
Abstract
Background:
Congenital hypothyroidism due to thyroid dysgenesis is a frequent congenital endocrine disorder for which the molecular mechanisms remain unresolved in the majority of cases. This situation reflects, in part, our still limited knowledge about the mechanisms involved in the early steps of thyroid specification from the endoderm, in particular the extrinsic signaling cues that regulate foregut endoderm patterning. In this study, we used small molecules and genetic zebrafish models to characterize the role of various signaling pathways in thyroid specification.
Methods:
We treated zebrafish embryos during different developmental periods with small-molecule compounds known to manipulate the activity of Wnt signaling pathway and observed effects in thyroid, endoderm, and cardiovascular development using whole-mount in situ hybridization and transgenic fluorescent reporter models. We used the antisense morpholino (MO) technique to create a zebrafish acardiac model. For thyroid rescue experiments, bone morphogenetic protein (BMP) pathway induction in zebrafish embryos was obtained by manipulation of heat-shock inducible transgenic lines.
Results:
Combined analyses of thyroid and cardiovascular development revealed that overactivation of Wnt signaling during early development leads to impaired thyroid specification concurrent with severe defects in the cardiac specification. When using a model of MO-induced blockage of cardiomyocyte differentiation, a similar correlation was observed, suggesting that defective signaling between cardiac mesoderm and endodermal thyroid precursors contributes to thyroid specification impairment. Rescue experiments through transient overactivation of BMP signaling could partially restore thyroid specification in models with defective cardiac development.
Conclusion:
Collectively, our results indicate that BMP signaling is critically required for thyroid cell specification and identify cardiac mesoderm as a likely source of BMP signals.
Introduction
The thyroid is an endoderm-derived gland developing from the most anterior part of the gut tube. Thyroid organogenesis starts with the specification of its anlage, a group of thyroid precursor cells that are characterized by coexpression of a unique combination of transcription factors comprising NKX2–1, PAX8, FOXE1, and HHEX (1).
The median anlage develops into a diverticulum that evaginates and loses contact with the ventral foregut endoderm to relocate deeper into the subpharyngeal mesenchyme. Terminal differentiation, which leads to functional follicle formation, is initiated during the migration of the thyroid primordium (2).
Congenital hypothyroidism (CH) is the most frequent congenital endocrine disorder, affecting approximately one of 2500 in human newborns (3). CH is characterized by reduced serum thyroid hormone levels at birth. It is caused in 85% of the cases by thyroid dysgenesis (TD) due to ectopy, athyreosis, or hypoplasia, resulting from an aberration of the thyroid gland embryological development. The molecular mechanisms leading to TD are mostly unknown, with mutations in the known thyroid transcription factors explaining only 5% of TD cases (1). It probably reflects our limited knowledge about intrinsic factors and external signals coordinating thyroid organogenesis and suggests that unknown genetic and/or epigenetic factors might be crucial for thyroid development (4).
Zebrafish is a valuable model system that has already helped us to improve further our understanding of the morphogenetic processes and gene networks involved in thyroid organogenesis (5). Over the past two decades, zebrafish has gained much attention as a genetically tractable vertebrate model to study organogenesis (6). Embryos' optical clarity allows direct visualization of developmental processes and their pathological deviations. Small size, high fecundity, external fertilization, rapid development, and short generation time are important attributes underscoring the utility of zebrafish study of early developmental processes.
The value of zebrafish for studies on thyroid development is supported by the fact that several aspects of thyroid morphogenesis are well conserved between zebrafish and mammals (7,8). Early morphogenetic events, such as thyroid specification, budding, and relocalization into the subpharyngeal mesenchyme, show many similarities in fish and mouse [reviewed in Porazzi et al. (9)]. Moreover, the developing thyroid expresses a similar, but not identical, set of transcription factors in mouse [NKX2–1, PAX8, HHEX, FOXE1 (1)] and zebrafish [nkx2.4b (10), pax2a, pax8, foxe1, hhex (7,10)] embryos. Ablation of the known thyroid transcription factors in mice leads to athyreosis (NKX2–1, PAX8, FOXE1, HHEX) or thyroid ectopy (FOXE1) (11). Zebrafish with loss-of-function of nkx2.4b, pax2a, and hhex similarly display athyreosis (7,9,12).
However, differences exist in the timing of specific morphogenetic events and the anatomy of the mature thyroid tissue. Indeed, although thyroid follicles are encapsulated in a compact organ in mouse, thyroid follicles are loosely scattered along the pharyngeal midline in zebrafish.
In this study, we mainly focused on molecular mechanisms that govern the first steps of thyroid specification. At present, the signals that trigger the specification of pharyngeal endodermal cells into thyroid precursors remain unknown. Recent studies demonstrate that tissue–tissue interactions, in particular with the cardiac mesoderm and pharyngeal blood vessels, are essential for correct thyroid development (5,13). Defective pharyngeal vessel development has been shown to cause thyroid anomalies in mice and zebrafish (14,15). There are also pieces of evidence coming from human disease studies since an increased prevalence of congenital cardiovascular anomalies is observed in patients with TD compared with the healthy population (16).
However, the mechanisms underlying tissue–tissue interactions during thyroid development (particularly between the developing thyroid and the cardiac mesoderm and pharyngeal vessels) and the signals involved in this cross talk are poorly understood. While a crucial role of fibroblast growth factor (FGF) expression in the mesoderm surrounding the developing thyroid has been demonstrated in mouse and zebrafish (17,18), the potential functions of other major signaling pathways in thyroid organogenesis are still to be clarified (2,19). A significant advantage of the zebrafish model in cardiovascular development studies is that zebrafish embryos and larvae can survive for several days without a functional heart or in the absence of blood circulation (20). This facilitates the characterization of developmental effects over an extended developmental period, compared with most mouse models with cardiovascular defects.
To improve our understanding of the role of extrinsic signaling cues in thyroid development, we recently exploited the amenability of zebrafish embryos for small-molecule screenings to identify candidate signaling pathways (21). From a literature review, we first identified a collection of small molecules known to interfere with common signaling pathways in zebrafish. The pharmacological screening readily identified modulators of Wnt, bone morphogenetic protein (BMP), FGF, and transforming growth factor-β signaling to cause distinct effect patterns of disturbed thyroid organogenesis (e.g., agenesis, hypoplasia, ectopy) (21). In this article, we focused on the phenotypic description, using thyroid and cardiovascular markers, of early thyroid development after Wnt modulation and propose a mechanistic explanation.
Wnt signaling pathway is an evolutionarily conserved signal transduction pathway that regulates crucial aspects of cell fate determination, cell migration, cell polarity, and cell differentiation during embryonic development (22). Wnt proteins are secreted glycoproteins that bind to the N-terminal extracellular cysteine-rich domain of the frizzled (Fz) receptor family, interacting with the LRP5/6 coreceptors.
To date, several intracellular signaling cascades downstream of the Fz receptors have been identified, including a canonical (β-catenin-dependent) pathway and a noncanonical (β-catenin-independent) pathway. Some Wnt ligands act preferentially on one pathway or the other, but the activated pathway mostly depends on the cellular context. Without Wnt canonical signaling activation, cytoplasmic β-catenin is degraded by a β-catenin destruction complex, which includes Axin, adenomatous polyposis coli, protein phosphatase 2A, glycogen synthase kinase 3 (GSK3), and casein kinase 1α. Binding of a Wnt ligand to its receptor leads to a series of events that disrupt the β-catenin destruction complex, which is required for the β-catenin translocation into the nucleus and activation of target genes (23,24).
The functions of Wnt/β-catenin signaling in embryogenesis have been extensively studied in different animal models. It plays a crucial biphasic role in heart development, as demonstrated in in vitro cells and zebrafish: in pregastrula stages, canonical Wnt promotes the specification of the precardiac mesoderm into cardiomyocyte progenitors, but it inhibits the cardiac differentiation of these cells during gastrula stages (25,26). It has already been demonstrated that this pathway also plays a very important role in the specification and development of endoderm-derived organs such as the liver and pancreas. An anterior–posterior gradient of Wnt activity in Xenopus plays a crucial role in endodermal patterning, the anterior endoderm giving rise among others to the liver, lung, and thyroid and the posterior endoderm leading to intestinal fate (27). At later stages in zebrafish, Poulain and Ober have shown that Wnt pathway is necessary for liver progenitor proliferation (28). However, very little data exist on the role of Wnt in thyroid development and maintenance.
Therefore, this study aims to examine the effects of the canonical Wnt signaling pathway on thyroid development in zebrafish, in regard to the adjacent cardiogenesis. We used different chemicals and genetic models to modulate canonical Wnt signaling during gastrula and early somitogenesis stages and observed effects of such modulation on thyroid organogenesis. We succeeded to partially rescue thyroid defects obtained after Wnt activation when combining this chemical treatment with induction of BMP pathway using heat-shock inducible transgenic lines.
Materials and Methods
Zebrafish husbandry and embryo culture
Zebrafish (Danio rerio) embryos were obtained from natural spawning of adult fish, raised at 28.5°C according to Westerfield (29) and staged in hours postfertilization (hpf) as described by Kimmel et al. (30) In addition to wild-type and casper mutant lines (31), different transgenic lines were used in this study: Tg(tg:mCherry) (5), Tg(kdrl:EGFP) (32), Tg(myl7:EGFP) (33), Tg(7xTCF-Xla.Siam:GFP)ia (34), Tg(sox17:EGFP) (35), Tg(hsp70l:bmp2b) (36), and Tg(hsp70l:wnt8a-EGFP) (37). Embryos were dechorionated at 24 hpf using 0.6 mg/mL pronase (Sigma, Saint-Louis, MO), anesthetized in 0.016% tricaine (Sigma), fixed in 4% phosphate-buffered paraformaldehyde (PFA; Sigma) overnight at 4°C, washed in phosphate-buffered saline (PBS) containing 0.1% Tween 20 (PBST), gradually transferred to 100% methanol, and stored at −20°C until used for in situ hybridization or immunofluorescence analyses. If indicated, pigmentation of embryos was inhibited by adding 0.003% 1-phenyl-2-thiourea (Sigma) (38) to the embryo medium at 24 hpf. All zebrafish work at the Institute of Interdisciplinary Research in Molecular Human Biology followed protocols approved by the Institutional Animal Care and Use Committee.
