
Abstract
Background:
Thyroid hormones (THs) are key regulators of development, tissue differentiation, and maintenance of metabolic balance in virtually every cell of the body. Accordingly, severe alteration of TH action during fetal life leads to permanent deficits in humans. The skin is among the few adult tissues expressing the oncofetal protein type 3 deiodinase (D3), the TH inactivating enzyme. Here, we demonstrate that D3 is dynamically regulated during epidermal ontogenesis.
Methods:
To investigate the function of D3 in a postdevelopmental context, we used a mouse model of conditional epidermal-specific D3 depletion. Loss of D3 resulted in tissue hypoplasia and enhanced epidermal differentiation in a cell-autonomous manner.
Results:
Accordingly, wound healing repair and hair follicle cycle were altered in the D3-depleted epidermis. Further, in vitro ablation of D3 in primary culture of keratinocytes indicated that various markers of stratified epithelial layers were upregulated, thereby confirming the pro-differentiative action of D3 depletion and the consequent increased intracellular triiodothyronine levels. Notably, loss of D3 reduced the clearance of systemic TH in vivo, thereby demonstrating the critical requirement for epidermal D3 in the maintenance of TH homeostasis.
Conclusion:
In conclusion, our results show that the D3 enzyme is a key TH-signaling component in the skin, thereby providing a striking example of a physiological context for deiodinase-mediated TH metabolism, as well as a rationale for therapeutic manipulation of deiodinases in pathophysiological contexts.
Introduction
The skin protects organisms from infection, prevents dehydration, and undergoes re-epithelialization after wound injury (1). In mammals, epidermal development is a multistage process consisting of epidermal specification, commitment, stratification, and terminal differentiation (1). To accomplish these steps, the epidermis constantly replenishes itself thanks to the presence of stem cells that are capable of self-renewal and of producing transiently amplifying progenitor cells. Dividing cells in the innermost (basal) epidermal layer continually execute a program of terminal differentiation, move outward, exit the cell cycle, and embark on a terminal differentiation pathway as they migrate toward the skin surface (1). During the whole process, distinct signaling patterns specify different developmental stages and these stage-specifically regulated signaling events ensure the correct morphogenesis of skin epidermis and its appendages (2). Basal layer keratinocytes express the keratins K5 and K15. As cells exit from the basal layer and begin their journey toward the skin surface, they cease expressing K15 and K5 and start expressing K1 and K10 (3,4). The first suprabasal cells are known as “spinous cells,” accordingly to their cytoskeleton content of K1/K10 filaments connected to robust cell–cell junctions; K6, K16, and K17 are also expressed suprabasally, but only in hyperproliferative situations such as wound healing (WH) (3,4).
The skin is a well-recognized target of thyroid hormone (TH), which is an important regulator of epidermal homeostasis (5). In the skin, TH exerts profound effects on fetal epidermal differentiation, barrier formation, hair growth, WH, keratinocyte proliferation, and keratin gene expression (6). Moreover, several TH responsive genes have been identified in the skin. For example, TH differentially regulates the K5, K14, and K17 gene promoters and the hairless gene (Hr) is a well-known TH-target gene in keratinocytes (7).
Although various studies have provided insights into how TH regulates specific stages of epidermal homeostasis (8,9), a few have examined the effects of an altered TH signal at a cell-specific level, and the consequence of imbalanced TH homeostasis in physiological conditions.
Three selenoenzymes, the deiodinases, play a crucial role in the control of TH intracellular availability. These enzymes metabolize THs in a stage- and tissue-specific manner by a mono-deiodination reaction that involves two distinct pathways. Type 1 and 2 deiodinases (D1 and D2) catalyze the conversion of the precursor thyroxine (T4) to the more active form triiodothyronine (T3)—a process that increases circulating T3 levels and the availability of the active hormone for the nuclear receptors (10). In contrast, type 3 deiodinase (D3) inactivates TH by converting T4 and T3 to the inactive metabolites reverse T3 (rT3) and T2, respectively. Notably, the combined actions of D2 and D3 are viewed as a cell autonomous prereceptoral mechanism that controls TH signaling in a time- and tissue-specific manner although it does not affect serum hormone concentrations (11 –13).
D3—the main physiologic inactivator of TH—controls TH homeostasis by protecting tissues from an excess of TH (14). It is abundantly present in fetal tissues and high D3 levels are expressed in the human placenta, where it blocks the excessive maternal-to-fetal transfer of T4 (15). In contrast, during late neonatal and adult life, D3 expression is more limited and present in the skin, brain, ovary, and the nonpregnant and pregnant uterus (16 –18). Notably, D3 is expressed in several pathophysiological conditions, among which are cardiac hypertrophy, chronic inflammation, critical illness (19), hypoxia, ischemia (20), and cancer. We and others demonstrated that D3 is upregulated in basal cell carcinoma (21), colon carcinoma (22), hemangiomas (23), astrocytomas, and glioblastomas (24). Because D3 is expressed in fetal and malignant tissues, it is referred to as an “oncofetal enzyme.”
Despite evidence that TH plays an important role in epidermal proliferation, the physiologic role of TH and its local modulation by D3 in skin is not well understood. Here, we investigated the physiological role of D3 in the maintenance of the proliferation/differentiation balance of keratinocytes. In vitro and in vivo experiments demonstrated that epidermal depletion of D3 dramatically alters the proportion of the epithelial layers, thus resulting in a drastic reduction of the skin thickness. Loss of D3 in the epidermis, by reducing the percentage of epidermal stem cells, impairs a correct closure of the wound after an injury and alters the execution of the hair follicle cycle. Notably, the clearance of T4 is reduced in the absence of D3 in the skin, thus revealing a critical role for D3 in the homeostatic control of local and systemic TH levels.
Materials and Methods
Cell cultures
Mouse primary keratinocytes were isolated from C57BL/6 newborn mice and cultured under low-calcium conditions (0.05 mM) in the presence of 4% calcium-chelated fetal bovine serum (FBS) (Invitrogen) and epidermal growth factor (Invitrogen), as previously described (25). For depletion of D3 in primary culture of mouse keratinocytes, adenoviral transduction of Cre-recombinase was performed as previously described (26). Freshly isolated mouse primary keratinocytes from K14CREERT−/− ; D3fl/fl and K14CREERT+/− ; D3fl/fl mice were incubated in suspension with control adenoviruses (Ad-IRES-GFP [Ad-GFP]; Vector Biolab) or adenoviruses expressing Cre recombinase (Ad-Cre-IRES-GFP [Ad-Cre]; Vector Biolab) at a multiplicity of infection of 50 for 1 hour at 37°C. To evaluate proliferation, cells were incubated with BrdU labeling reagent (B-5002 Sigma) for two hours before the harvest. For immunofluorescence staining, cells were fixed with 4% formaldehyde, permeabilized with 50 mM NaOH, blocked with 5% goat serum, and incubated with monoclonal Ab G3G4 (Developmental Studies Hybridoma Bank) and DNase. Alexa Fluor 595-conjugated secondary antibody was used. The cell DNA was stained with 300 nM 4′,6-diamidino-2-phenylindole (Molecular Probes; Invitrogen).
