gene expression required for normal corticogenesis.
Introduction
C
Thyroid hormone plays an essential role in prenatal and neonatal neurological development in mammals (7 –11), influencing neuronal growth and differentiation and the development of neuroglial cells (12 –14). Thyroid hormone modulates the transcription of specific genes so that they are expressed at a developmentally appropriate time and in specific cell types. T3-responsive genes in the cerebellum, including calbindin, inositol 1,4,5-triphosphate receptor, Purkinje cell protein-2 (PCP-2), and myelin basic protein (MBP), are responsive to thyroid hormone stimulation during a specific window in the second and third weeks of postnatal life in the mouse (15). The postnatal increase in T3 sensitivity in the cerebellum correlates with reduced expression of COUP-TF1 (7).
Several mechanisms have been identified for COUP-TF1 transcriptional inhibition of T3 and RA signaling. These include direct competition with thyroid hormone receptor (THR), retinoic acid receptor, or other steroid receptors binding to the DNA response element; heterodimerization with RXRs, the essential nuclear receptor partner; and enhancing the silencing activity of nuclear receptor corepressors (2,16,17). The PCP-2 and Camkiv genes are stimulated by T3 and inhibited by COUP-TF1. In both gene promoters, there is a tandem arrangement of sites that bind COUP-TF1 and THR (18,19). These studies indicate that COUP-TF1 modulates T3 regulation of gene expression in the developing cerebellum, likely by binding near the thyroid hormone response element (THRE) and inhibiting THR binding. In general, when expression of COUP-TF1 is reduced, thyroid hormone stimulation of these genes is enhanced.
Due to the complexity of the cerebral cortex and the cell type–specific regulation of thyroid hormone, it has been difficult to identify a model appropriate to study COUP-TF1 modulation of T3-responsive genes in neuronal development (20). In this study, an in vitro model of neuronal differentiation was modified, and it was applied to mouse embryonic stem (mES) cells (21). This approach allowed the differentiation of pyramidal neurons of cortical occipital cortex (areas that highly express COUP-TF1) in order to study the role of COUP-TF1 in modulating thyroid hormone action. The goal was to determine whether COUP-TF1 modulates the timing and magnitude of expression of T3-responsive genes and is required for modulating thyroid sensitivity in pyramidal neuron differentiation. This model was applied to determine the role of COUP-TF1 in modulating the timing of T3-responsive gene expression required for normal corticogenesis.
Materials and Methods
ES cell culture
Irradiated mouse embryonic fibroblasts (catalog # S1520-100; Invitrogen, Carlsbad, CA) were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (vol/vol). ES-E14TG2a mES cells (catalog # CRL-1821T; ATCC, Manassas, VA) were seeded onto feeder fibroblasts after they were cultured for two days. Mouse ES cells were cultured in Knockout™ DMEM supplemented with 20% Knockout™ serum replacement (vol/vol), LIF (1000 IU/ml), non-essential amino acids (0.1 mM), glutamine (2 mM), sodium pyruvate (1 mM), penicillin and streptomycin (50 IU/ml of each), and 2-mercaptoethanol (0.1 mM) in a humidified incubator with an atmosphere of 5% CO2 at 37°C. The third passage of mES cells were utilized for experiments.
Cortical occipital pyramidal neuronal differentiation
A culture method that promotes mouse embryonic stem cells to differentiate into cortical pyramidal neurons was adapted (21). Cortical pyramidal neuronal differentiation of mES cells occurs in two stages, with specific conditioned medium used for each stage. In stage 1 of differentiation, mES cells were plated at low density (5000 cells/cm2) on gelatin-coated dishes and cultured in DMEM/F12/N2 medium, without any morphogen or serum. Cyclopamine (1 μM) was added from day 2 to day 10 of differentiation. T3 (1 nM) was added, beginning on day 2. In stage 2 of cortical pyramidal neuronal differentiation, day 12, neuronal progenitor clusters were trypsinized and re-plated on polylysine/laminin/gelatin-coated dishes and cultured in DMEM/F12/N2 (laboratory-made B27*) medium. The commercial formulation of neuron primary culture serum-free supplement, B27® (Invitrogen), contains RA and T3. Therefore, a similar neuron growth supplement was made (7), based on a published formulation of B27*, but without RA or T3 (22).
COUP-TFI shRNA lentiviral particles transduction mouse embryonic stem cells and establishing COUP-TF1 knockdown cell lines
COUP-TFI shRNA knockdown was performed according to the shRNA lentiviral particle transduction protocol. Briefly, feeder cells and mES cells were grown in 48-well plates (density 1.0 × 104/cm2 for feeders and 3.0 × 104/cm2 for mES cells), with mES cells kept at 60% confluence. The medium was removed from each well and replaced with a polybrene/media mixture. Then, 100 μL (multiplicity of infection [MOI] 12.5) of shRNA-COUP-TFI lentiviral particles (sc-38821-V; Santa Cruz Biotechnology, Inc., Dallas, TX) and control shRNA-control lentiviral particles (sc-108080) was added to each well. The plate was swirled gently to mix, and then it was incubated for 11 hours. The culture medium was removed and replaced with mES cell medium. Two days later, the mES cells were trypsinized and split into 6 cm dishes with puromycin-resistant feeder cells, and puromycin (1.8 μg/mL), to select successfully transduced mES cells. The puromycin selection was continued until only resistant cells remained. The fraction of mES cells that remained after selection comprised 5–10% of shRNA-COUP-TFI transduced and 10% of shRNA-control transduced mES cells. Single clones were picked, expanded, and then selected again with puromycin.
Fluorescence-activated cell sorting
On day 8 of differentiation, cells were trypsinized and washed with pH 7.4 phosphate-buffered saline (PBS)-Tween and centrifuged at 1000 rpm (113 g) for 3 min and the supernatant was discarded. The pellet was re-suspended and the cell number counted. Cells (2 × 106) were re-suspended in 2% paraformaldehyde and stored at 4°C. To prepare the cells for flow cytometry, ice-cold acetone was added for 10 min at −20°C for permeabilization. Then, the cells were washed with PBS (containing 0.1% triton). Conjugated antibody, 10 μL in 0.5 mL of antibody solution (1% bovine serum albumin [BSA]/0.5%Triton/PBS), was added and incubated for 30 min at room temperature in the dark. The flow cytometer (BD FACS Canto II; BD Biosciences, Franklin Lakes, NJ) was used for analysis. The conjugated antibodies were a mouse monoclonal anti-nestin-phycoerythrin (catalog # IC2736P 1:50; R&D Systems, Inc., Minneapolis, MN) and a mouse monoclonal anti-neuron-specific β-3 tubulin-APC (catalog # IC1195A 1:50; R&D Systems, Inc.).
Immunocytochemistry
Monolayer mES cells were fixed with ice-cold acetone and incubated at −20°C for 10 min. The slides were rinsed three times in PBS-Tween and incubated in 1% Triton (in PBS) at room temperature for 30 min. Samples were blocked in 5% normal goat serum (1% BSA/0.2%Triton-X100/PBS) for 1 h at room temperature. The primary antibody was diluted in 1% BSA/0.05%Triton-X100/PBS and 300 μL added per chamber slide and incubated overnight at 4°C. The second antibody was diluted in 1% BSA/0.05%Triton-X100/PBS and 300 μL per chamber slides of the solution was added and incubated for 1 h at room temperature in the dark. The slide was rinsed three times in PBS-Tween for 3 min each in the dark. The coverslip was prepared with anti-fade mounting medium, and the slide was sealed with nail polish. Primary antibodies were: rabbit polyclonal to nestin (catalog # ab24692 1:200; Abcam, San Francisco, CA); rabbit polyclonal VGLUT1 (catalog # 135303 1:300; Synaptic Systems GmbH, Göttingen, Germany); mouse monoclonal to neuron-specific class 3 β-tubulin (TuJ1, catalog # MMS-435P 1:2000; Covance, Inc., San Diego, CA); and rabbit polyclonal GFAP (catalog # ab7260 1:250; Abcam). Secondary antibodies were: goat anti-rabbit immunoglobulin G (IgG) H&L (FITC; catalog # ab97050 1:250; Abcam); and goat anti-mouse IgG H&L (Alexa Fluor® 647; catalog # ab150115 1:250; Abcam).