Small-molecule treatment
We used timed embryo treatment with BIO (Sigma; B1686), 1-azakenpaullone (AZA, A3734; Sigma), and IWR-1 (Sigma; I0161) to modulate the canonical Wnt signaling pathway during distinct developmental periods. BIO and AZA act as canonical Wnt activators due to GSK3β inhibition (39). In contrast, IWR-1 stabilizes the destruction complex, thereby acting as an inhibitor of Wnt signaling (40). Stock solutions of BIO (5 mM), AZA (5 mM), and IWR-1 (10 mM) were prepared in dimethyl sulfoxide (DMSO). Test solutions were prepared by diluting the stock solutions in embryo medium. A control treatment containing 0.1% DMSO was used in all experiments involving a drug treatment.
Heat-shock treatments
Timed global overactivation of Wnt and BMP signaling was induced by heat-shock treatment of the progeny from matings of wild-type fish with heterozygous Tg(hsp70l:wnt8a-GFP) (37) or Tg(hsp70l:bmp2b) (36) fish, respectively. Embryos obtained from these matings were raised under standard conditions until heat-shock to induce global overexpression of enhanced green fluorescent protein (EGFP)-tagged Wnt8a or Bmp2b. For the heat-shock treatments, embryos were transferred to dishes containing prewarmed embryo medium at 40°C and incubated in an incubator for 30 min at 40°C. After heat-shock treatment, embryos were transferred to Petri dishes containing fresh medium and allowed to develop further at 28.5°C under standard conditions. Carriers of the hsp70l:wnt8a-EGFP transgene were identified three hours after heat-shock by means of their EGFP expression. Embryos carrying the hsp70l:bmp2b transgene were identified by their dorsalized phenotype if heat-shocked at early somitogenesis or by polymerase chain reaction (PCR) genotyping if embryos were heat-shocked at later stages (15 or 20 hpf). For the latter, PCR genotyping was performed after completion of whole-mount in situ hybridization (WISH) experiments as previously described (41) using the following primers: forward 5′-CATGTGGACTGCCTATGTTCATC-3′ (primer located in hsp70l promoter sequence); and reverse 5′-GAGAGCGCGGACCACGGCGAC-3′ (primer located in bmp2b coding sequence).
Whole-mount in situ hybridization
DNA templates for synthesis of nkx2.4b, tg, nkx2.5, gfp, mef2cb, gata4, gata5, foxa2, foxa3, hhex, pdx1, prox1, and bmp4 riboprobes were generated by PCR (see Supplementary Table S1 for primer sequences). Plasmids for amhc, vmhc, myl7, scl, and hand2 riboprobes have been used as described (42 –44). Single-color WISH using digoxigenin (DIG)-labeled riboprobes and anti-DIG antibody conjugated to alkaline phosphatase was performed essentially as described in Thisse and Thisse (45) and Opitz et al. (46). Riboprobe hybridization was performed at 65°C overnight, and probes were detected using an anti-DIG antibody (1:6000; Roche, Bâle, Switzerland). Staining reactions were performed with the alkaline phosphatase substrates BM Purple (Roche) or nitro-blue tetrazolium (NBT)/5-bromo-4-chloro-3′-indolyphosphate (BCIP) (Roche). For dual-color WISH, riboprobes labeled with DIG, dinitrophenol (DNP), or fluorescein (FLU) were used, and sequential alkaline phosphatase staining was performed with BM Purple and Fast Red (Sigma) as described (21). We first detect the DIG or DNP riboprobes with anti-DIG or anti-DNP antibody using BM Purple (for most genes) or NBT/BCIP (for tg) staining solution and used an anti-FLU antibody in combination with FastRed for detection of the FLU probe in a second step. Removal of the antibodies after the first staining step was performed by 2 × 5 minutes of incubation in 100 mM glycine-HCl (pH 2.2).
Fluorescent WISH (FISH) using a DIG-labeled riboprobe for nkx2.4b was performed as described (5). Antibodies used in WISH and FISH experiments are listed in Supplementary Table S2. Stained embryos were washed in PBST, postfixed in 4% PFA, and embedded in 90% glycerol for whole-mount imaging or in 7% low melting point agarose (Lonza) for vibratome sectioning. Whole-mount images of WISH and FISH were acquired using an MZ16F Leica stereomicroscope equipped with a DFC420C camera or a Leica microscope DMI6000B equipped with a DFC365FX camera, respectively. Vibratome tissue sections at 50–60 μm thickness were cut on a Leica VT1000S vibratome and mounted in Glycergel (Dako). Confocal images of vibratome sections were acquired using an LSM510 confocal microscope (Zeiss).
Whole-mount immunofluorescence
Whole-mount immunofluorescence (WIF) was performed essentially as described (46). Briefly, after rehydration into PBST, embryos were immersed in blocking buffer (PBST containing 1.0% DMSO, 1% bovine serum albumin [BSA], 5% horse serum, and 0.8% Triton X-100) for two hours. Embryos were then incubated overnight in blocking buffer containing primary antibodies at 4°C. After several washing steps in PBST containing 1% BSA, embryos were incubated with secondary antibodies overnight at 4°C. Specifications and sources of primary and secondary antibodies used to detect EGFP and pSMAD1/5/8 (47) protein in zebrafish embryos are provided in Supplementary Table S2. Stained embryos were washed in PBST and embedded in 90% glycerol for fluorescence microscopy. Images were acquired with a Leica DFC365FX camera mounted on a Leica DMI6000B. Combined FISH and WIF staining was performed as described (5). Confocal images were acquired using an LSM510 confocal microscope (Zeiss).
RNA extraction and reverse transcription quantitative polymerase chain reaction
For total RNA preparation, pools of 10–20 embryos per sample were lysed in RNeasy lysis buffer (Qiagen, Hilden, Germany) containing 1% 2-mercaptoethanol (Sigma). Total RNA was isolated using the RNeasy RNA preparation microkit (Qiagen) according to the manufacturer's instructions, including on-column treatment with DNase I (Qiagen). Reverse transcription was done using Superscript II kit (Invitrogen). Reverse transcription quantitative PCR (RT-qPCR) was performed in duplicate using Kapa SYBR Fast (Kapa Biosystems, Wilmington, MA) mix and a CFX Connect Real-Time PCR System (Bio-Rad, Hercules, CA).
Relative values for target transcript abundance in individual samples were determined by the comparative CT method (ΔΔCT) according to Pfaffl (48), and data are presented as relative expression values normalized to the reference gene rpl13 (NM_212784.1) whose expression remained constant upon treatment. Primers used were as follows: rpl13, forward TCTGGAGGACTGTAAGAGGTATGC, reverse AGACGCACAATCTTGAGAGCAG (49); gfp, forward AGAACGGCATCAAGGTGAAC, reverse TGCTCAGGTAGTGGTTGTCG. Gene expression profile was confirmed in triplicate (three different batches of embryos). Data are expressed as mean ± standard deviation. Pairwise comparisons were performed using the Student t-test.
Fluorescence-activated cell sorting
Cell suspensions were prepared from Tg(sox17:EGFP) transgenic embryos at 30 hpf. Embryos were digested in HBSS (Gibco, Waltham, MA) containing 0.25% trypsin (Gibco) and 2 mM ethylenediaminetetraaceticacid (EDTA; Invitrogen). Single-cell solutions were obtained by constant pipetting. Reaction was stopped by adding CaCl2 (final concentration: 1 mM) and fetal bovine serum (final concentration: 10%). Cells were pelleted by centrifugation and washed with PBS containing 0.4% BSA and 2 mM EDTA. Finally, resuspended cells were passed through a 40 μM nylon mesh (Falcon, Corning, NY) into a fluorescence-activated cell sorting (FACS) tube (Falcon) and the GFP+ fractions were analyzed by a fluorescent-activated cell sorter (FACS Aria: FACSDiva Software; BD, Franklin Lakes, NJ). About 50 embryos were sampled per condition, and each experiment was performed in triplicate.
Morpholino microinjection
For inhibition of the mef2ca and mef2cb transcripts, zebrafish embryos were injected with morpholino (MO) antisense oligonucleotides that have previously been validated for their knockdown specificity and efficacy (50,51). We injected 4 nL (2 ng/nL) of a translation-blocking mef2d/c-MO (tb-MO; 5′-ATGGGGAGGAAAAAGATCCAGATTC-3′) as previously described (50). Working solutions of MOs were prepared in 0.12 M KCl containing phenol red, and 4 nL was microinjected into the yolk of one- to two-cell stage embryos. Noninjected embryos served as control embryos.
Results
Drug-induced manipulation of canonical Wnt signaling disrupts early thyroid development
We recently performed a small-molecule screen to identify signaling pathways involved in early zebrafish thyroid development (21). In these experiments, pharmacological manipulations of canonical Wnt signaling during the gastrula stage resulted in abnormal thyroid marker expression, and, most notably, in a severe reduction of nkx2.4b expression at thyroid anlage stages (28 hpf) following overactivation of Wnt signaling.
To characterize in more detail the thyroidal effects resulting from enhanced Wnt signaling, we performed additional treatment experiments with BIO and AZA, two drugs known to activate Wnt/β-catenin signaling (39). Consistent with our previous results, treatment of zebrafish embryos with BIO or AZA during the gastrulation period (6–10 hpf) caused concentration-dependent decreases of nkx2.4b expression in 28 hpf embryos (Fig. 1). Maximal effects on thyroidal nkx2.4b expression (i.e., complete absence of a detectable WISH staining) were evident with the concentration of 5 μM BIO or 5 μM AZA.