Animals, histology, and immunostaining
D3KO (K14CreERT-D3fl/fl ) mice were obtained by crossing the keratinocyte-specific conditional K14CreERT mouse (27) with D3fl/fl (25) mice. Depletion was induced in 6-day-old male mice by treatment with 10 mg of tamoxifen at different time points, as indicated in each experiment. K14CreERT-D3fl/fl ; Gt (ROSA)26Sortm4(ACTB-tdTomato,-EGFP)Luo were obtained by crossing K14CreERT-D3fl/fl with Gt(ROSA)26Sortm4(ACTB-tdTomato,-EGFP)Luo (The Jackson Laboratory). Mice were collected at postnatal day 30 for analysis of epidermal morphology.
For immunofluorescence and histology, dorsal skin from D3KO and control mice was embedded in paraffin, cut into 7-μm sections, and hematoxylin and eosin (H&E)-stained. Slides were baked at 37°C, deparaffinized by xylenes, dehydrated with ethanol, rehydrated in phosphate-buffered saline (PBS), and permeabilized by placing them in 0.2% Triton X-100 in PBS. Antigens were retrieved by incubation in 0.1 M citrate buffer (pH 6.0) or 0.5 M tris buffer (pH 8.0) at 95°C for 5 minutes. Sections were blocked in 1% bovine serum albumin/0.02% Tween/PBS for 1 hour at room temperature. Primary antibodies were incubated overnight at 4°C in blocking buffer and washed in 0.2% Tween/PBS. Secondary antibodies were incubated at room temperature for 1 hour, and they were washed in 0.2% Tween/PBS. Images were acquired with an IX51 Olympus microscope and the Cell*F Olympus Imaging Software.
For epidermal thickness analysis, 7-μm serial paraffin sections were stained with H&E. For assessment of 5-ethynyl-2′-deoxyuridine (EdU) incorporation, mice were intraperitoneally (i.p.) injected with 10 mg/mL EdU (E10415; Invitrogen) into the abdominal cavity. Two hours after EdU injection, mice were euthanized and the dorsal skin was formalin fixed and paraffin embedded as previously described. Immunostaining was performed with Click-iT™ EdU Cell Proliferation Kit (C10339; Thermofisher).
All animal experiments and mouse husbandry were carried out in the animal facility of CEINGE—Biotecnologie Avanzate, Naples, Italy, in accordance with institutional guidelines (Authorization no. 354/2019-PR by the Ministero della Salute).
Hair follicle cycle
Characterization of the hair follicle cycle of CTR and D3KO mice was performed as previously described (28). Briefly, dorsal backs of three-month-old anesthetized D3KO and control mice were shaved by using a clamp. The mice were harvested after 6 days for the anagen, 10 days for the catagen, and 30 days for the telogen analysis; dorsal skin was collected for molecular and histological analysis (29).
Wound preparation, macroscopic examination, and histological analyses
Skin wounds from CTR and D3KO mice were prepared as previously described (30). Briefly, mice were deeply anesthetized with ketamine. After shaving and cleaning with 70% alcohol, excisional, full-thickness skin wounds were aseptically made on the dorsal skin by picking up a fold skin at the midline and using a sterile, disposable biopsy punch with a diameter of 8 mm to punch through the two layers of skin. In this manner, two wounds were made on each side of the midline at the same time. Each wound site was digitally photographed by using the Nikon FX-35A at the indicated time intervals, and wound areas were determined on photographs by using Image J without prior knowledge of the experimental procedures. Changes in wound areas are expressed as the percentage of the initial wound areas. For histological analyses of wound sites, skin wounds were fixed in 4% formaldehyde buffered with PBS (pH 7.2) and were then embedded with paraffin. Sections (6-μm thick) were stained with H&E for histological analysis, and immunofluorescence analyses for keratin 14 (K14) were performed.
Real-time polymerase chain reaction
Messenger RNAs were extracted with Trizol reagent (Life Technologies Ltd.). Complementary DNAs were prepared with Vilo reverse transcriptase (Life Technologies Ltd.), as indicated by the manufacturer. The cDNAs were amplified by polymerase chain reaction (PCR) in an iQ5 Multicolor Real-Time Detector System (BioRad) with the fluorescent double-stranded DNA-binding dye SYBR Green (Biorad). Specific primers for each gene were designed to work under the same cycling conditions (95°C for 10 minutes followed by 40 cycles at 95°C for 15 seconds and 60°C for 1 minute), thereby generating products of comparable sizes (about 200 bp for each amplification). Primer combinations were positioned whenever possible to span an exon–exon junction, and the RNA was digested with DNAse to avoid genomic DNA interference. Primer sequences are indicated in the Supplementary Data. For each reaction, standard curves for reference genes were constructed based on 6 fourfold serial dilutions of cDNA. All samples were run in triplicate. The template concentration was calculated from the cycle number when the amount of PCR product passed a threshold established in the exponential phase of the PCR. The relative amounts of gene expression were calculated with cyclophilin A expression as an internal standard (calibrator). The results, expressed as N-fold differences in target gene expression, were determined as follows: N × target = 2(DCt sample−DCt calibrator). For primer sequences, see Table 1.
Oligonucleotides Used for Real-Time Polymerase Chain Reaction
Protein extraction from skin and Western blot analysis
Dorsal skin was removed from mice and immediately snap-frozen in liquid N2. Then, 800 μL of lysis buffer (0.125 M tris pH 8.6; 3% sodium dodecyl sulfate (SDS), protease inhibitors including phenylmethylsulfonyl fluoride 1 mM and phosphatase inhibitors) was added to all dorsal skin samples, which were then homogenized with Tissue Lyser (Qiagen). Total protein extracts from cells and from skin tissue were run on a 10% sodium dodecyl sulphate-polycrylamide gel electrophoresis gel and transferred onto an Immobilon-P transfer membrane (Millipore). The membrane was then blocked with 5% nonfat dry milk in PBS; probed with anti-K5, anti-Loricrin, anti-D3, anti-extracellular signal-regulated kinase (ERK), and anti-tubulin antibodies overnight at 4°C; washed; incubated with horseradish peroxidase-conjugated anti-mouse immunoglobulin G secondary antibody (1:3000); and detected by chemiluminescence (Cat. No. WBKLS0500; Millipore). After extensive washing, the membrane was incubated with anti-Tubulin specific antibodies (Santa Cruz Biotechnology) that served as a loading control. All Western blots were run in triplicate, and bands were quantified with ImageJ software. Antibodies and primer sequences are indicated in Table 2.
List of Antibodies Used for Western Blot and Immunofluorescence Analysis
Dentice et al. (21).
K14, keratin 14.