Quantitation of VGLUT1-positive cells
VGLUT1-positive cells were stained with an anti-VGLUT1 antibody and imaged by laser scanning confocal microscope (Zeiss LSM720). The VGLUT1-positive cells were identified by green fluorescence. β-tubulin 3-positive cells were identified by red fluorescence and represent the total number of cells differentiated into neurons. The number of VGLUT1-positive and β-tubulin 3-positive cells were counted in three consecutive frames at high magnification and averaged. The number of VGLUT1-positive cells is expressed as the mean percentage of the number of β-tubulin 3-positive cells, with standard error.
Neuronal recordings
The extracellular recordings of cultured neurons were performed using a MED64 System (Alpha MED Scientific, Osaka, Japan). Signals from all 64 electrodes were sampled simultaneously at 20 kHz and stored in a computer (MED64 Mobius software; Alpha MED Scientific). The culture temperature was maintained at 37°C during the measurements. Spontaneous firings and extracted spikes of the cortical neurons were analyzed by software (Mobius Pro package, MED64 Mobius software; Alpha MED Scientific). Spike detection was carried out with a threshold of 5.5 times the standard deviation of the noise level.
Quantitative real-time polymerase chain reaction analysis of gene expression
RNA preparation was performed using the RNeasy RNA preparation minikit (Qiagen Inc., Valencia, CA). Total RNA (2 μg) was reverse transcribed using SuperScript® III First-Strand Synthesis System (Invitrogen). Quantitative real-time polymerase chain reaction (PCR) was performed, as previously described (23). In brief, cDNA was diluted at a ratio of 1:5, 1:25, and 1:125 to generate a standard curve for each pair of primers. For GAPDH mRNA, cDNA was diluted at a ratio of 1:10, 1:100, and 1:1000 and used for the standard curve. The data were normalized to the GAPDH mRNA level and expressed as arbitrary units. The quantitative real-time PCR was performed on an Opticon DNA Engine (M.J. Research, Incline Village, NV). The primer sequences used for real-time PCR are provided in Supplementary Table S1 (Supplementary Data are available at
Cell viability and proliferation assay
The effects of COUP-TF1 on cell viability were assessed by the XTT Cell Proliferation Assay Kit (ATCC), according to the manufacturer's instructions. Briefly, on day 8 of differentiation, control mES cells and COUP-TF1 knockdown mES cells (1 × 105/well) were trypsinized and re-plated in flat-bottom 96-well plates in DMEM/F12/N2 medium at 37°C in 5% CO2 for 24 h. The medium was removed and replaced with fresh medium (150 μL) containing XTT, and the cells were then cultured at 37°C for 6 h. Absorbance was determined on a plate reader at 490 nm (Multi-Mode Plate Reader; Promega, Madison, WI).
Statistical analysis
All data were analyzed using SPSS Statistics for Windows v17.0 (SPSS, Inc., Chicago, IL). Values are expressed as mean ± standard error of the mean (SEM). A two-tailed Student's t-test was performed to compare means. Comparison of the proportions was performed with the use of the chi-square test. Differences were considered significant at p < 0.05.
Results
Differentiation of mES cells to pyramidal neurons
It was previously demonstrated that T3 and RA play distinct roles in differentiation of mES cells into neurons (24). Thyroid hormone induces neuronal differentiation and reduces RA-induced apoptosis. These in vitro culture systems, however, rely on RA as a differentiation morphogen, which may secondarily influence T3-induced gene regulation (25). Therefore, a stem-cell differentiation model for corticogenesis was modified to study T3 action in neuronal differentiation (21). Mouse ES cell neuronal differentiation is divided into two stages and utilizes two different conditioned media (Fig. 1). In stage 1, mES cells are differentiated in the presence of medium containing cyclopamine, which inhibits the hedgehog signaling pathway (Fig. 1). Toward the end of the first step of differentiation, day 8, the majority of the cell population consists of neuronal progenitors, as shown by uniform nestin staining (light green; Fig. 1C). In stage 2, differentiation of the neuronal progenitors continues in conditioned medium 2, supplemented with growth factors, but without RA or T3, for an additional 10–12 days (Fig. 1D and E). Cell morphology and neuronal differentiation markers were analyzed with immunocytochemistry. On day 12, staining for both nestin (green) and β-tubulin 3, an early neuron marker, was detectable (Fig. 1D). On day 22, pyramidal neuron morphology could be seen (Fig. 1E). Neurogenesis precedes gliogenesis during brain development in vertebrates. Glial precursor cells grow under the neurons and guide their outgrowth and connections. From day 22 to day 30, the number of GFAP-positive astrocytes increased (Fig. 1F). High-power magnification of neurons at day 22 shows strong staining for VGLUT1, a pyramidal neuron-specific marker (green), as well as β-tubulin 3 (Fig. 1G–I). Utilizing this culture method, a high percentage of nestin-positive progenitors in the first phase (about 50%) and cortical neurons in the second phase of differentiation (about 60%) was obtained.

Cortical pyramidal neuron differentiation utilizing a mouse embryonic stem (mES) cell model. In stage 1, mES cells were plated on gelatin coated dishes and treated from day 2 to day 10 with medium (no morphogen or serum derivative added), and cyclopamine to inhibit sonic hedgehog, which leads to the generation of nestin-positive (nestin+) neuronal progenitors (shown in schematic). In experiments studying triiodothyronine (T3) effects, T3 (1 nM) was added, beginning on day 2. In stage 2, cells were trypsinized and re-plated on dishes coated with polylysine/laminin/gelatin, leading to the generation of β-tubulin-3+ neurons, followed by the development of glial cells as shown by the expression of Glial Fibrillar Acidic Protein (GFAP). (
Electrical activity of stem cell–derived neurons
The neuronal identity of the stem cell–derived neurons was validated by morphology, mRNA, and neuron-specific protein expression. Additionally, a specialized culture system was utilized to record individual electrophysiology profiles of neuronal firing (Fig. 2). This electrical activity has previously been shown to be a property of cultured pyramidal neurons derived from pluripotent stem cells (26), and provides a strong functional measure of the neurons in this system.