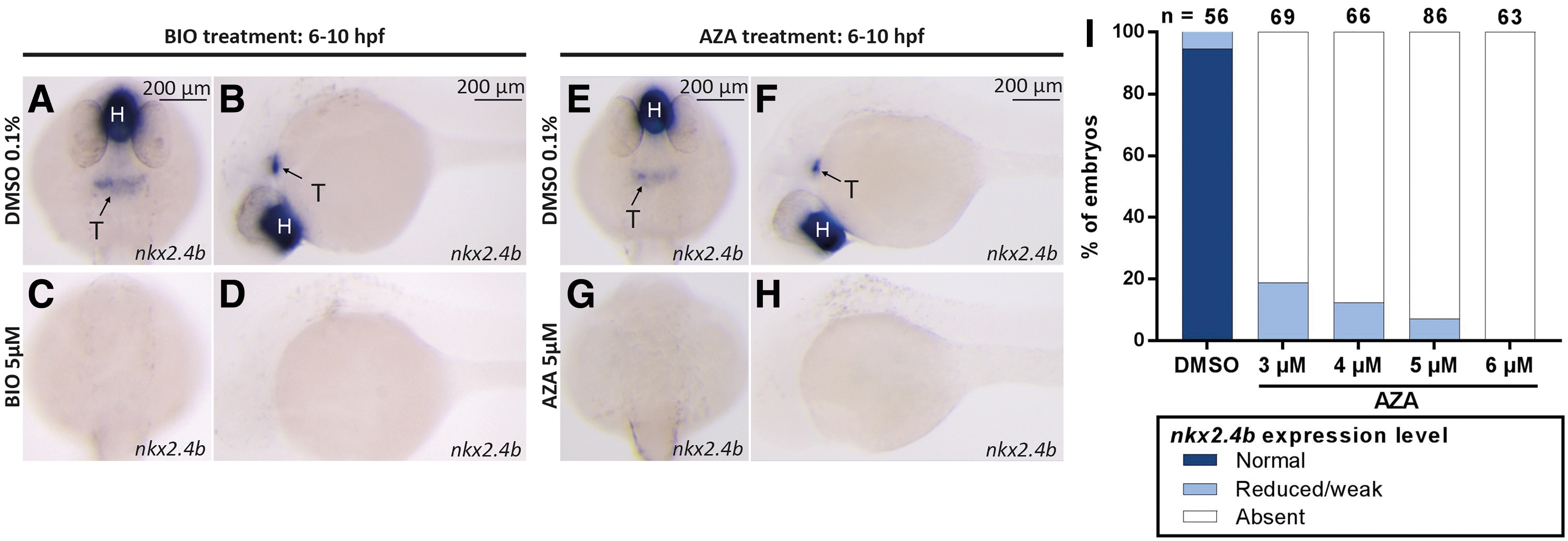
Drug-induced overactivation of canonical Wnt signaling during gastrulation impairs thyroid specification. (
In addition to the marked effects on the thyroidal nkx2.4b expression domain, BIO and AZA treatments also caused concentration-dependent decreases in the size of the nkx2.4b expression domain in the forebrain. The drug-induced effects on forebrain nkx2.4b expression correlated closely with other visible signs of global posteriorization, including loss of anterior neural tissue and reduced size or absence of the eyes (52,53). Although we did not quantify the relative size reductions of thyroidal and forebrain nkx2.4b expression domains in 28 hpf embryos following different BIO and AZA treatments, we noted a similar sensitivity to drug treatment for both expression domains when embryos were treated between 6 and 10 hpf.
Concentration-dependent effects on thyroid development were also evident when BIO-treated embryos were analyzed for thyroid marker expression at 55 hpf (Fig. 2). WISH analysis of nkx2.4b expression revealed concurrent reductions in the size of thyroidal and forebrain expression domains in treated embryos. Moreover, BIO treatment strongly reduced the expression of the functional thyroid differentiation marker, thyroglobulin (tg), in 55 hpf embryos. However, we also noted that the majority of 55 hpf embryos treated with high drug concentrations during gastrula stages displayed at least some residual staining for nkx2.4b and tg despite undetectable thyroid marker expression at the thyroid anlage stage (28 hpf). This was observed in all experiments involving drug treatment from 6 to 10 hpf.
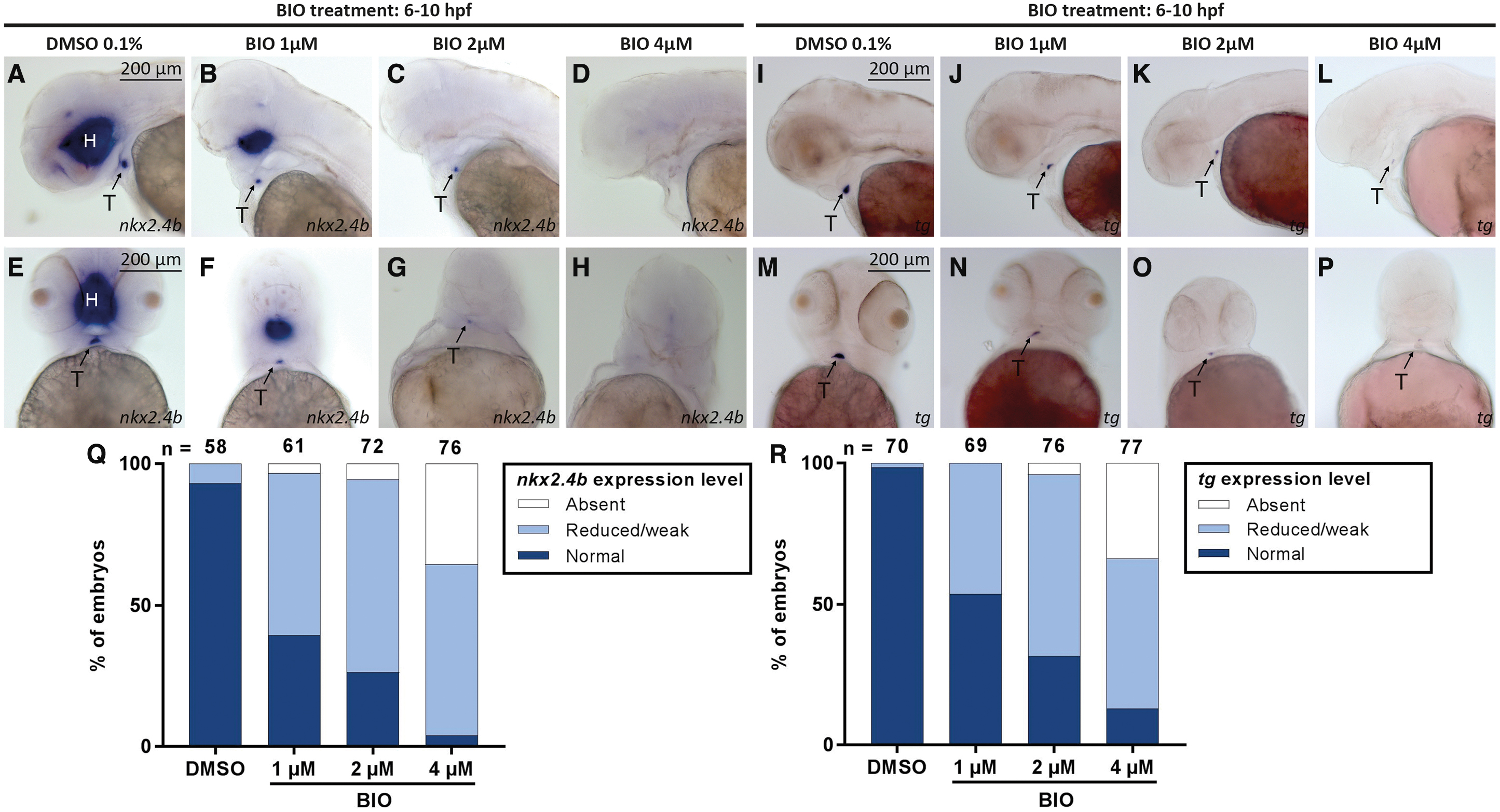
Drug-induced overactivation of canonical Wnt signaling during gastrulation impairs the thyroid primordium formation. (
Conversely, treatment of gastrulating embryos with 10 μM IWR-1, a small-molecule inhibitor of canonical Wnt signaling, resulted in additional ectopic domains of thyroidal nkx2.4b and tg expression (Supplementary Fig. S1). In 28 hpf embryos, the additional ectopic nkx2.4b expression was detected posterior to the orthotopic thyroidal nkx2.4b expression domain, whereas in 55 hpf embryos, supernumerary clusters of tg expression were detectable at irregular posterior positions. Taken together, these data indicate that the canonical Wnt pathway negatively regulates thyroid specification during the gastrulation stage in zebrafish as drug-mediated activation or inhibition of Wnt activity during gastrulation impairs or amplifies thyroid specification, respectively.
Small-molecule compounds rapidly act on Wnt signaling
We next took advantage of an available Wnt signaling biosensor line, Tg(7xTCF-Xla.Siam:GFP)ia4 , to verify that the small-molecule compounds BIO, AZA, and IWR-1 effectively alter canonical Wnt signaling activities within a short space of time under the experimental conditions that resulted in irregular thyroid development. For this purpose, we treated Tg(7xTCF-Xla.Siam:GFP)ia4 embryos with BIO (5 μM), AZA (5 μM), or IWR-1 (10 μM) from 6 to 10 hpf and monitored the expression of GFP mRNA during the course of drug treatment by WISH and RT-qPCR (Supplementary Fig. S2).
WISH analyses of GFP mRNA expression in BIO- and AZA-treated embryos showed a rapid and robust upregulation of GFP mRNA expression when compared with DMSO-treated embryos (0.1% DMSO) indicating effective overactivation of canonical Wnt signaling in drugged embryos within two hours after the beginning of drug treatment. Consistently, IWR-1 treatment resulted in reduced GFP mRNA expression, evident within 2–3 hours after the beginning of drug treatment. Although several embryos treated with these small-molecule compounds experienced slight developmental delay, the drug-induced alterations in reporter expression were robustly detectable in comparison with vehicle control embryos.