Isolation of epithelial stem cells
Epithelial stem cells from D3KO and control mice were obtained as previously described (31). Immunostaining was performed by using allophycocyanin (APC)-anti-mouse CD34 (code 119310; Biolegend), PE-rat anti human α6-integrin (code 555736; BD Pharmingen). Fluorescence-activated cell sorting analysis was performed by using FACS Canto2 software (FACS Canto2; Becton Dickinson).
Dorsal skin was removed from mice and treated with trypsin 0.25% overnight after removal of adipose tissue. The epidermal layer was separated from the dermis, chopped, and incubated with trypsin 0.25% for 7 minutes at 37°C. After digestion, FBS was added to the sample to inactivate trypsin. The cells were filtered with a 70-μm cell strainer. Immunostaining was performed by using APC-anti-mouse CD34 (cod 119310; Biolegend), PE-rat anti human α6-integrin (cod 555736; BD Pharmingen), by incubation for 1 hour room temperature. Fluorescence-activated cell sorting analysis was performed by using FACS Canto2 software (FACS Canto2; Becton Dickinson).
T4 clearance experiment
Eight-week-old mice were i.p. injected with 20 μg L-T4/100 g body weight. Starting 1 day before the injection, the blood was collected by retro-orbital collection at 24 and 48 hours after a single L-T4 injection. The sera were separated, and the determination of T4 was measured by Rodent T4 Elisa Test Kit (Endocrine Technologies, Inc.). Experiments were performed with 6 mice/group and repeated twice in these conditions.
High performance liquid chromatography-mass spectrometry measurement of T3, T4, and rT3
Standard stock solutions of THs (3,3′,5,5′-tetraiodo-L-thyronine (L-T4), 3,3′,5-triiodothyronine (T3), and 3,3′,5′-triiodothyronine (rT3) were prepared in methanol. Dilutions of each standard were prepared in methanol/water (v/v, 50/50). Ten milliliters of cell media was deproteinated by using nine volumes of cold acetone and then centrifuged at 14,000 rpm. The supernatants were reduced to 200 μL under N2 for instrumental analysis. The high performance liquid chromatography system Jasco Extrema LC-4000 system (Jasco, Inc., Ithaca, NY) was coupled to an Advion Expression mass spectrometer (Advion, Inc., Ithaca, NY) equipped with an electrospray ionization (ESI) source. Ten millimolars ammonia acetate in deionized water was used as the aqueous mobile phase, and 0.1% acetic acid in methanol was used as the organic mobile phase. The analyses were performed in the positive ESI mode. Six replicates were run for each sample.
Serum thyrotropin (TSH) was measured by ELISA kit (E-EL-M1153; Elabscience) according to the manufacturer's protocol. Experiments were performed with 6 mice/group and repeated twice.
Statistics
The results are shown as means ± standard deviation. Differences between samples were assessed by the Student's two-tailed t test for independent samples. Relative mRNA levels (in which the control sample was arbitrarily set as 1) are reported as results of real-time PCR, in which the expression of cyclophilin A served as housekeeping gene. In all experiments, differences were considered significant when p was less than 0.05. Asterisks indicate significance at *p < 0.05, **p < 0.01, and ***p < 0.001.
Results
Endogenous D3 is dynamically expressed during epidermal development and the hair follicle cycle
To assess the localization of D3 during epidermal development and in adult skin, we analyzed its expression during mouse embryogenesis (E11.5 to E18.5). We found that D3 expression starts at E15.5, peaks at E17.5, and declines thereafter (Supplementary Fig. S1). As shown in Figure 1A–C, D3 expression is highly dynamic in early postnatal life. In fact, it is much lower at P2 than during embryogenesis. It peaks at P10, decreases thereafter, and reaches a plateau in adult life (Fig. 1A–C), in accordance with the previous finding of a neonatal peak in skin D3 at postnatal day 12 (16). Immunofluorescence analysis showed that D3 localizes in all the epidermal layers (Fig. 1A), and is expressed also in hair follicles mainly at P10, where it reaches its highest levels. Interestingly, this profile positively correlates with miR21 expression and negatively with GRHL3 expression (Fig. 1D), both of which have been shown to coordinately regulate D3 expression in skin (25). During the hair follicle cycle, D3 is highly expressed at anagen and catagen, declining in telogen (Fig. 1E, F). To assess the specific localization of D3 in the hair follicle, we analyzed the profile of D3 protein expression in the adult epidermis. Immunohistochemical analysis showed that D3 is highly expressed in the bulge region and in sebaceous gland of the hair follicle and it is absent in the dermal papilla. D3 was also detected in infundibulum cells and in the interfollicular epidermis (Supplementary Fig. S2). Collectively, these results indicate that in normal epidermis, D3 is dynamically expressed in the epidermal layers of the skin and its expression correlates with hair follicle formation. Intriguingly, in normal skin, D3 expression correlates with expression of miR-21/GRHL3, suggesting the existence of a regulatory cascade in the control of epidermal homeostasis.