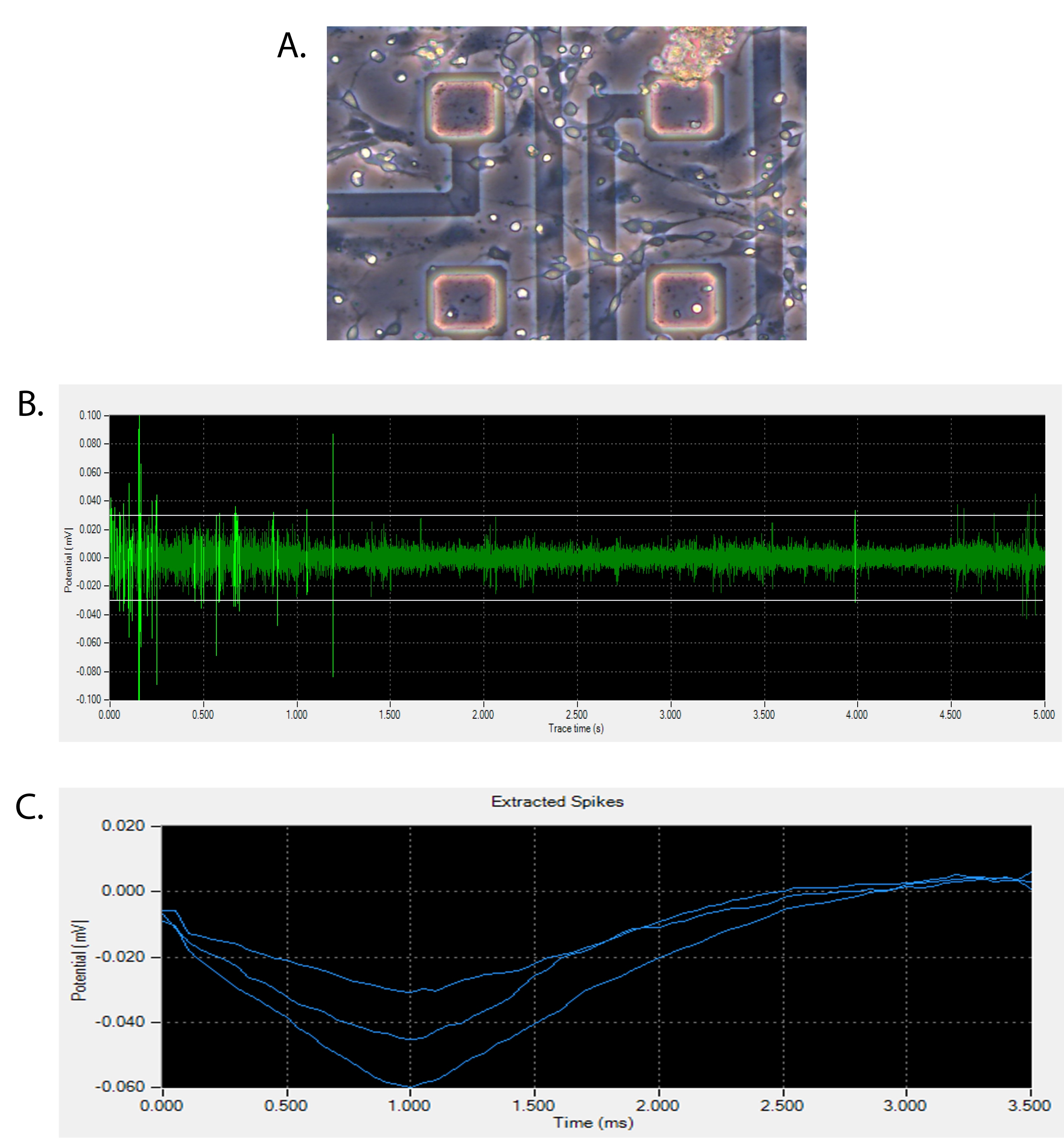
Electrical activity of stem cell–derived neurons. (
COUP-TF1 and THR expression, and COUP-TF1 knockdown
COUP-TF1 and THR mRNA levels were determined at the various stages of mES cell differentiation (Fig. 3). COUP-TF1 mRNA was expressed in mES cells at baseline (day 0) and peak expression was observed on day 18. THRα1 mRNA was expressed in mES cells at baseline and increased with further differentiation and showed peak levels on day 30. In order to determine the role of COUP-TF1 in neuronal differentiation, COUP-TF1 mRNA expression was reduced utilizing shRNA-COUP-TFI lentiviral particle–transduced mES cells. COUP-TF1 mRNA was reduced by 76% at baseline and 50% at the peak level of COUP-TF1 mRNA expression on day 18 of differentiation compared to the control cells (Fig. 4).
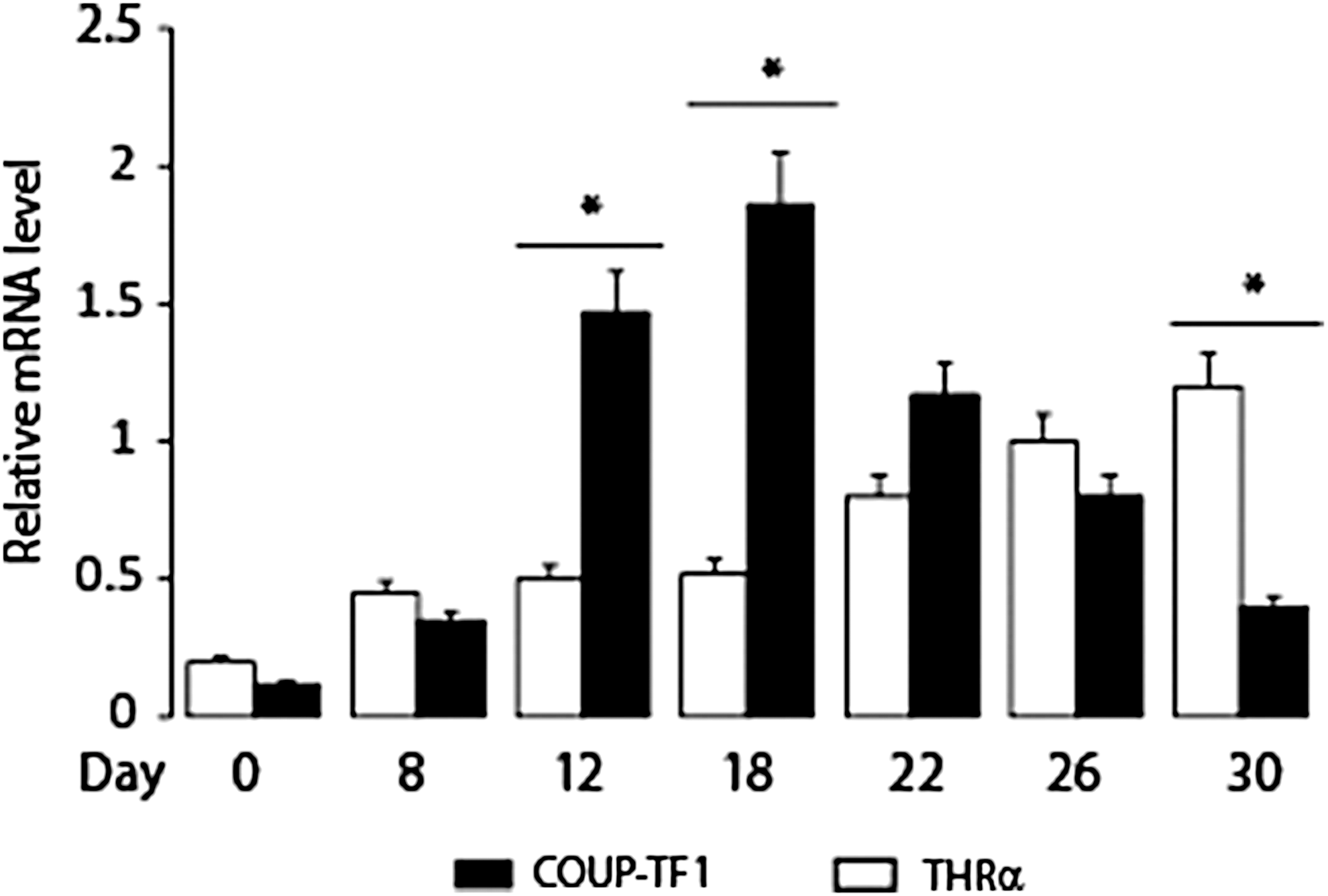
Expression of chicken ovalbumin upstream-transcription factor 1 (COUP-TF1) and thyroid hormone receptor (THRα1) mRNA during mES cell neuronal differentiation. Mouse ES cells were differentiated as described (see legend to Fig. 1). Quantitation of COUP-TF1 and THRα1 mRNA during in vitro mES cell differentiation. Day of in vitro differentiation is shown. *p < 0.05 compared to baseline level peak expression for COUP-TF1 and THRα1 mRNA.