In further experiments, we also corroborated the effects of BIO treatment on GFP mRNA expression by RT-qPCR. As shown in Supplementary Figure S2, treatment of Tg(7xTCF-Xla.Siam:GFP)ia4 embryos with 5 μM BIO resulted in 2.5- to 3-fold increases of whole-embryo expression levels of GFP mRNA within two hours after initiation of BIO treatment. Together, these data demonstrate that, under our experimental conditions, the drug treatments faithfully altered canonical Wnt signaling.
To provide an independent line of evidence that defective thyroid development is due to transient overactivation of Wnt/β-catenin signaling during gastrulation, we manipulated Wnt/β-catenin signaling by heat-shock-induced overexpression of wnt8a in Tg(hsp70l:wnt8a-GFP) embryos (37). Wnt8a specifically activates Wnt/β-catenin signaling in zebrafish embryos and a 30-minute heat-shock of transgenic Tg(hsp70l:wnt8a-GFP) embryos at 6 hpf resulted in a marked neural posteriorization phenotype and disturbed early thyroid development (Supplementary Fig. S3).
Nontransgenic embryos showed no discernible phenotype in response to the heat-shock treatment. The thyroid phenotype caused by transient wnt8a overexpression in the genetic model closely resembled the phenotype observed following treatment with 4–5 μM BIO or AZA. Specifically, our WISH analyses failed to detect nkx2.4b expression in the thyroid anlage region of heat-shocked embryos carrying the wnt8a-GFP transgene at 28 hpf and a detectable, although strongly diminished, tg expression in 55 hpf transgenic embryos (Supplementary Fig. S3). Taken together, these experiments verified that the selected drug concentrations cause overactivation of Wnt/β-catenin signaling and that enhanced Wnt/β-catenin signaling during gastrula stages leads to defects in thyroid development.
Effects of early Wnt overactivation on endoderm development
Our initial experiments with BIO and AZA showed that treatment with different drug concentrations during gastrulation causes thyroid abnormalities and a strong posteriorization of neural ectoderm. Considering that the latter phenotype is a well-characterized effect of enhanced Wnt signaling during early development (54), we next examined whether endodermal development is similarly affected by a global posteriorization activity due to enhanced Wnt signaling and might thus explain the loss of thyroid marker expression in the anterior endoderm.
To address this question, we first treated embryos of the transgenic Tg(sox17:EGFP) line with BIO (5 μM) or AZA (5 μM) between 6 and 10 hpf and analyzed the gross morphology of their foregut endoderm at different embryonic stages, in comparison with that of DMSO-treated embryos. While a neural posteriorization phenotype (loss of anterior neural tissue) was readily detectable in BIO- and AZA-treated Tg(sox17:EGFP) embryos, the gross morphology of the anterior endoderm appeared unaffected in these embryos (Supplementary Fig. S4). In addition, we used FACS to compare the number of GFP+ cells in 30 hpf Tg(sox17:EGFP) embryos following treatment with DMSO (0.1%) and BIO (5 μM), but did not detect a decrease in the number of GFP+ cells in BIO-treated embryos (Supplementary Fig. S4). Thus, in contrast to the overt effects on anterior neural tissue development, no visible dysgenesis of the anterior endoderm was detected in Tg(sox17:EGFP) following early Wnt overactivation.
To further study possible global endodermal patterning defects in BIO- and AZA-treated embryos, we analyzed the expression of a panel of informative endodermal marker genes by WISH (Supplementary Fig. S5). We first confirmed that thyroidal expression of nkx2.4b and hhex is affected similarly by BIO treatment (Supplementary Fig. S5A–D) and AZA treatment (data not shown). In addition to the thyroid, hhex is expressed in the developing liver and pancreas. Interestingly, hhex expression was selectively repressed in the thyroid anlage of BIO-treated embryos and was maintained in the liver/pancreas region (Supplementary Fig. S5D). Notably, the shape and position of the hepatic and pancreatic hhex expression domains were altered in BIO-treated embryos.
Therefore, we analyzed the expression of foxa2, a master regulator of foregut endoderm patterning. Notably, our WISH analyses did not reveal gross changes in the foxa2 anterior–posterior expression pattern following BIO treatment (Supplementary Fig. S5E, F). However, when analyzing foxa3 expression, a key regulator of posterior endoderm development, we noted that its expression domain appeared extended more anteriorly in many embryos after BIO treatment (Supplementary Fig. S5G, H). Analyses of pdx1 and prox1a expression confirmed that hepatic and pancreatic markers are not repressed in BIO-treated embryos and that diminished expression in response to canonical Wnt overactivation is limited to thyroidal marker genes (Supplementary Fig. S5I–L).
Collectively, our analyses indicate that the loss of thyroid marker expression caused by overactivation of canonical Wnt signaling during gastrulation is not due to a general defect in anterior foregut formation. However, as indicated by the irregular anterior foxa3 expression in BIO-treated embryos and the posterior extension of thyroid marker expression in IWR-1-treated embryos, we cannot rule out that a Wnt-induced foregut endoderm posteriorization might contribute to the thyroid defects seen in the most anterior part of the foregut.
Cardiac phenotype is correlated with thyroid phenotype after gastrula Wnt activation
A recurrent theme in the development of endoderm-derived organs is the critical role of tissue–tissue interactions, particularly between the endoderm and adjacent mesodermal tissues (28,55). Observations made during our initial small-molecule screening experiments (21) indicated that overactive Wnt signaling might induce the concurrent cardiac and thyroidal maldevelopment. To examine this possible relationship in more detail, we first confirmed that drug-induced overactivation of Wnt signaling during gastrula stages results in a severe inhibition of heart formation (Supplementary Fig. S6), as previously reported for genetic models with overactivation of canonical Wnt signaling (25,26). When treated between 6 and 10 hpf, effects of Wnt-activating drugs on heart development appeared concentration-dependent, and we noticed that development of ventricular cardiomyocytes was more severely affected, compared with that of atrial cardiomyocytes (Supplementary Fig. S6). Notably, the concentrations at which BIO and AZA caused severe cardiac maldevelopment (Supplementary Fig. S7) overlapped the range at which repression of thyroid markers was detected.
To characterize the evolution of the cardiac differentiation defects in drugged embryos, we examined the expression of various cardiac differentiation markers at earlier stages of cardiac development (Supplementary Fig. S7). Comparative WISH analyses of BIO- and DMSO-treated embryos showed severely reduced expression of transcription factors regulating cardiovascular development such as nkx2.5, mef2cb, and scl in response to Wnt overactivation (Supplementary Fig. S7A–F).
Consistent with previous studies using genetic zebrafish models (26), expression levels of these genes were dramatically decreased as early as 13 hpf in BIO-treated embryos. Conversely, we observed only mild changes, if any, in the expression of transcription factors with a broader lateral plate mesoderm expression domain, including gata4, gata5, and hand2 (Supplementary Fig. S7G–L). A concentration-dependent effect of BIO treatment on the myocardial marker myl7 expression was evident at 19 hpf, when bilateral cardiac fields are supposed to fuse and form the cardiac cone (Supplementary Fig. S7M–P). Given that endodermal progenitors of the thyroid lineage develop in close vicinity to the anterior lateral plate mesoderm containing cardiac precursors in zebrafish (17), we hypothesized that diminished availability of cardiac mesoderm-borne signaling cues contributes to the defective thyroid development in BIO-treated embryos.
To evaluate a possible causal relationship between inhibited cardiac development and TD, we examined the co-occurrence of cardiac and thyroid developmental defects over a range of drug concentrations by dual-color WISH of cardiac (myl7) and thyroid (nkx2.4b) marker expression in 28 hpf embryos (Fig. 3). For embryos treated with increasing concentrations of BIO from 6 to 10 hpf, we observed a gradual loss of cardiomyocytes correlated with a gradual reduction in thyroid marker expression (Fig. 3A–F). Notably, although we observed embryos without any detectable thyroidal nkx2.4b expression but still displaying remnant myl7 positive cardiac tissue, we never observed any embryo with detectable thyroid marker expression in the complete absence of myl7 cardiac expression.