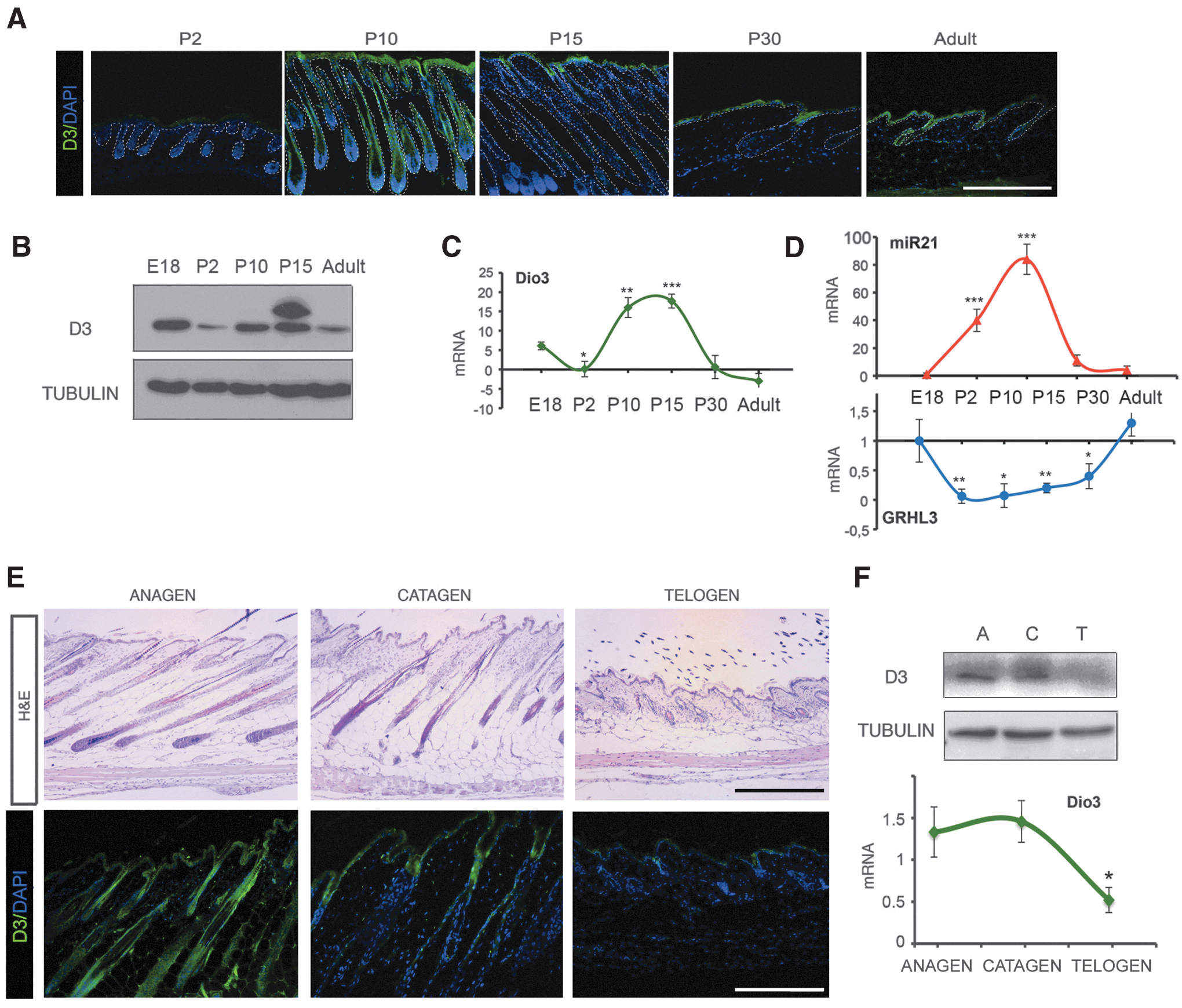
D3 is dynamically expressed in adult epidermis and hair follicle cycle. (
Spatiotemporal D3 genetic depletion accelerates basal cell differentiation
To investigate the functional role of D3 in epithelial homeostasis, we generated an animal model for epidermal D3 loss of function. To deplete D3 in the epidermis, mice in which the Dio3 gene is flanked by Lox-P sequences (32) were crossed with transgenic mice with keratinocyte-specific expression of a CreERT recombinase under the control of the K14 promoter (Supplementary Fig. S3A–C) that is responsible for ubiquitous recombination in hair follicles and basal cells of the epidermis (33). Because D3 expression is dynamically regulated during ontogenesis (Fig. 1A), we depleted D3 at day P6, before the massive induction of D3 in the epidermis (Fig. 2A). Mice with epidermal D3 depletion (D3KO) were euthyroid at systemic levels, as shown in Supplementary Figure S4. Loss of D3 in the epidermal layers resulted in reduced skin thickness (Fig. 2B–G). The number of EdU-positive cells and the expression of the proliferative marker K6 and the basal marker K5 were lower in the epidermis of D3KO mice than in control mice (Fig. 2B–F). Interestingly, Loricrin, which marks the most differentiative layers in the control mice, was higher in D3KO mice and localized not only in the upper layers as in the control mice but also in the lower epidermal layers (Fig. 2C). Together with the fewer EdU-positive cells and reduced thickness, these data are indicative of the accelerated differentiation of the keratinocytes in the D3KO epidermis.
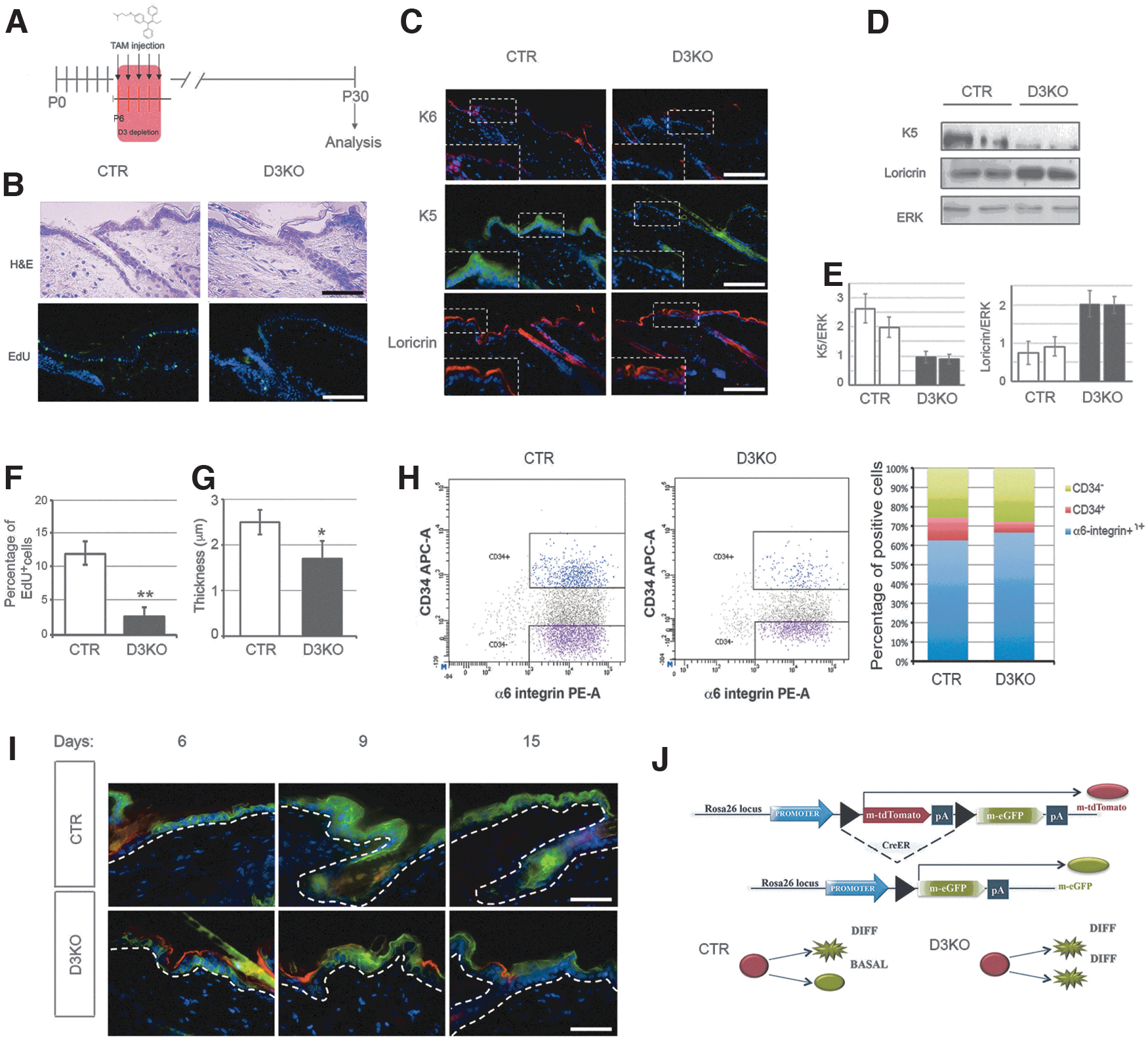
Genetic depletion of D3 in the epidermis reduces the thickness of the skin and increases the expression of differentiative markers. (
Since the basal layer marker K5 is reduced in D3KO mice, we asked whether D3 also affects the stem cell compartment. Therefore, we measured the percentage of CD34+ stem cells by FACS analysis (α6-integrin+/CD34+ cells). As shown in Figure 2H, the number of CD34+ cells was much lower in the D3KO epidermis than in control mice (Fig. 2H).