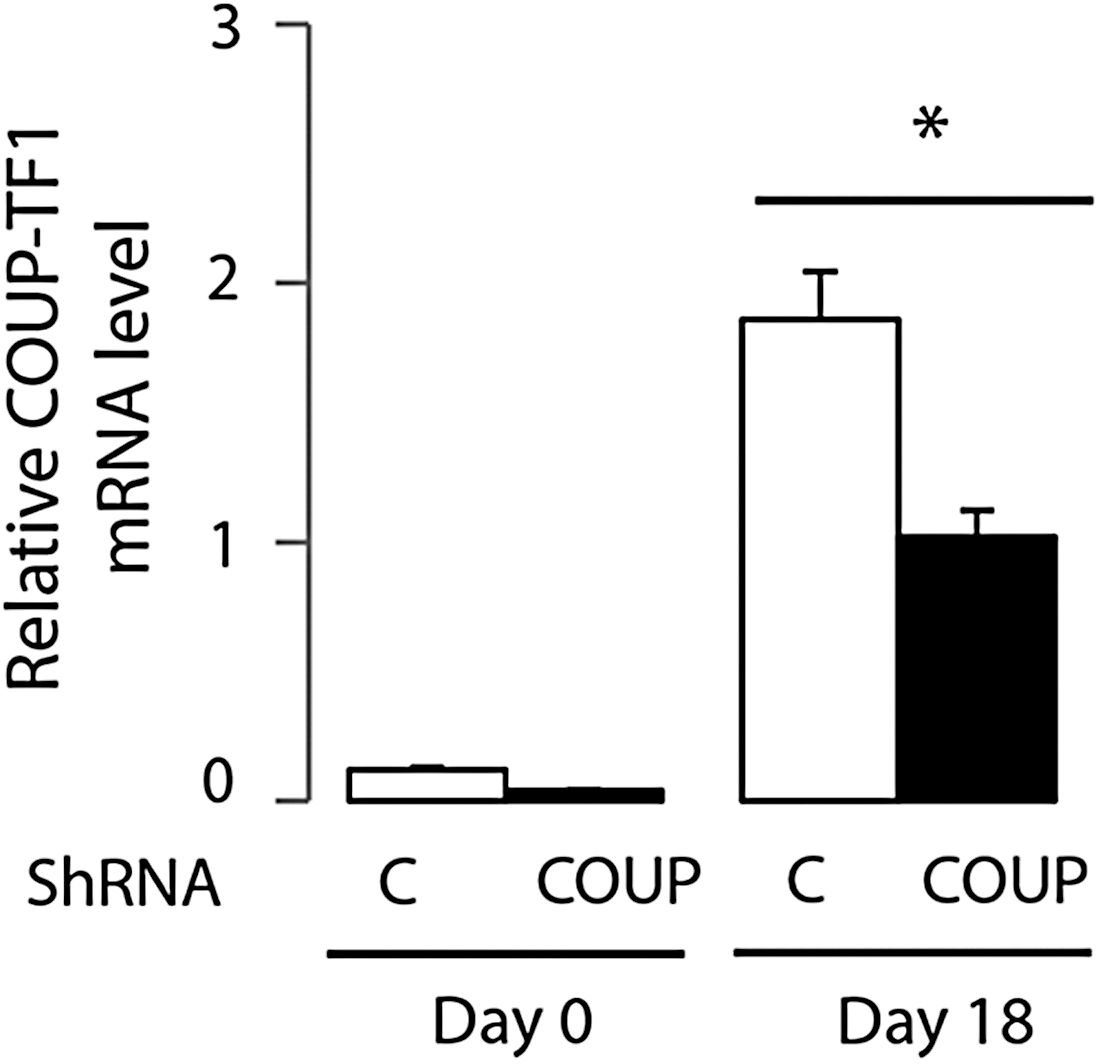
Expression of COUP-TF1 mRNA during mES cell neuronal differentiation, with and without shRNA knockdown. Mouse ES cells were infected with a lentivirus vector expressing control shRNA (C) and COUP-TF1 shRNA knockdown (COUP), and resistant clones were selected after several days of puromycin treatment. Quantitative real-time polymerase chain reaction (PCR) analysis of COUP-TF1 mRNA expression was performed on day 0 and day 18. Values shown are mean ± SE (standard error; error bars) of triplicates in different experiments. A two-tailed Student's t-test was performed to compare means. *p < 0.05, control vs. COUP-TF1 knockdown at day 18.
T3 enhances differentiation of nestin-positive neuronal progenitors
Mouse ES cells were plated in monolayer culture, and T3 was added two days later. In response to T3 (1 nM) treatment, the cell number increased by day 8, including both nestin-positive and -negative cells (Fig. 5A). The number of nestin-positive progenitors in the T3 treatment and nontreatment conditions was compared by fluorescence-activated cell sorting (FACS) analysis (Fig. 5B), and expression of nestin mRNA by real-time PCR (Fig. 5C). T3 treatment (1 nM) enhanced the number of nestin-positive neuronal progenitors on day 8 compared to control. The in vitro study suggests that Nestin may be a T3-responsive gene. The expression of other telencephalic progenitor marker genes, such as Emx2 and Pax-6, was tested, but these were not induced by T3 treatment (data not shown).