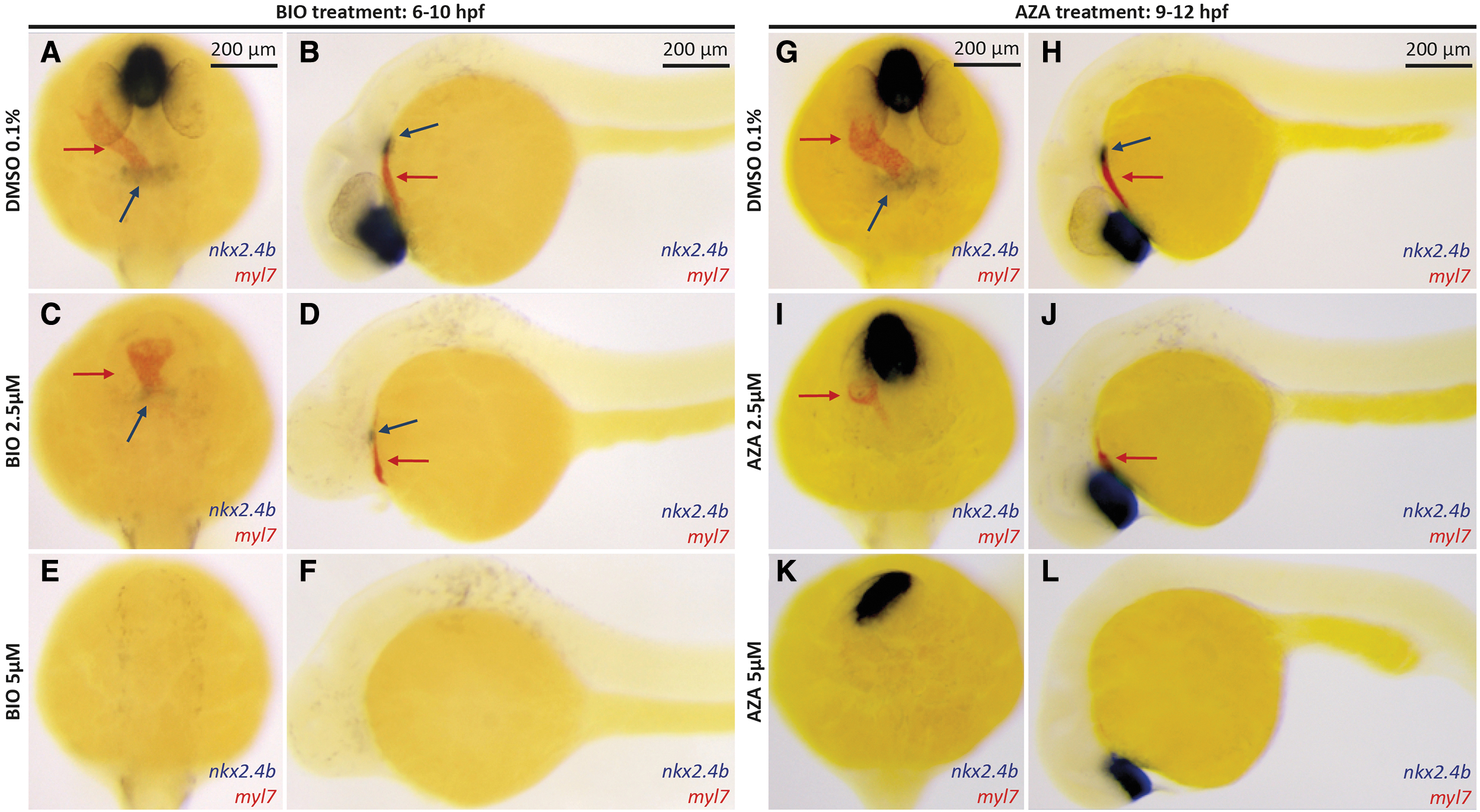
Thyroid specification defects coincide with impaired cardiac development in embryos following drug-induced overactivation of canonical Wnt signaling. (
While these observations are compatible with the hypothesis that the availability of cardiac mesoderm and associated signaling cues are prerequisites for thyroid lineage specification to occur, the concurrent global posteriorization phenotype of the embryos prevented us from distinguishing the relative impact of cardiac dysgenesis against a possible posteriorized patterning in the etiology of thyroid specification defect.
It has been demonstrated that the later the embryos are treated with Wnt activators during gastrulation, the weaker is the posteriorization effect. Still, cardiac development remains highly sensitive to these later treatments (26). Therefore, we progressively shifted the treatment periods toward later stages and assessed the impact of drug-induced Wnt signaling on global posterization and cardiac and thyroid development in these embryos. As expected, gross morphological signs of posterization were greatly reduced if embryos were treated from 9 to 12 hpf (Fig. 3G–L). However, Wnt overactivation at these later stages still caused a severe inhibition of cardiac development. Moreover, it appeared that the prevalence and severity of cardiac dysgenesis and thyroid misspecification correlated closely in embryos treated with Wnt-activating drugs between 9 and 12 hpf. We made the same observations when we treated the embryos from 10 to 13 hpf or from 9 to 24 hpf (Supplementary Fig. S8E–H, K, and L). However, we noted that initiation of drug treatment at 11 hpf or later caused progressively milder effects on both cardiac and thyroid differentiations as assessed by WISH for cardiac myl7 expression and thyroidal nkx2.4b expression (Supplementary Fig. S8I, J).
Based on this experiment, the window of treatment that causes the most striking effect on thyroid development stands during the end of gastrulation and the beginning of somitogenesis. Moreover, we observed a stronger impact on thyroid and cardiac development when AZA treatment was applied from 9 to 24 hpf than from 9 to 12 hpf. This was not surprising since it has been demonstrated that cardiomyocyte progenitor differentiation requires Wnt signaling during midsomitogenesis, at least until 13 hpf (eight somites) (26).
Taken together, these results show a correlation between cardiac and thyroid specification defects, despite a minimal global posteriorization effect when chemical treatment is shifted at the end of gastrula and/or at the early beginning of somitogenesis.
Cardiomyocyte depletion is associated with TD
To further examine the hypothesis that depletion of cardiac mesoderm results in a failure of thyroid cell specification, we sought for cardiac differentiation-defective zebrafish models. To the best of our knowledge, a zebrafish model completely lacking cardiac cells while maintaining a normal anterior endoderm has not yet been described. Hinits and Hughes and Hinits et al. recently described a severe deficiency in cardiomyocyte differentiation in a dual loss-of-function model for zebrafish cardiac transcription factors mef2ca and mef2cb (50,51). For our study, we used an antisense MO (mef2c/d-MO) developed by the same group that reportedly ablates several Mef2 proteins and faithfully replicates the cardiac phenotype present in mef2ca/mef2cb double mutant fish (51).
In our hands, injection of the mef2c/d-MO into wild-type embryos, hereafter called Mef2c-deficient embryos, robustly inhibited cardiomyocyte development as evident from only faint myl7 staining of small and very thin heart tube remnants at 28 and 55 hpf (Fig. 4). Compared with control embryos, we also observed a greatly reduced expression of nkx2.4b in the thyroid anlage of 28 hpf mef2c/d-morphant embryos (Fig. 4A–D). Across several mef2c/d-MO injection experiments, we observed that the prevalence and severity of cardiac malformation correlated with the extent of impaired thyroid marker expression in 28 hpf morphant embryos. Detectable domains of nkx2.4b expression displayed a weaker staining intensity and were of a smaller size in Mef2c-deficient embryos relative to control embryos. Comparison of control and Mef2c-deficient embryos at 55 hpf revealed an irregular organization of tg-expressing cells in Mef2c-deficient embryos, which displayed a fainter tg staining of individual cells (Fig. 4E–H). Collectively, even if we could not acquire completely acardiac embryos with this technique, observations made in the Mef2c-deficient zebrafish model of impaired cardiomyocyte differentiation support the contention that thyroid anlage specification relies on normal cardiac development.
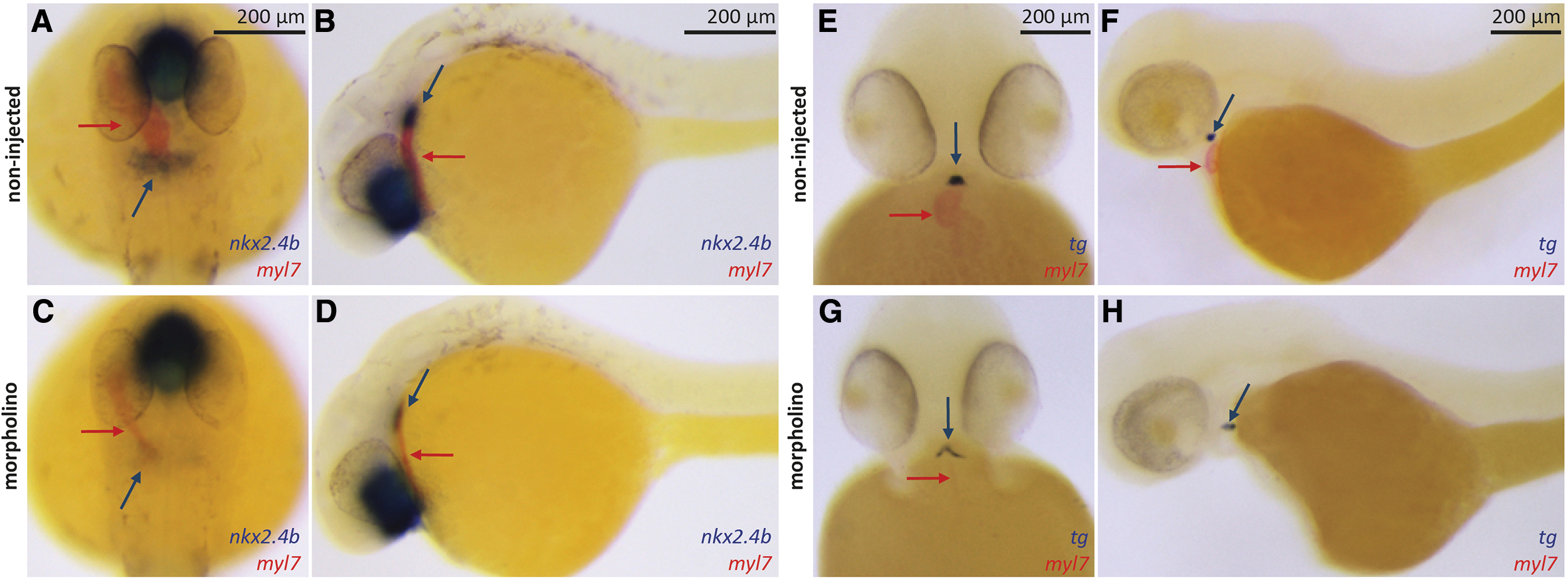
Impaired cardiac development in mef2c/d MO-injected embryos is associated with reduced thyroid marker expression. (
Enhancing BMP signaling causes partial rescue of thyroid defects in BIO-treated embryos
Cardiac mesoderm expresses a number of diffusible growth factors, including several FGF and BMP ligands (17,51) with potential thyroid specification-inducing capacity (17,56,57). In this respect, our recent small-molecule screening showed that pharmacological inhibition of either FGF or BMP signaling during somitogenesis causes a failure of thyroid anlage formation in zebrafish (21). Thus, we reasoned that reduced availability of cardiac-borne signaling cues might explain the defective thyroid anlage formation observed in our experimental models (Wnt overactivation, mef2c/d-morphants) displaying perturbed cardiac development in this study.