To perform lineage tracing, we crossed D3KO mice with the double fluorescent Tomato/GFP mouse. We observed that the control mice expressed green cells in both basal and suprabasal layers, as expected by the normal replenishment model of basal keratinocytes. Contrarily, in the D3KO mice, the green cells produced by Cre-mediated Tomato-GFP shift moved rapidly to the upper layers and at day 15 after recombination, were located only in the outer epidermis (Fig. 2I), consistently with the hypothesis that excessive T3 exposure induces premature differentiation of keratinocytes in D3KO mice. These data show that alteration of the thyroid status by reduced expression of D3 induces deregulation of epidermal homeostasis and supports a model in which higher levels of T3 in D3KO epidermis induce keratinocyte differentiation (Fig. 2L).
D3 depletion and TH treatment increases keratinocyte differentiation in vitro
The epidermal in vivo alterations observed may reflect a direct role of D3 and TH in keratinocyte growth/differentiation control. To test the effects of D3 depletion or T3 treatment in the control of the differentiation/proliferation balance, we used primary cultures of keratinocytes. We first observed that D3 is expressed in freshly isolated keratinocytes and that its expression declines during the differentiation induced by increased Ca2+ concentration in the medium (Fig. 3A). D3 was then depleted in primary keratinocytes from D3KO mice infected with an adenovirus carrying the CRE (Ad-CRE) recombinase or with an adeno-GFP (Ad-GFP) virus as control. Loss of D3 caused a reduction in cell proliferation in D3-depleted cells measured by BrdU incorporation (Fig. 3B, C). Accordingly, mRNA expression of cyclin D1 and K14 was reduced while the expression of differentiation markers K10 and involucrin was upregulated (Fig. 3D). Further, D3 depletion induced an alteration of keratinocyte morphology. As shown in Figure 3E, keratinocytes were smaller and round, which suggested spontaneous, early differentiation (Fig. 3E).
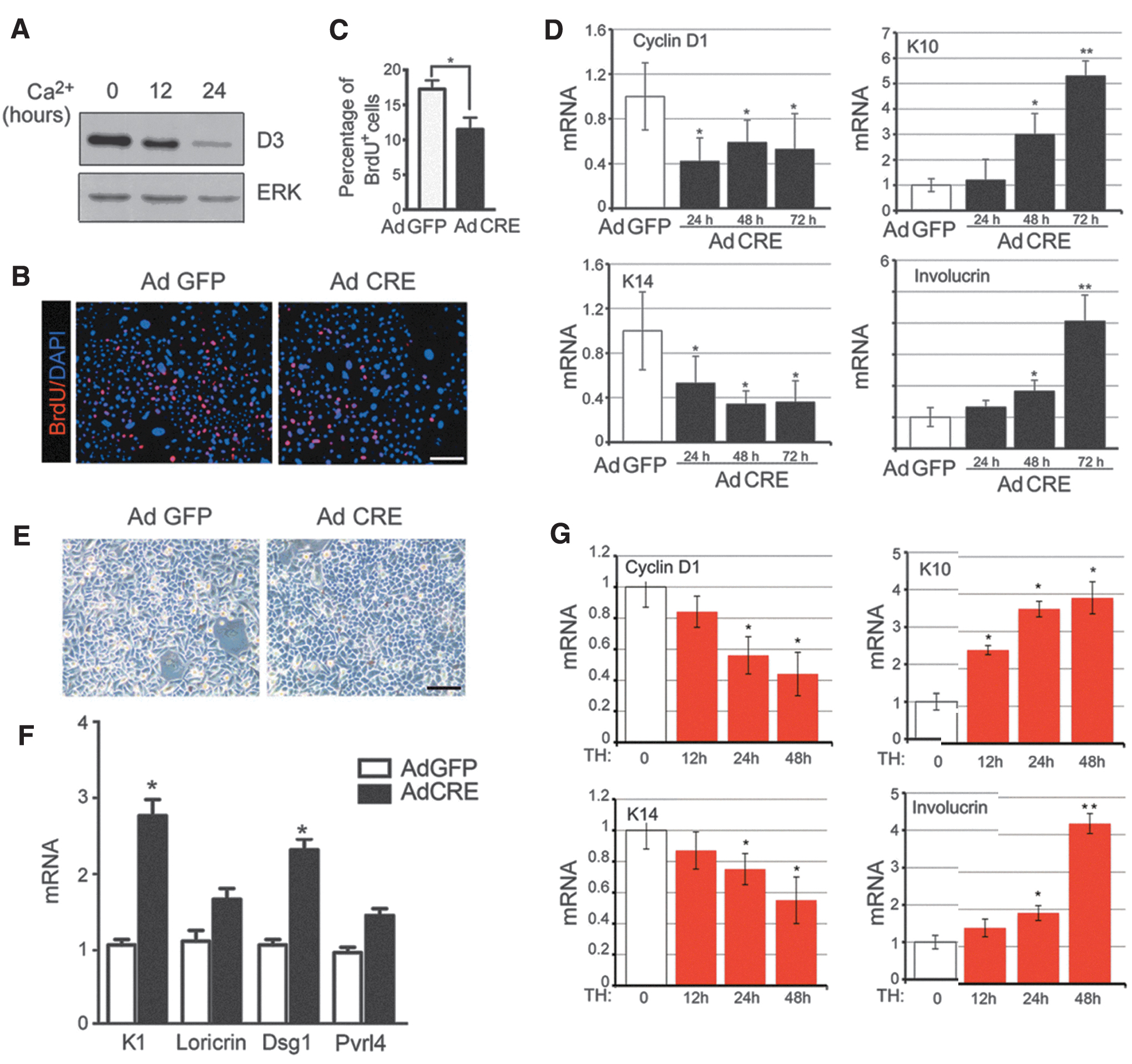
In vitro depletion of D3 and treatment with T3 accelerates differentiation and reduces proliferation of mouse keratinocytes. (
To investigate the differentiation potential of D3KO keratinocytes, we induced cell differentiation by Ca2+ treatment and measured various differentiation markers, namely, K1, loricrin, Pvrl4 [a cell adhesion molecule mainly implicated in the formation of cadherin-based adherens junctions (34)], and desmoglein 1 [Dsg1, an epidermal adhesion molecule (35)] in Ad-CRE- and Ad-GFP-infected keratinocytes. All these markers were upregulated in D3-depleted keratinocytes compared with control keratinocytes (Fig. 3F), which confirms that local TH excess drives the cells toward functional differentiation. Treatment with 30 nM T3 paralleled the effects of D3 depletion in a time-dependent manner, with reduction of cyclin D1 and K14 and upregulation of K10 and involucrin (Fig. 3G).