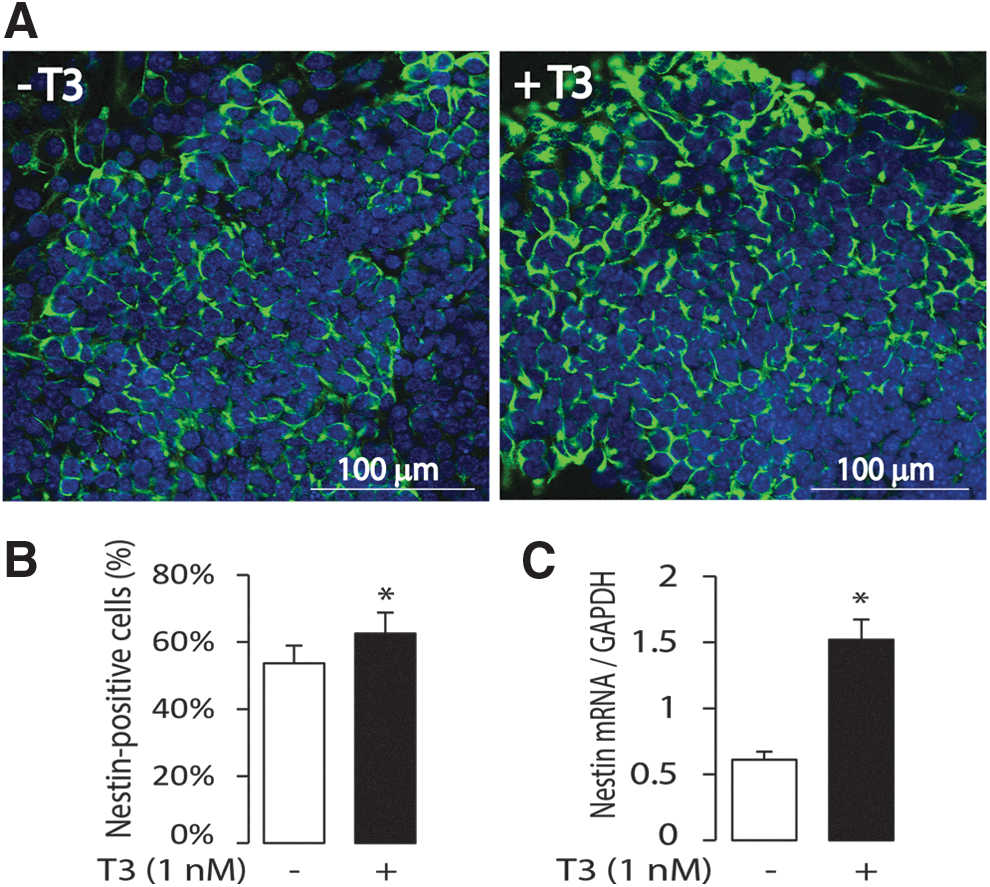
Influence of T3 on neuronal progenitor differentiation. (
T3 stimulates expression of pyramidal neuron markers
The neuronal progenitor clusters were trypsinized on day 12 of differentiation and re-plated, with or without the addition of T3. At stage 2 of neuron differentiation, the progenitors began to differentiate into pyramidal neurons and GFAP-positive astrocytes. The extent of neuronal differentiation was characterized on each day to detect the time course of T3 action. The number of pyramidal neurons increased in response to T3 treatment, from day 12 to day 22, and the influence of T3 on increasing the number of pyramidal neurons on day 22, based on staining for β-tubulin-3, is shown in Figure 6A. The neuronal cells were quantified on day 22 using FACS analysis. T3 enhanced the number of β-tubulin 3-positive neurons compared to expression without T3 (57.7% vs. 46.6%; p < 0.05; Fig. 6B). Several pyramidal neuron and glia cell marker genes were selected (Supplementary Fig. S1), including the pyramidal neuron markers: Neuroligin1, Emx1 (Empty Spiracles Homeobox 1), Emx2 (Empty Spiracles Homeobox 2, pyramidal progenitor and early pyramidal neuron marker); Tbr1 (T-box brain 1, early-born postmitotic pyramidal neuron marker), Reln (Cajal-Retzius marker); β-tubulin-3 (early neuron marker); oligodendrocyte markers: Mbp (Myelin basic protein) and Olig-2 (Oligodendrocyte transcription factor-2); and astrocyte markers: S100B, Gfap, and Glast (glial high affinity glutamate transporter, Slc1a3). mRNA expression was measured by real-time PCR. Several known T3-responsive genes (Rc3 neurogranin, Oct-1, and NSP-A) important for cerebral cortex development were also tested. Among these genes, the expression of mRNA from two cortical neuron-specific genes, Tbr1 and Rc3, was significantly increased by T3 treatment at day 22 (Fig. 6C and D; p < 0.05), consistent with the increased number of pyramidal neurons. These findings demonstrate a stimulatory effect of T3 on corticogenesis.
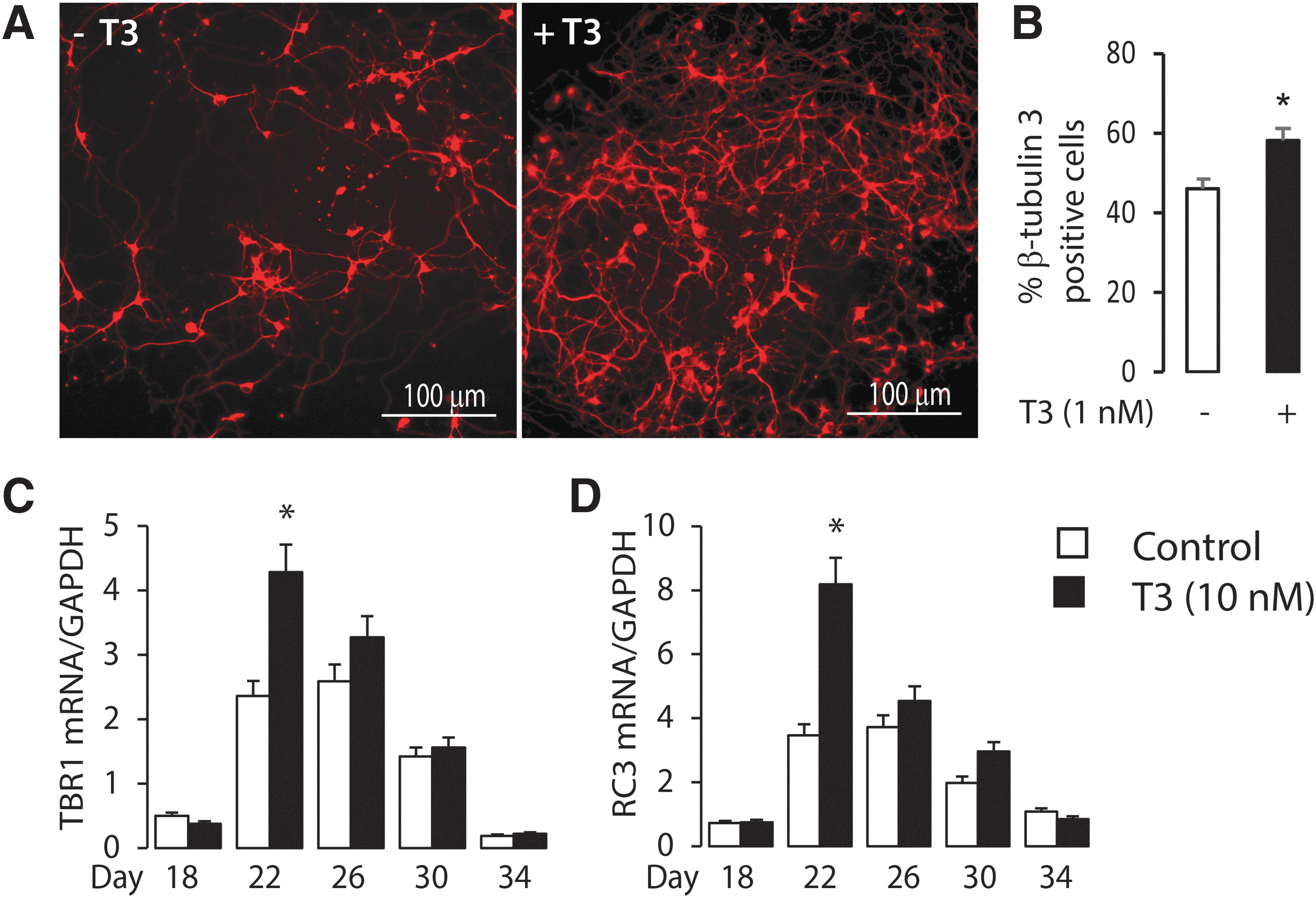
Influence of T3 treatment on pyramidal neuron differentiation. (
COUP-TF1 knockdown results in apoptosis of progenitors and early neurons during the first stage of mES cells differentiation
Mouse ES cells were infected with a lentivirus encoding control shRNA and COUP-TF1 shRNA, designated as ES-CT (control) and ES-KD (knockdown), respectively. In the ES-KD dishes, compared to the ES-CT dishes, there was increased cell death, beginning on day 4 of differentiation. The dead cells included both neuronal and non-neuronal cells. The cell survival rate, as assessed by the XTT kit, was tested after COUP-TF1 knockdown, and it was found that cell survival was reduced to 30% of that shown in the control group (Fig. 7A). The COUP-TF1-induced apoptosis was further evaluated by determining the expression of apoptosis marker genes. COUP-TF1 knockdown enhanced the expression of mRNA from genes important for apoptosis, including Bax, Cas-3, Case-9, and APaf1 (Fig. 7B and C). The effect of COUP-TF1 on the number of neuronal progenitors (nestin+) and early neurons (β-tubulin-3+) was further tested. Flow cytometry was used to identify and quantify the nestin+ neuronal progenitors and β-tubulin-3+ neurons. COUP-TF1 knockdown decreased both the nestin+ neuronal progenitors and β-tubulin-3+ neurons (Fig. 7D and E). T3 treatment in the first stage of differentiation increased nestin+ neuronal progenitors in both ES-CT and ES-KD. T3 treatment increased the nestin+ neuronal progenitors in ES-KD more than in ES-CT, consistent with greater T3 sensitivity in the absence of COUP-TF1 (Fig. 7D). These data demonstrate that both COUP-TF1 and T3 promote the differentiation of the nestin+ neuronal progenitors.