To address the question that diminished BMP signaling might contribute to thyroid phenotypes observed in embryos after Wnt overactivation, we first examined endogenous bmp4 expression in control embryos and embryos treated with BIO from 6 to 10 hpf. WISH analyses of 26 hpf embryos showed that the bmp4 expression of cardiac tissue adjacent to the thyroid anlage region in normally developing embryos (Supplementary Fig. S9A) is completely ablated in BIO-treated embryos (Supplementary Fig. S9B). Similarly, Hinits et al. reported that Mef2c-deficient embryos lack bmp4 expression in the cardiac region (51). In normally developing 42 hpf embryos, we also observed that cells contained in the forming thyroid bud display enhanced BMP signaling compared with the adjacent foregut tissue (Supplementary Fig. S9C–E).
Next, we used a heat-shock-inducible system to globally enhance BMP signaling during somitogenesis stages and assessed the effects of ectopically induced BMP signaling on thyroid development in control and BIO-treated embryos. For this purpose, we treated embryos from Tg(hsp70l:bmp2b) founders with either DMSO (0.1%) or BIO (5 μM) from 8 to 11 hpf and then exposed these embryos to heat-shock treatment at either early somitogenesis (11 hpf), midsomitogenesis (15 hpf), late somitogenesis (20 hpf), or repeatedly during the course of somitogenesis (at 11, 15, and 20 hpf). Subsequent WISH analyses of thyroid and cardiac markers at 28 hpf revealed different phenotypes depending on the pretreatment (DMSO, BIO) and the timing of heat-shock-induced bmp2b overexpression.
Consistent with a proposed role of BMP for thyroid cell specification, global overactivation of BMP signaling in DMSO-treated embryos resulted in enhanced expression of nkx2.4b (Fig. 5A–I). Not only was nkx2.4b staining strongly increased in these embryos, but the majority of affected embryos also showed a dramatic and irregular expansion of thyroid marker expression along the anterior–posterior axis.
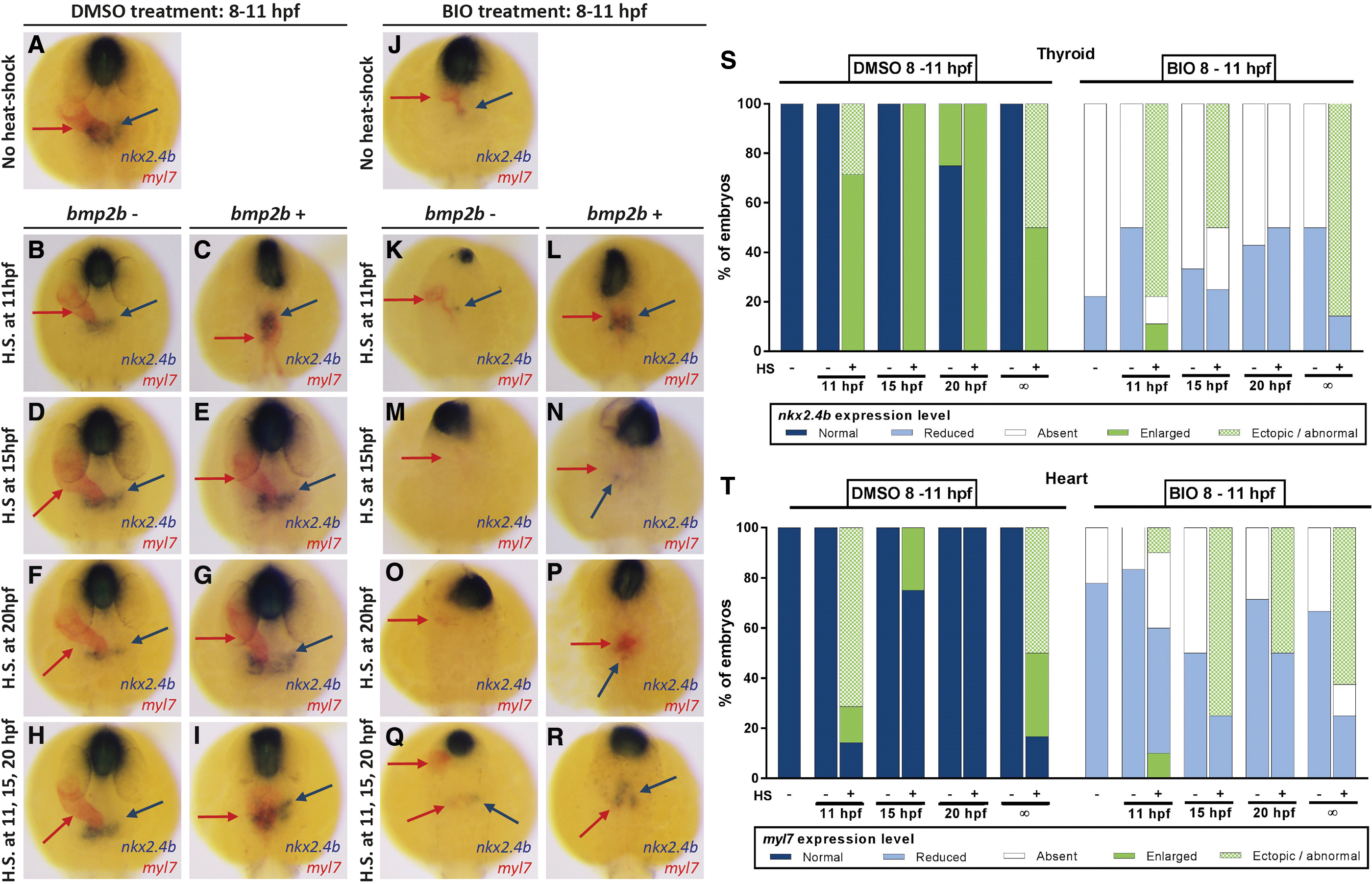
Conditional overactivation of BMP signaling in Tg(hsp70l:bmp2b) embryos partially rescues BIO-induced thyroid specification defects. (
Concurrently, DMSO-treated embryos, when heat-shocked at early somitogenesis, failed to form a normal heart tube but showed a severely perturbed expression pattern of the cardiac marker myl7 with bilateral expression domains extending irregularly along the anterior–posterior axis (Fig. 5C). When we applied heat-shock on Tg(hsp70l:bmp2b) embryos at middle or late somitogenesis stages, we observed that the thyroid anlage and primitive heart tube formed at orthotopic positions (Fig. 5D–G). We also observed a moderate enlargement of the nkx2.4b expression domain in these embryos. Embryos that were repeatedly heat-shocked during somitogenesis showed a phenotype similar to early somitogenesis heat-shock (Fig. 5H, I). Collectively, the timed heat-shock experiments in DMSO-treated embryos showed that the endoderm is competent at all somitogenesis stages to respond to BMP overactivation with an enhanced thyroid marker expression.
Heat-shock experiments with embryos pretreated with BIO (5 μM) from 8 to 11 hpf showed that global overactivation of BMP signaling could partially rescue the BIO-induced lack of thyroid marker expression at 28 hpf (Fig. 5J–R). WISH analyses and subsequent genotyping of stained specimens showed that heat-shock treatment of nontransgenic BIO-treated embryos did not improve nkx2.4b expression, irrespective of the timing of heat-shock induction during somitogenesis.
In stark contrast, BIO-treated embryos carrying the hsp70l:bmp2b transgene showed a strong nkx2.4b expression in the thyroid region when heat-shock induction of BMP signaling was performed at early somitogenesis (11 hpf) (Fig. 5K, L) or when heat-shock was repeatedly applied at 11, 15, and 20 hpf (Fig. 5Q, R). Just as observed in DMSO-treated embryos (Fig. 5C), overactivation of BMP signaling in BIO-treated embryos at early somitogenesis resulted in irregularly shaped and positioned nkx2.4b expression domains, often characterized by an elongated shape along the anterior–posterior axis (Fig. 5L). Additional WISH analyses for another early thyroid marker, hhex, showed a similar rescue of thyroid hhex-positive cells in AZA-treated embryos (Supplementary Fig. S10G, H).
The capacity of BMP overactivation to rescue thyroid marker expression in BIO-treated embryos decreased markedly at later somitogenesis stages (Fig. 5N), and no rescue of nkx2.4b expression was detectable for heat-shock treatments carried out at 20 hpf (see Fig. 5P). Thus, in contrast to DMSO-treated embryos, embryos treated with BIO lacked the competence to enhance thyroidal nkx2.4b expression in response to overactivation of BMP signaling at late somitogenesis stages. Accordingly, in this model, enhancing BMP signaling alone at late somitogenesis stages is not sufficient to rescue thyroid specification defects.
One characteristic feature of BIO-treated embryos displaying a rescue in nkx2.4b expression following early heat-shock treatment (11 hpf) was the presence of irregular ectopic patches of myl7-expressing cells near the nkx2.4b-expressing cells. Although we observed variable amounts of myl7-expressing cells across individual embryos from all heat-shock treatment groups (Fig. 5N, P), it was only in the group of BIO-treated embryos receiving heat-shock treatment at early somitogenesis (11 hpf) that we consistently observed a surplus of myl7-expressing cells in the vicinity of nkx2.4b-expressing cells. However, we note that we never observed a near full rescue of heart tube formation due to our heat-shock treatments in BIO-treated embryos.
Enhancing BMP signaling causes partial rescue of thyroid defects in Mef2c-deficient embryos
Given that enhanced BMP signaling could only achieve a rescue of thyroid specification defects in BIO-treated embryos under conditions that concurrently induced a small but consistent cardiac cell differentiation, we next applied a similar rescue approach to Mef2c-deficient embryos. The Mef2c-deficient model provides a promising alternative rescue scenario as global BMP overactivation was deemed unlikely to overcome the block of cardiomyocyte differentiation resulting from the deficiency of Mef2 protein expression. In these experiments, we injected embryos from Tg(hsp70l:bmp2b) founders with mef2c/d-MO or maintained Tg(hsp70l:bmp2b) embryos as a noninjected control group. Injected and noninjected embryos were then heat-shock treated at early (10 hpf), middle (15 hpf), or late somitogenesis (20 hpf) and thyroid and cardiac markers were analyzed at 28 hpf (Fig. 6).