WH repair is delayed and hair follicle cycle is accelerated in D3KO skin
Having ruled out defects in epithelial commitment and differentiation in epithelia lacking D3, we evaluated the ability of D3KO mice to repair the skin damage induced by a wound. We first examined the expression patterns of D3 after the wound created in the dorsal skin in mice. Three days after full-thickness injury, expression levels of D3 mRNA and protein increased significantly, reached a peak at day 6, and declined thereafter (Fig. 4A, B). Compared with the control mice, D3KO mice exhibited a sharp delay in their regenerative ability as demonstrated by an increased wound area and a delay in the wound closure (Fig. 4C, D). Indeed, while in control mice, wound areas were smaller with linear kinetics during the 8-day period of observation, at the end of which ∼95% closure was observed, wound closure was significantly delayed in D3KO mice, which showed a 60% wound closure at day 6, and only 70% wound closure at day 8 (Fig. 4D).
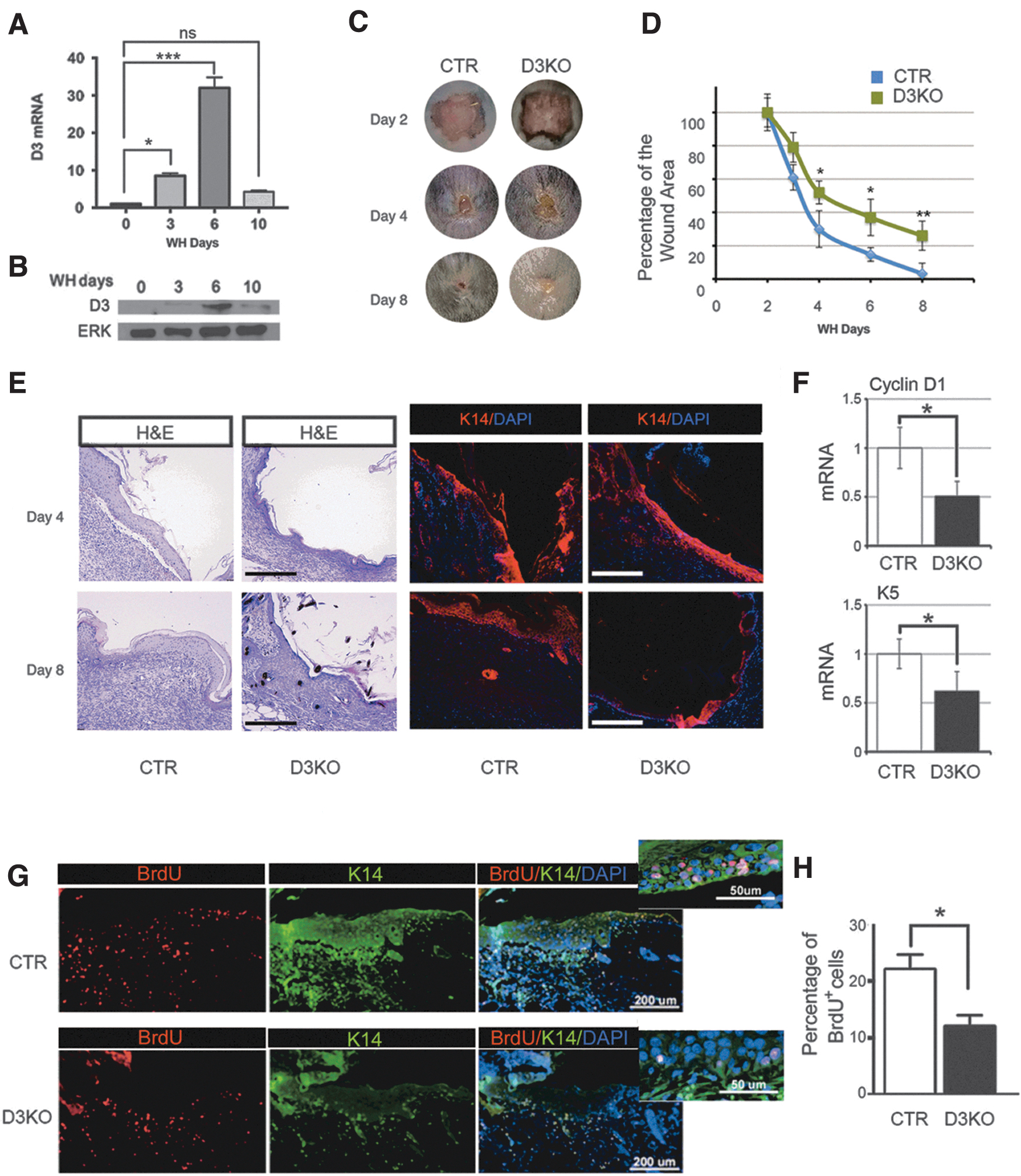
D3KO mice have delayed epidermal regeneration. (
Histological examination confirmed the delay in skin regeneration, as shown by the absence of an epidermal patch in D3KO skin at day 8 postinjury, in contrast to the formation of the normal epidermal layer under the scar in control skin (Fig. 4E and Supplementary Fig. S5). We speculated that this phenotype could be due to a proliferative defect of D3KO keratinocytes. Indeed, mRNA levels of cyclin D1 and K5 were lower in the D3KO epidermis than in control mice (Fig. 4F). Moreover, there were fewer BrdU+ nuclei in the zone surrounding the wound in D3-depleted skin than in control mice (Fig. 4G, H).