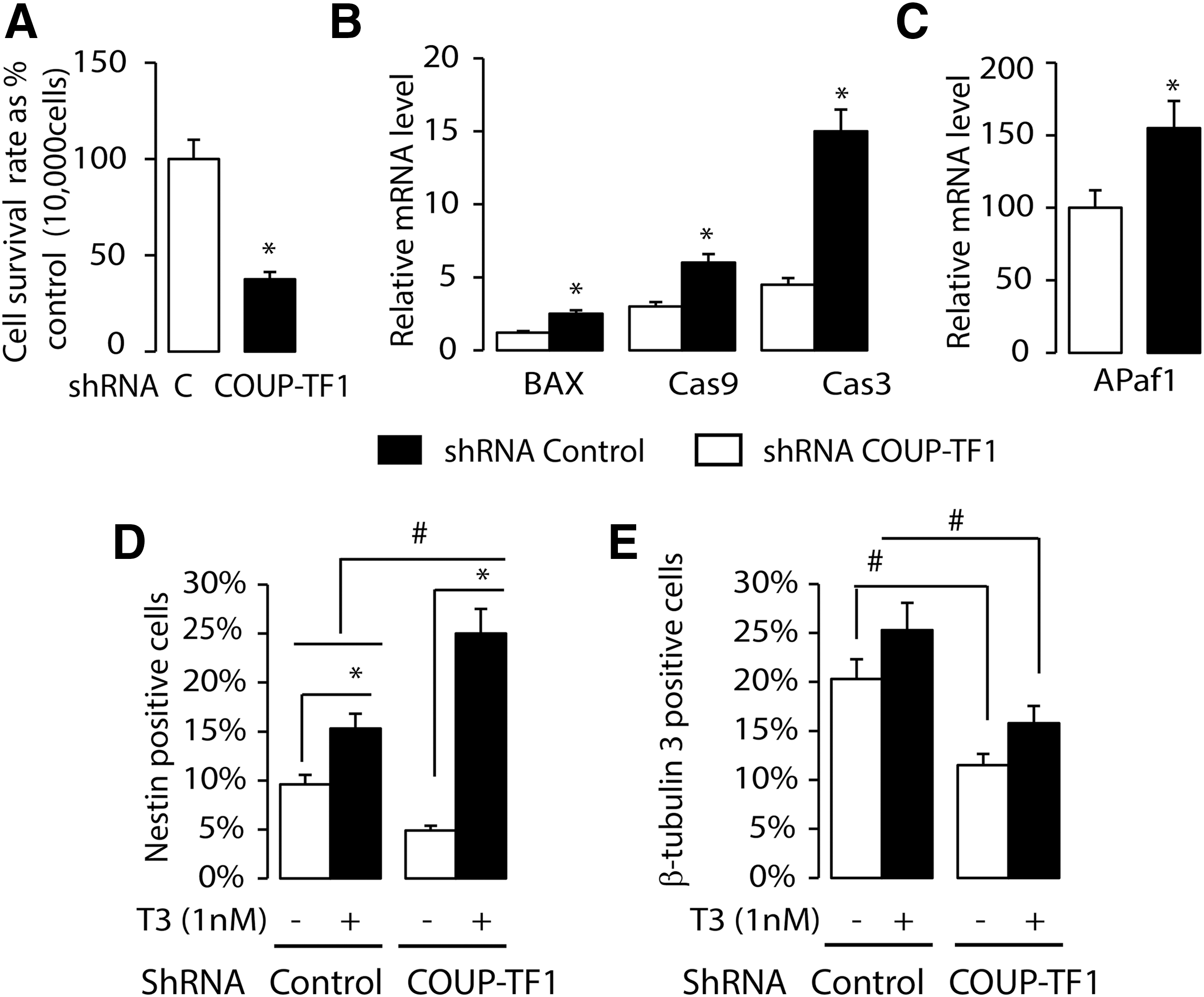
The influence of COUP-TF1 knockdown and T3 on neuronal proliferation and apoptosis. (
COUP-TF1 knockdown accelerates the initiation and the differentiation of pyramidal neurons and enhanced T3-responsive gene expression
The neuronal progenitors were re-plated in differentiation medium and the extent of neural differentiation assessed at day 18. The ES-KD group differentiated earlier than the ES-CT group, as shown by a greater fraction of VGLUT1-positive cells in the COUP-TF1 knockdown (p < 0.05; Fig. 8). mRNA was analyzed from three genes expressed by pyramidal neurons (Tbr1, Rc3, and Emx1) and Camkiv (previously reported to be co-regulated by COUP-TF1 and THR; Fig. 9) (19). COUP-TF1 knockdown resulted in earlier peak expression of all four genes on day 18 compared to the control (Fig. 9A–D), although with different patterns. Rc3 enhancement by COUP-TF1knockdown and T3 treatment on day 18 is consistent with findings that rc3 gene expression is co-regulated by COUP-TF1 and T3 at specific time points. Tbr1 expression was enhanced by COUP-TF1 knockdown on day 18, with T3 treatment. Camkiv and Emx1 mRNA expression were enhanced by COUP-TF1knockdown, but not by T3 treatment in the control, suggesting that other transcription factors required for pyramidal neuron differentiation may be modified by COUP-TF1. The nature of the ES-derived occipital cortical pyramidal neurons derived in this study may differ from the previous models studied, in which T3 stimulated Camkiv gene expression (19).