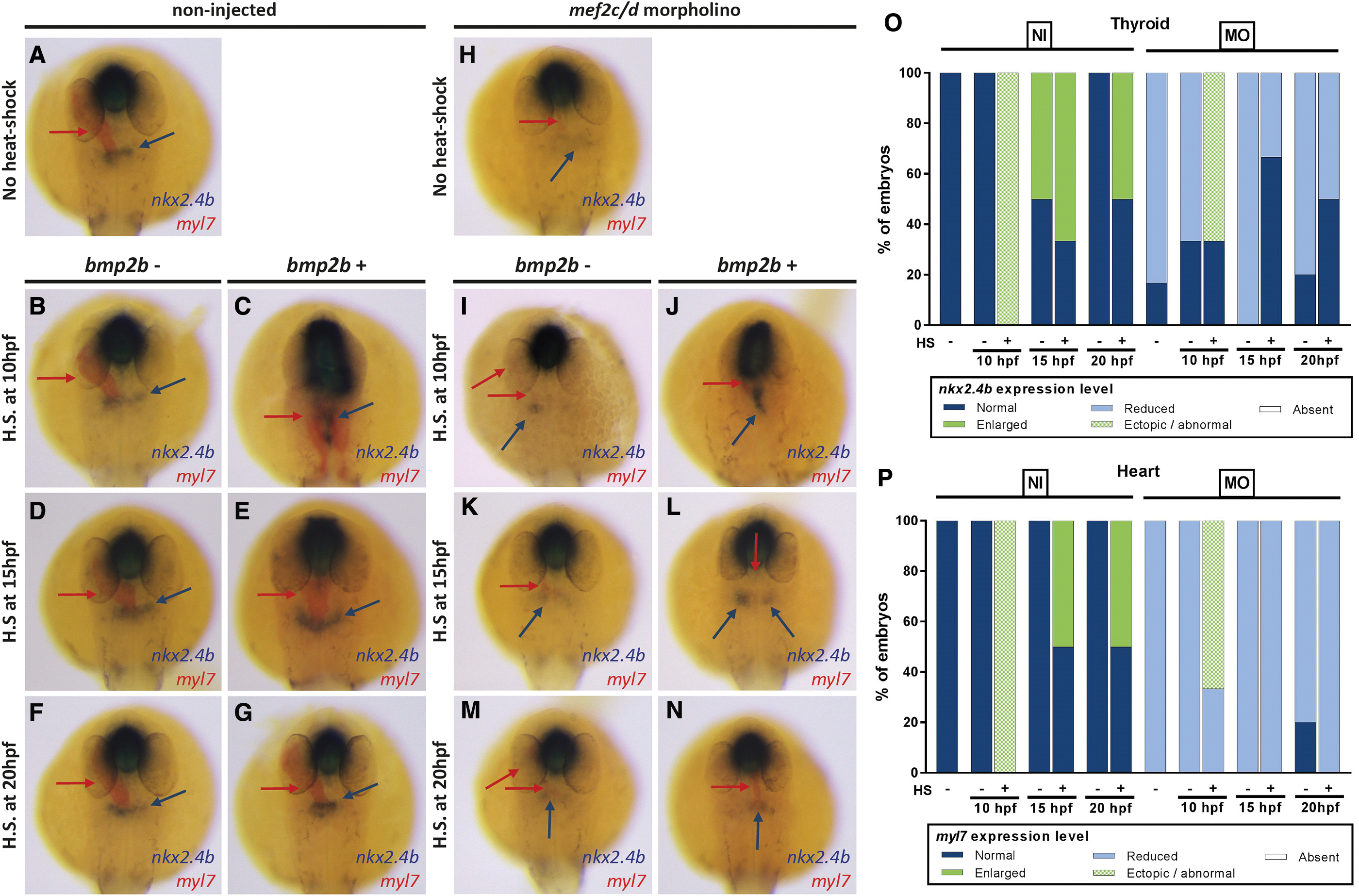
Conditional overactivation of BMP signaling in Tg(hsp70l:bmp2b) embryos partially rescues mef2c/d MO-induced thyroid specification defects. (
In the non-injected control embryos, we observed changes in thyroid and cardiac development (Fig. 6A–G) that were very similar to the effects seen in DMSO-treated control embryos in the previous experimental series (Fig. 5A–G). Depending on the timing of heat-shock treatment, we observed ectopic expansions of nkx2.4b expression along the anterior–posterior axis (heat-shock at 10 hpf) or moderately enhanced nkx2.4b expression when heat-shock was applied at middle (15 hpf) or late somitogenesis stages (20 hpf). In addition to the irregular shape and position of the thyroidal nkx2.4b expression domain, we noted again abnormal myl7 expression domains, irregularly extending along the anterior–posterior axis (Fig. 6C).
WISH analyses of the thyroid and cardiac markers expression of Mef2c-deficient embryos at 28 hpf showed a considerable reduction in the thyroid nkx2.4b staining (Fig. 6H). The severity of the thyroid specification defects correlated with the severity of a concurrent cardiac developmental phenotype, as judged from the expression of myl7. As expected, irrespective of their timing, heat-shocks of non-transgenic Mef2c-deficient embryos failed to rescue the thyroid or cardiac phenotype (Fig. 6I, K, and M). In Mef2c-deficient embryos carrying the hsp70l:bmp2b transgene, heat-shock induction of BMP signaling at early somitogenesis (10 hpf) resulted in a robust thyroid nkx2.4b expression, even though the thyroid's shape and position were abnormal (Fig. 6J). This phenotype largely resembled the expansion along the anterior–posterior axis seen in control embryos heat-shocked at 10 hpf (Fig. 6C).
Similar to the series of heat-shock experiments performed on BIO-treated embryos, a partial rescue of thyroid specification defects was generally limited to heat-shock induction of BMP signaling at early somitogenesis stages. In Mef2c-deficient embryos carrying the hsp70l:bmp2b transgene, heat-shock induction of BMP signaling at middle (15 hpf) or late somitogenesis (20 hpf) only mildly increased levels of nkx2.4b expression compared with untreated mef2c/d morphants (Fig. 6L, N). Compared with the thyroid rescue observed in BIO-treated embryos, a notable difference in the Mef2c-deficient model was that heat-shock induction of BMP signaling did not rescue the cardiac differentiation phenotype. Thus, our observations in the cardiac-deficient mef2c/d morphant model indicate that ectopically induced BMP signaling is sufficient to rescue the thyroid specification defect even in the absence of cardiomyocyte differentiation.
The rescue of thyroid tissue was visible with two different early thyroid markers (nkx2.4b and hhex). We tried to confirm these results by tracing specific thyroid markers such as tg. However, these markers only appear at later times (>50 hpf), which, combined with the partial and limited time efficiency of the MO used (resulting in incomplete cardiac ablation), makes that at this later stage thyroid rescue is not evident (data not shown). It is likely that early defects in thyroid development are compensated by delayed catching up of cardiomyocyte differentiation and the subsequent adequate supply of ligands, allowing the induction of thyroid specification into the endoderm.
Discussion
In this study, we described severe defects in early zebrafish thyroid development as a result of globally modulated canonical Wnt signaling during gastrula and early somitogenesis stages. While TD in response to drug-induced Wnt signaling modulation was first detected in the course of our recent small-molecule screening study with zebrafish embryos (21), the current study provides corroborative evidence that the thyroid specification defects observed following treatment with BIO and AZA are due to the upregulation of canonical Wnt signaling. We used a well-characterized zebrafish biosensor line (34) to confirm that BIO and AZA treatments are effectively inducing canonical Wnt signaling in vivo during zebrafish development.
Our results are, therefore, in line with numerous studies in zebrafish, Xenopus, and mammalian cell systems utilizing these two compounds to globally induce Wnt signaling (34,58,59). Importantly, the thyroid specification defects observed in drugged embryos were faithfully reproduced in a genetic zebrafish model, thereby corroborating critical evidence that the thyroid defects are indeed caused by enhanced Wnt signaling and are not due to possible nonspecific side effects of drug treatments.
Thyroid phenotyping of drugged embryos at the thyroid anlage stage (28 hpf), using nkx2.4b expression as a proxy for thyroid lineage commitment, revealed concentration-dependent losses of nkx2.4b-expressing cells in the prospective thyroid region. In zebrafish embryos, thyroidal expression of nkx2.4b typically becomes detectable at around 23/24 hpf, and a global assessment of developmental timing in drugged embryos indicated that reduced detection of nkx2.4b expression was not solely due to delayed development. Remarkably, gastrula treatment with a high concentration (5 μM) of BIO or AZA caused a complete absence of detectable nkx2.4b expression in 28 hpf embryos.
Although the latter phenotype was highly penetrant at 28 hpf (affecting almost 100% of drugged embryos), analyses of treated embryos at later stages (55 hpf) showed that almost all treated embryos developed at least a very tiny thyroid primordium comprising very few cells expressing the early functional thyroid marker tg. It is currently unknown if these few remaining tg-expressing thyroid cells at 55 hpf are derived from thyroid precursors expressing nkx2.4b at such low levels that are undetectable by our WISH at 28 hpf or if some precursor cells are specified after 28 hpf to give rise to tg-expressing thyroid cells. Despite the presence of very few differentiated thyroid cells at later stages, we can conclude that increased canonical Wnt signaling during early zebrafish development impairs the specification of thyroid precursor cells.