Loss of D3 induces premature involution to telogen during hair follicle cycle
Given that D3 expression is upregulated during anagen (Fig. 1D, F), we asked whether the hair follicle cycle is also affected by D3 depletion. Consequently, as shown in Figure 5A, D3 was depleted in 3-month-old mice by tamoxifen treatment; 1 week later, mice were shaved by a clamp and the progression of the hair cycle was assessed 6 (anagen), 10 (catagen), and 30 days (telogen) thereafter. Interestingly, while hair morphology was normal in D3KO mice in anagen, hair bulbs were narrower and hair length was shorter in catagen, which indicates premature involution of the follicle to the telogen phase and altered evolution of the hair follicle (Fig. 5B, C). In line with the premature catagen
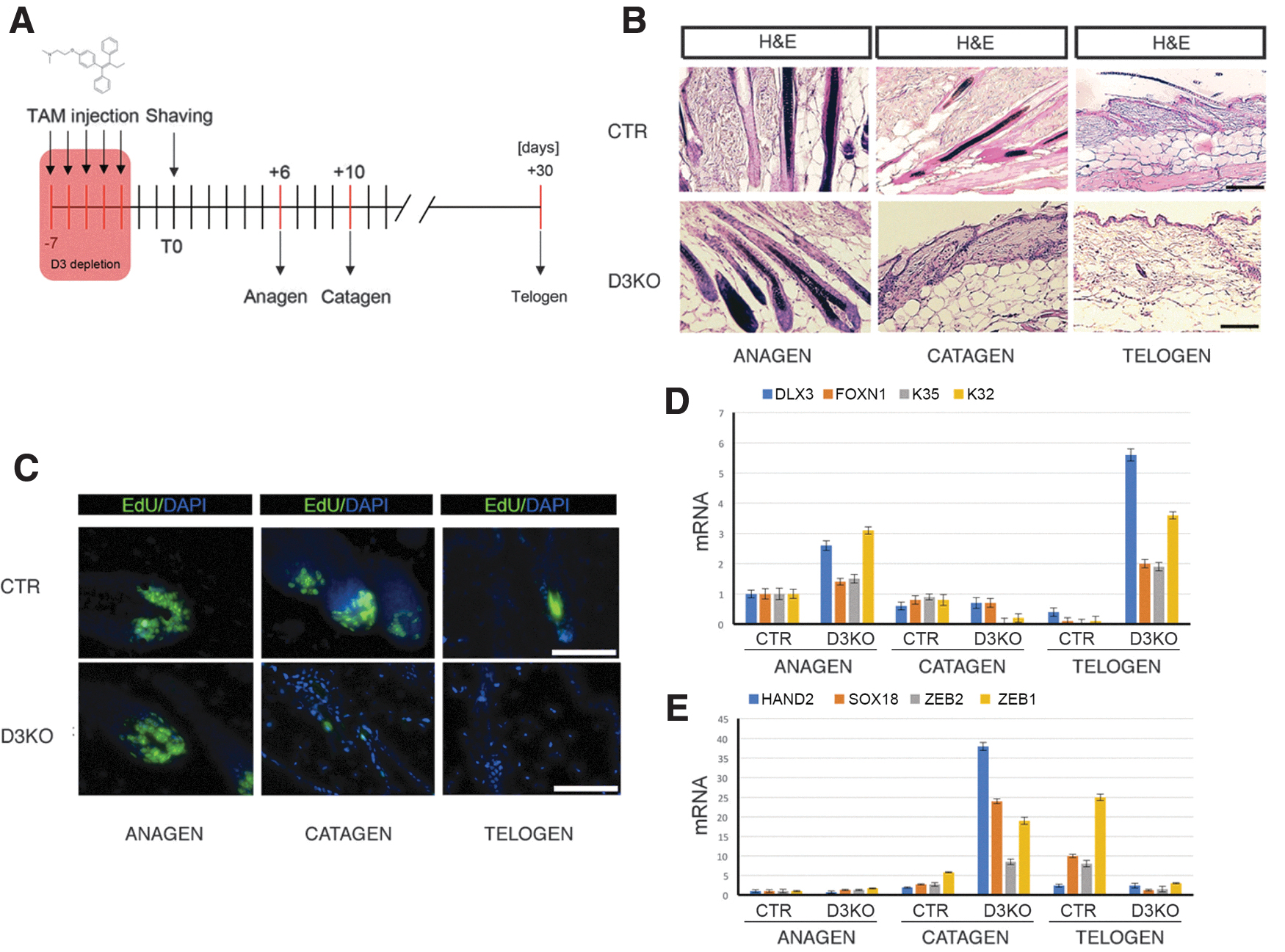
Hair follicle cycle is accelerated in D3KO mice. (
These data are in accordance with a premature differentiation of the hair follicle cycle cells induced by D3 depletion.
Epidermal D3 expression plays a central role in T4 clearance
Since D3 is expressed in adult mice only in brain and skin, and considering that the skin is the largest organ of the body, we wondered whether epidermal D3 might play a role in systemic T4 clearance. To this aim, we measured the levels of D3 in the skin of control mice and mice in which we i.p. injected a supraphysiological dose of T4 (20 μg/100 g) weight for 3 and 7 days. As shown in Figure 6A and B, D3 expression was upregulated in hyperthyroid mice, thereby confirming the homeostatic feedback regulation of TH inactivating enzyme by its substrate. Next, we i.p. injected a supraphysiological dose of T4 and evaluated T4 clearance in D3KO mice and control mice by measuring the systemic levels of T4 in the blood at 24 and 48 hours after injection. Notably, the ability to metabolize T4 was sharply reduced (40% at 24 hours and of about 33% at 48 hours) in D3KO mice versus control mice (Fig. 6C). These results reveal a new role of epidermal D3 in the regulation of plasma TH concentrations.
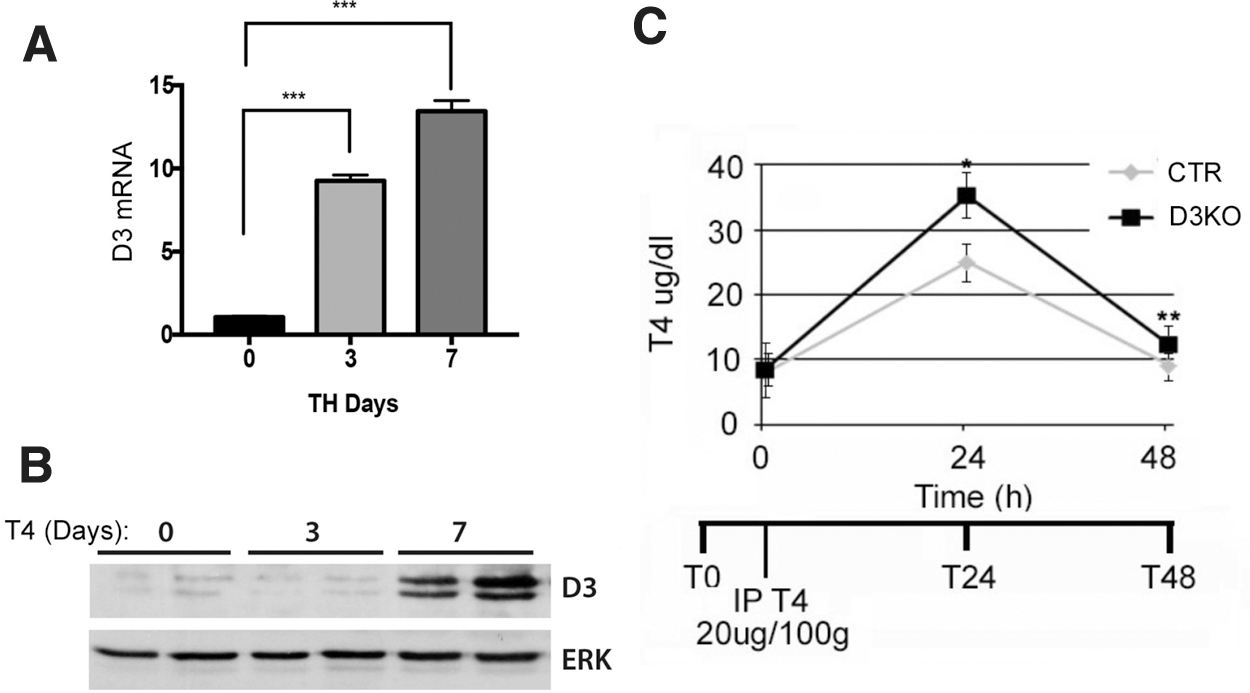
D3KO mice have reduced T4 clearance. (
Discussion
Mouse epidermis is an attractive model system with which to study the control of epithelial cell growth and differentiation (37). TH is important for the development of many tissues, and in adults regulates the balance between proliferation and differentiation of virtually every cell of the body (38,39). TH is a key regulator of epidermal proliferation and differentiation, hair growth, and WH in humans (8) and mice (5,40). A striking example is given by the amphibian metamorphosis, during which TH governs various differentiation programs, including the switch from a bilayered nonkeratinized epithelium to a stratified, keratinized epidermis (41).