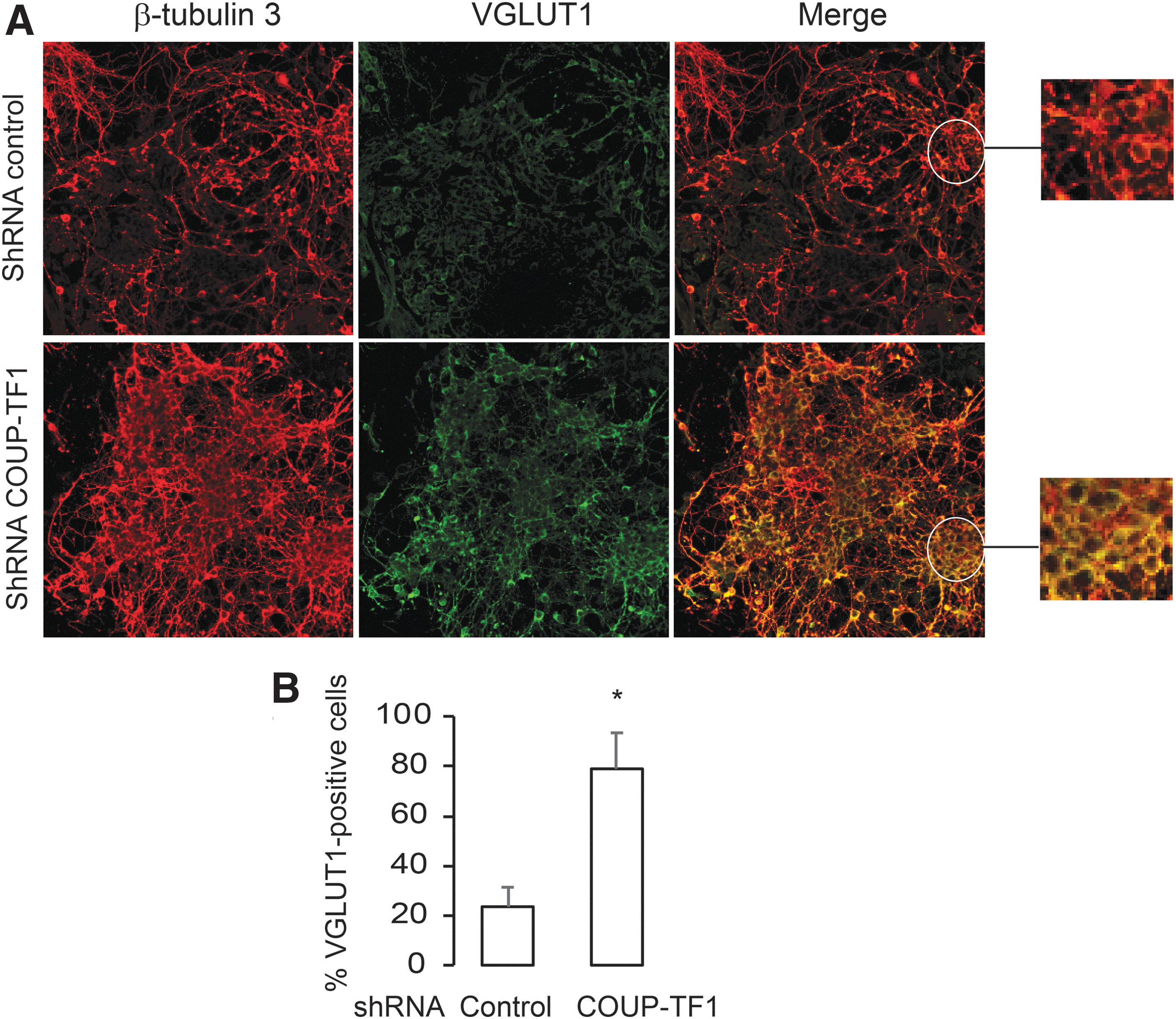
The influence of COUP-TF1 knockdown on the differentiation of pyramidal neurons. (
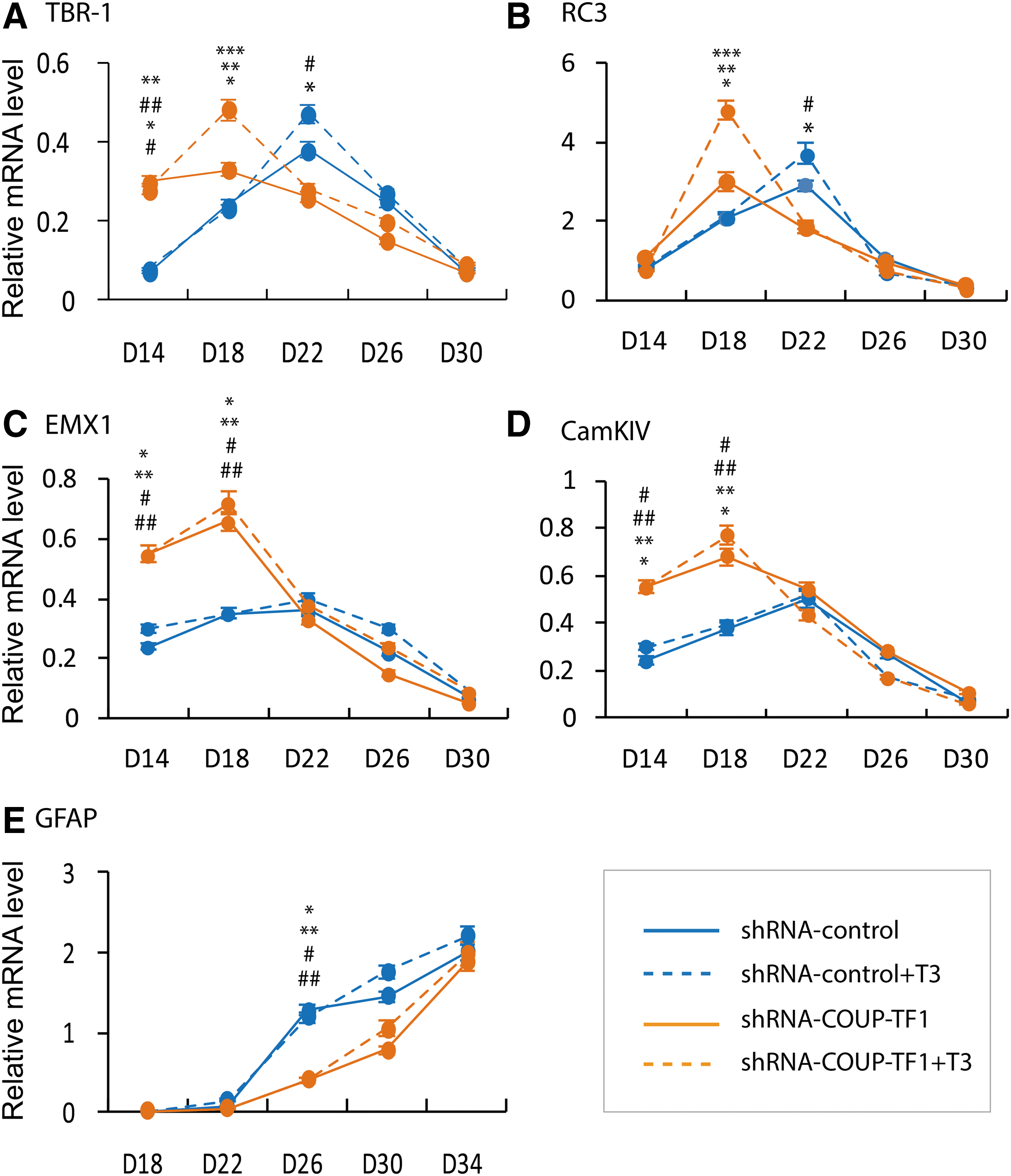
The effects of T3 treatment and COUP-TF1 knockdown on pyramidal neuron marker genes, T3-responsive genes, and astrocyte marker genes. Mouse ES cells, with and without COUP-TF 1 knockdown, were cultured as described in the legend to Figure 1. T3 (10 nM) was added on day 12, and cells were grown for as long as 34 days. Cells were taken at intervals, as shown, to extract mRNA. Quantitative real-time PCR analysis of mRNA was performed after COUP-TF1 knockdown and mRNA expressed as arbitrary units, with and without T3 treatment. Pyramidal neuron marker gene mRNA: (
In contrast to the finding in pyramidal neurons, COUP-TF1 knockdown delayed the initiation of GFAP mRNA expression in glial cells (Fig. 9E). This result is consistent with another in vitro study in which knockdown of both COUP-TF1 and COUP-TFII inhibited the initiation of gliogenesis (27). Therefore, COUP-TF1 has both inhibitory and stimulatory actions in brain development, inhibiting neurogenesis and stimulating the initiation of gliogenesis.