By progressively shifting the timing of short-term drug treatments, we observed that the primary events leading to this TD phenotype occur during gastrula and early somitogenesis stages. Therefore, the question arises whether increased Wnt signaling directly affects the development of endodermal progenitors, later giving rise to the foregut endoderm and the thyroid cell lineage, or whether indirectly through another cell type (e.g., precardiac mesoderm) that is required later in the development as a local source of signals to initiate thyroid specification in the foregut endoderm zebrafish (18,27).
During early vertebrate development, Wnt signaling plays a key role in the anteroposterior patterning, mainly by acting as a posteriorizing factor (60). This anterior–posterior patterning role of Wnt signaling has been extensively studied during neural development (54). In our studies, a robust posteriorizing activity of Wnt on neural development was evident from the severe forebrain and eye development defects. Since the thyroid develops from the anterior foregut endoderm, the posteriorizing activity of Wnt on the endoderm could provide a plausible hypothesis to explain the observed TD. Surprisingly, few studies have addressed the precise role of Wnt on endoderm patterning. Studies in Xenopus embryos indicated that canonical Wnt signaling during gastrulation might pattern the endoderm in a way much similar to what is known for the nervous system (27). Notably, forced Wnt8 expression in cells fated to become endoderm was found to block anterior endoderm development in Xenopus embryos, resulting in specification failure of foregut organ primordia.
In contrast to the aforementioned Xenopus studies, morphological and molecular analyses of thyroid-lacking zebrafish embryos revealed the formation of a morphologically fairly normal anterior endoderm with a foregut identity (foxa2 expression) and confirmed timely expression of various other foregut organ markers (hhex, pdx1, prox1a). Moreover, data presented by McLin et al. (27) indicate that thyroid specification was preserved in Xenopus embryos if enhanced β-catenin signaling was cell autonomously restricted to the endoderm lineage. When comparing these study results with our observations, we conclude that posteriorization effects of BIO-induced Wnt activity were by far not as dramatic as reported in the Xenopus studies and that posteriorization of anterior endoderm patterning was likely not the primary cause for the severe thyroid specification defects in zebrafish embryos.
Another striking effect of drug-induced Wnt activation was a severe impairment of cardiac differentiation. Consistent with previous studies, we confirmed that enhanced Wnt signaling during zebrafish gastrulation in zebrafish limits cardiac differentiation within the precardiac mesoderm (25,26). Intriguingly, we observed a close correlation between the disruption of cardiac progenitor differentiation and the thyroid abnormalities. This was true for experiments involving increasing concentrations of BIO and AZA, as well as for experiments with different treatment periods. In this respect, it is noteworthy that treatment periods, which caused little if any global posteriorization effects, still showed reduced cardiac differentiation concurrent with thyroid specification defects. Thus, the apparent relationship between the diminished formation of cardiac tissue and the corresponding defects in thyroid specification led us to formulate the hypothesis that the primary effect of enhanced Wnt signaling might be the blockage of cardiac differentiation, while the thyroid defects might be a secondary event resulting from a reduction of cardiac-borne signaling cues.
This hypothesis is supported by results from several previous studies in different vertebrate models, which provided evidence for a critical role of precardiac and cardiac mesoderm in the induction of thyroid precursor cell signaling (17,18). Our own studies provided further supporting lines of evidence for this hypothesis by showing that a similar correlation between impaired cardiogenesis and defective thyroid specification exists in Mef2c-deficient embryos (51), a model in which heart formation is blocked independent of perturbations in Wnt signaling. Since the mesoderm, and not the foregut endoderm, expresses mef2ca and mef2cb, this model provided more direct evidence for the contention that thyroid specification relies on proper differentiation of cardiac tissue. A role of mesoderm as a source of signals coming to induce specification of thyroid progenitors (directly or indirectly by making the anterior endoderm more competent to specify into thyroid anlage) within the foregut endoderm is also consistent with what has been already described concerning the specification of other endoderm-derived organs (61 –63).
While these observations strongly suggest an important role of cardiac mesoderm as a signal source involved in the early steps of thyroid development, it raises the question of the nature of the signaling cues involved in this tissue–tissue interaction. Based on results from our small-molecule screening (21) and previous studies in several other model systems, BMP signaling was deemed a primary candidate of particular importance for thyroid specification (56).
In vitro experiments with murine stem cell models identified key conserved roles for BMP signaling in regulating thyroid lineage specification from the foregut endoderm (56). On the contrary, Wnt was not required for thyroid specification from the endoderm in vitro, like it had been previously suggested (64). Expression profiles during embryogenesis also support a potential role of BMP signaling as a key factor in regulating early steps of thyroid development. In mouse, BMP4 is the Bmp ligand expressed at high levels by the cardiac mesoderm, in particular within the secondary heart field, the structure that gives rise among others to the outflow tract, which the developing thyroid lies nearby (65,66).
Danesh et al. also demonstrated in mouse embryos that BMPR1a is highly expressed in regions of the pharyngeal endoderm, where endodermal progenitors will differentiate into thyroid cells (66). Thus, Bmp4 could be one critical endogenous Bmp ligand acting from mesoderm to endoderm to initiate thyroid development through interaction with its receptor Bmpr1a. For zebrafish, we further confirmed a high bmp4 expression in cardiac mesoderm around the time of thyroid specification (Supplementary Fig. S9), and ectopic BMP signaling during somitogenesis caused an expansion of the thyroid anlage (Figs. 5 and 6).
Against this background, it is noteworthy that timed overactivation of BMP signaling during zebrafish somitogenesis stages could partially restore thyroid specification in both models of impaired cardiac development. Interestingly, in our model of cardiac dysgenesis due to enhanced Wnt signaling, the restoration of thyroidal nkx2.4b expression achieved by subsequent BMP overactivation appeared strictly dependent on a concurrent rescue of cardiac cell differentiation. At this stage, it is therefore not possible to conclude if thyroid specification was directly restored by the ectopic BMP activity in this model.
Several lines of evidence indicate that thyroid specification requires the combined action of BMP and FGF signaling (17,18,56,57,67). One possible interpretation might be that BMP signaling alone might not be sufficient to induce thyroid specification in embryos with almost no cardiac mesoderm formation. In contrast, restoration to some extent of cardiac differentiation by transient BMP overactivation could provide a source of endogenous BMP, FGF, and possibly other signaling cues to induce thyroid specification.
Rescue of thyroid specification was also evident in Mef2c-deficient embryos after BMP overactivation. However, in this model, the BMP-induced thyroid rescue was less explicit, possibly due to the incomplete penetrance of the cardiac phenotype in morphant embryos. Still, we observed many Mef2c-deficient embryos lacking cardiomyocytes but presenting a partial thyroid rescue. These observations indicate that ectopic BMP activation might indeed restore thyroid specification even in the absence of detectable cardiomyocytes. To understand the differences between the thyroid rescue induced by ectopic BMP activity in the two cardiac dysgenesis models, the different mechanisms leading to deficient cardiomyocyte development need to be taken into account. Enhanced Wnt signaling at early stages leads to a block of early cardiac mesoderm differentiation with a pronounced deficit in cardiac progenitor differentiation (Supplementary Fig. S7).
In contrast, studies by Hinits et al. (51) showed that early cardiac development is uncompromised in Mef2c-deficient embryos and that the last steps involved in cardiomyocyte differentiation are blocked in this model, including the upregulation of bmp4 expression in the cardiac field. Thus, the precardiac mesoderm present in Mef2c-deficient embryos might serve as a source of FGF and other signaling factors needed to act in concert with BMP to induce thyroid specification. Given that Mef2c-deficient embryos lack bmp4 expression in the cardiac mesoderm and that ectopic overexpression of mef2cb mRNA can induce bmp4 in the cardiac field (51), it would be interesting to investigate further this model, with respect to the expression of other potential thyroid-inducing pathways (e.g., FGF) and their role in acting together with BMP signaling in the process of thyroid specification. Also, results obtained in Mef2c-deficient embryos indicate that the thyroid of Bmp4-deficient embryo models might bring further insight into the role of BMP signaling in thyroid specification.
In summary, our studies confirmed and validated our previous results from a small-molecule screen that enhanced Wnt signaling during gastrulation and early somitogenesis stages results in severe inhibition of thyroid specification. Moreover, we provide corroborating evidence for the contention that this TD results from a blockage of cardiac development and, consequently, the lack of instructive cardiac-derived signaling cues for the specification of thyroid precursor cells within the foregut endoderm. This hypothesis is supported by a similar perturbation of thyroid specification in Mef2c-deficient embryos displaying a lack of cardiomyocyte differentiation. Finally, we demonstrate that ectopic activation of BMP signaling can partially rescue these thyroid phenotypes identifying BMP signaling as one critical component of this tissue–tissue interaction.
Footnotes
Acknowledgments
We thank the laboratory that provided us transgenic lines. We greatly appreciate the help of J. Bakkers, E. Moro, S. Abdelilah-Seyfried, and G. Weidinger for providing us with transgenic lines used in this study. J. Bakkers for Tg(kdrl:EGFP) and Tg(myl7:EGFP) lines; E. Moro for Tg(7xTCF-Xla.Siam:GFP); S. Abdelilah-Seyfried for Tg(hsp70l:bmp2b) line; and G. Weidinger for Tg(hsp70l:wnt8a-EGFP). We thank Véronique Janssens for the technical assistance.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the Belgian National Fund for Scientific Research (FNRS) (FRSM 3-4598-12; CDR-J.0145.16), the Action de Recherche Concertée (ARC) de la Communauté Française de Belgique (ARC AUWB-2012-12/17-ULB3), the Fonds d'Encouragement à la Recherche de l'Université Libre de Bruxelles (FER-ULB), the Fund Yvonne Smits (King Baudouin Fundation) and the Berlin Institute of Health (BIH, CRG-TP2). I.V. is FNRS research fellow, B.H., P.G are Fund for Research in the Industry and the Agriculture (FRIA) research fellows; R.O is FNRS postdoctoral fellow; S.C. is FNRS research Director.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Figure S9
Supplementary Figure S10