Hyperthyroidism in humans leads to alterations in skin homeostasis; increases perspiration and heat dissipation; and can be associated with urticaria, vitiligo, or enhanced pigmentation (42). Notably, the epidermis is thinner in hyperthyroid patients than in euthyroid subjects (42). In contrast, hypothyroidism is associated with a dry, cold, and rough epidermis, with signs of hyperkeratosis, sometime diffuse myxedema, and it can be associated with alopecia (42). Although the effects of thyroid alterations on skin homeostasis have been widely described, the molecular mechanisms by which TH influences keratinocyte turnover and differentiation are still poorly understood. The effects of TH on keratinocyte dynamics are mostly exerted through transcriptional regulation mediated by the genomic action of the two TH nuclear receptors, thyroid hormone receptor alpha and thyroid hormone receptor beta, which act as both positive and negative regulators of transcription on different promoters and in different pathophysiological conditions (10).
The novelty of our study is that it not only analyzes the effects of pathological levels of hyperthyroidism on keratinocyte proliferation/differentiation balance but also addresses the physiological role of the D3 enzyme in the skin. Together with the brain and the placenta, the skin is among the few tissues that expresses D3 (14). What is the reason for such a specific expression profile?
Our study shows that targeted disruption of the Dio3 gene in the epidermis critically affects the basal turnover of keratinocytes in the adult skin. D3 depletion leads to a thinner epidermis, to enhanced expression of stratified layers and a sharp decrease in the stem cell compartment, as well as to a reduction in the basal, proliferative cells. One of the consequences of the altered homeostatic balance between epidermal layers is the delayed regenerative ability of D3-depleted skin after inflicting a wound. In this scenario, massive induction of D3 expression in the early phases of regeneration may well correlate with a requirement of increased cellular proliferation. Thus, these data indicate that D3 is an essential component of the epidermal homeostatic control of keratinocyte turnover, and they suggest that attenuation of TH signaling is critical for the normal physiology of a high-turnover, self-repairing, and self-renewing organ such as the skin. We observed that the systemic levels of THs and TSH were not altered by the epidermal specific D3 depletion (Supplementary Fig. S4). This is in contrast to the observations that plasma T3, T4, and TSH are markedly reduced in global D3 knockout mice due to the loss of D3 during development and the consequent modification of the setting of the hypothalamic pituitary thyroid axis (43). The euthyroid phenotype of the mice with epidermal specific D3KO in our case indicates that loss of D3 after birth is not sufficient to influence the central regulation of thyroid function, although resulting in altered epidermal physiology.
Hair undergoes periods of growth (anagen), regression (catagen), and rest (telogen), thereby generating and maintaining the hair cycling (29). Notably, D3 is highly expressed in anagen, thus coinciding with the highest levels of the Shh pathway that have been shown as a positive regulator of D3 expression (21) and one of the well-known anagen-inducing pathways (44). Since D3 is also expressed in the anagen and catagen phase of the hair follicle cycle, its depletion also alters the normal progression of the hair follicle cycle, and causes premature evolution of the hairs to the telogen phase.
Finally, disruption of the D3-mediated TH signal attenuation also leads to a drastic reduction of the number of epidermal stem cells. In the skin, a population of adult stem cells with self-renewal capacity is located in the basal layer of the epidermis (“interfollicular epidermal stem cells”) and in the bulge of the hair follicle (45). Epidermal stem cells are responsible for the epidermal renewal throughout life (46). Moreover, after an injury, the epidermal stem cells undergo amplification and the differentiation to replenish the mature epidermis in adult skin (46).
D3 expression has been shown to play critical roles in regenerative processes of different organs, such as muscle regeneration (32), liver regeneration (47), and brain repair (48). The finding that D3 depletion reduces the percentage of the epidermal stem cells reinforces the role of D3 in the regenerative capacity of the adult tissues and indicates D3 as an important source for novel therapeutic approaches in the management of wounds.
The results of this study have several important implications: They (1) reinforce previous studies indicating that the intracellular control of TH by deiodinases critically affects the balance between proliferation and differentiation (38); (2) demonstrate that in mouse epidermis, excess of T3 accelerates the differentiation of keratinocytes, induces premature differentiation, thereby resulting in thinner epidermis; (3) indicate that TH is one of the signaling pathways regulating the spatiotemporal control of skin development and turnover; (4) establish D3 as a key determinant of the coordinated transition between anagen-to-catagen in the hair follicle cycle; and (5) provide a new example of the physiological relevance of D3 in the morphogenesis, homeostatic regulation, and regeneration of adult skin.
Footnotes
Acknowledgment
The authors thank Jean Ann Gilder (Scientific Communication srl., Naples, Italy) for writing assistance.
Authors' Contributions
G.M., A.S., C.L., E.D.C., C.M., A.G.C., A.N., S.S., M.A.D.S., D.D.G, T.P., M.M., and F.S. performed in vitro and in vivo experiments and prepared figures; R.A. generated plasmids and mouse models; M.S. performed HPLC-MS analysis of THs; G.P. and P.F. performed analysis of serum TSH; M.D. designed the overall study, supervised the experiments, analyzed the results, and wrote the article; and all authors discussed the results and provided input on the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from AIRC to M.D. (IG 13065), by the ERCStG2014 grant from European Research Council under the European Union's Horizon2020 Programme (STARS—639548), by the EU FP7 contract Thyrage (grant number 666869) to M.D., and by the grant FARE from MIUR (R16KPLYF38) to M.D.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5