Discussion
Thyroid hormone plays an essential role in mammalian brain development (28), and COUP-TF1 has been implicated in the control of thyroid hormone action (18). An ES cell differentiation model was utilized to determine the role of COUP-TF1. A specific pattern of T3-responsive gene expression is associated with pyramidal neuron differentiation. COUP-TF1 plays an important role in modulating the timing of T3-stimulated gene expression required for corticogenesis. COUP-TF1 knockdown promoted apoptosis of neuronal progenitors and the premature differentiation of pyramidal neurons.
Cerebral corticogenesis is a tightly regulated developmental process involving neuronal progenitor proliferation, neuronal differentiation, and migration (29,30). Previous studies have reported that corticogenesis is regulated by T3. The earliest post-mitotic neuronal cells that give rise to the pre-plate in mice are derived from progenitor cells in the ventricular zone on embryonic day 11.5. On embryonic day 13.5, the cortical neurons migrate radially along radial glial fibers toward the pial surface. When they arrive in the pre-plate, they split it into an upper marginal zone and a lower sub-plate layer. These earliest cortical plate neurons form the deepest layer (layer V and layer VI) of the cortical plate. The latter cortical plate neurons migrate past the previous layers and reside below the marginal zone to form the superficial zone.
In situ hybridization experiments indicate that COUP-TF1 expression can be detected at around embryonic day 8.5. Expression of COUP-TF1 peaks in the forebrain around embryonic day 13.5, and declines before birth (1). COUP-TF1 mRNA expression in the mES cells in vitro differentiation model peaked at day 18 of differentiation, corresponding to embryonic day 13.5 in the in vivo studies.
THR subtypes have distinct patterns of spatiotemporal expression in the developing brain, cochlea, and retina. THRα1 is highly expressed in the ventricular zone, intermediate zone, and cortical plate of the developing cortical cortex (31,32). In contrast to COUP-TF1 expression, THRα1 expression in this in vitro study increased with neuronal differentiation.
A specific pattern of T3-responsive gene expression is associated with pyramidal neuron differentiation
It was found that T3 treatment accelerated pyramidal neuronal differentiation, including increasing both the numbers of neurons and the connections between the pyramidal neurons from day 18 to day 22 of differentiation. Several cortical pyramidal neuron marker genes and known T3-responsive genes were tested. The expression of Tbr1 and Rc3 mRNA was responsive to thyroid hormone stimulation on day 22. Rc3/Neurogranin is a well-characterized T3-responsive gene (33,34). Rc3 mRNA was previously reported to be expressed on embryonic day 14 in the cortex and regulated by thyroid hormone from embryonic day 16 onwards, as well as in adults (35). These findings are consistent with the in vitro study, showing that after day 18 (corresponding to embryonic day 13.5), Rc3 mRNA was responsive to T3 treatment. Tbr1 is expressed in the glutamergic neurons of the developing cerebral cortex, mainly in the pre-plate and layer VI neurons. Tbr1 is important for neuronal differentiation and axon guidance in the developing neocortex (36). These findings suggest that Tbr1 may be a T3-responsive gene.
COUP-TF1 plays an important role in modulating the timing of T3-stimulated gene expression required for corticogenesis
To detect whether COUP-TF1 modulates T3 action in cortical neuron differentiation, stable mES cell lines with COUP-TF1 knockdown were generated. It was found that reduced COUP-TF1 mRNA expression accelerated the differentiation of pyramidal neurons compared to control. At the same time, pyramidal neuron marker genes and T3-responsive genes were analyzed. COUP-TF1 knockdown enhanced the expression of Tbr1, Emx1, and Camkiv mRNA on day 14, with or without T3 treatment, indicating that apart from THR, other transcription factors required for the regulation of these genes are also influenced by COUP-TF1. Fgf signaling is important for cortical development and stimulates cortical progenitors. This signaling pathway is activated by Sp8 and inhibited by COUP-TF1 (37).
These data are consistent with COUP-TF1 playing an important role in preventing premature development of pyramidal neurons. COUP-TF1 knockdown enhanced T3-responsive gene expression (Rc3 and Tbr1) on day 18 (the peak expression time of COUP-TF1). In addition to a more generalized inhibitory role in neuronal differentiation, COUP-TF1 specifically repressed T3-responsive genes required for corticogenesis. COUP-TF1 plays an important role in modulating the timing of T3-stimulated gene expression required for corticogenesis. A direct mechanism of COUP-TF1 interference with THR binding to its TRE has been demonstrated for several T3-responsive genes (2,16), but other mechanisms may also be involved in repression of the non T3-regulated genes.
COUP-TF1 knockdown promotes apoptosis of neuronal progenitors and the premature differentiation of pyramidal neurons
The in vitro COUP-TF1knockdown model also demonstrated an effect of COUP-TF1 promoting neuronal progenitor apoptosis. Previous COUP-TF1 studies reported that COUP-TF1 knockdown resulted in a 30% increase of progenitors in the ventral regions of the pallium, but resulted in excessive progenitor cell death in the glossopharygeal ganglia (38). As described above, the mES cell-derived pyramidal neurons have properties corresponding mainly to those found in the occipital/visual cortex (21). The previous report of cortex-specific deletion of COUP-TF1 resulted in the marked compression of the occipital cortex (6). These data support that COUP-TF1 regulation of neuronal progenitors occurs in a region-specific manner.
Another finding of this study is that COUP-TF1 knockdown can result in premature differentiation of pyramidal neurons, as evidenced by earlier expression of pyramidal neuron marker genes. A previous study reported that COUP-TF1 null mice had abnormal neuronal radial migration in the subventricular zone (SVZ) and intermediate zone (IZ) of the somatosensory cortex, with overexpression of Tbr1 at embryonic day 16.5 (39). It is speculated that the premature development of pyramidal neurons resulting from COUP-TF1 knockdown may model the abnormal pattern of cortical pyramidal layers in the COUP-TF1 null mutant mouse. Crucial developmental “windows” for the inactivation and activation of T3 by the 5-deiodinase type 3 and 5′-deiodinase type 2 have been well described for sensory development (40).
T3 and COUP-TF1 both enhanced the differentiation of nestin+ progenitors and increased the expression of Nestin mRNA. COUP-TF1 inhibits most T3-stimulated genes and enhances T3-inhibited genes, but Nestin mRNA was enhanced by both. This finding suggests that there is no direct action of COUP-TF1 on T3 signaling.
In conclusion, this study found that T3 treatment promoted differentiation of mES cells into neuronal progenitors and cortical pyramidal neurons. T3 action on the known T3-responsive genes, Rc3 and Tbr1, required for pyramidal neuron differentiation, occurred in a specific temporal pattern. COUP-TF1 knockdown resulted in the premature maturation of pyramidal neurons, accompanied by the earlier peak expression of the pyramidal neuron marker genes (Rc3, Tbr1, and Emx1) and a T3-responsive gene (Camkiv). COUP-TF1 plays an important role in modulating the timing of T3-responsive gene expression required for corticogenesis, in part by modulating sensitivity to thyroid hormone action. This in vitro model demonstrates many features of the in vivo studies, but provides the opportunity for to determine specific mechanisms of COUP-TF1 and T3 regulation of neuronal differentiation and function.
Footnotes
Acknowledgments
This work was supported by Veterans Affairs Merit Review Grant (to G.A.B.), NIH RO1 DK98576 (to G.A.B.), and National Science Foundation of China Grant 81570711 (to X.T.).
Author Disclosure Statement
The authors have nothing to disclose.