
Abstract
Background:
Thyrotropin-releasing hormone (TRH) neurons in the paraventricular nucleus of the hypothalamus (PVN) have been identified as direct regulators of thyrotropin (TSH) and thyroid hormone (TH) levels. They play a significant role in context of negative feedback by TH at the level of TRH gene expression and during fasting when TH levels fall due, in part, to suppression of TRH gene expression.
Methods:
To test these functions directly for the first time, we used a chemogenetic approach and activated PVN TRH neurons in both fed and fasted mice. Next, to demonstrate the signals that regulate the fasting response in TRH neurons, we activated or inhibited agouti-related protein (AgRP)/neuropeptide Y (NPY) neurons in the arcuate nucleus of the hypothalamus of fed or fasted mice, respectively. To determine if the same TRH neurons responsive to melanocortin signaling mediate negative feedback by TH, we disrupted the thyroid hormone receptor beta (TRβ) in all melanocortin 4 receptor (MC4R) neurons in the PVN.
Results:
Activation of TRH neurons led to increased TSH and TH levels within 2 hours demonstrating the specific role of PVN TRH neurons in the regulation of the hypothalamic–pituitary–thyroid (HPT) axis. Moreover, activation of PVN TRH neurons prevented the fall in TH levels in fasting mice. Stimulation of AgRP/NPY neurons led to a fall in TH levels despite increasing feeding. Inhibition of these same neurons prevented the fall in TH levels during a fast presumably via their ability to directly regulate PVN TRH neurons via, in part, the MC4R. Surprisingly, TH-mediated feedback was not impaired in mice lacking TRβ in MC4R neurons.
Conclusions:
TRH neurons are major regulators of the HPT axis and the fasting-induced suppression of TH levels. The latter relies, at least in part, on the activation of AgRP/NPY neurons in the arcuate nucleus. Interestingly, present data do not support an important role for TRβ signaling in regulating MC4R neurons in the PVN. Thus, it remains possible that different subsets of TRH neurons in the PVN mediate responses to energy balance and to TH feedback.
Introduction
The discovery of thyrotropin-releasing hormone (TRH) was pivotal to the understanding of the brain and its endocrine role, as well as our understanding of physiological and pathological phenomena, giving rise to the field of neuroendocrinology. 1 –3 Crucial for adequate production of thyroid hormones (TH), TRH neurons of the hypothalamic paraventricular nucleus (PVN) control thyrotropin (TSH) synthesis, secretion, and bioactivity. 4 –6 Therefore, the regulation of circulating TH levels depends on the proper regulation of these neurons. Importantly, TH levels feedback at the level of the TRH neuron in the PVN such that TRH levels are high in primary hypothyroidism when TH levels are low. Impaired TRH signaling has important impacts on human health and has been linked to central hypothyroidism, subclinical hypothyroidism, as well as to resistance to TH. 7 –9
Hypothalamic TRH neurons are critical metabolic regulators, 10 and their activity is affected by variations in both internal and external environments. For example, energy availability has remarkable effects on hypothalamic TRH neurons. Most interesting, a negative energy balance results in suppressed TRH despite lower TH levels. 11,12 Hence, although the negative feedback exerted by TH 13 –15 on TRH expression, TRH neurons can be strongly regulated by other signaling systems.
The hypothalamic arcuate nucleus (ARC) is an important metabolic center, encompassing neurons critical for appetite control, and required for proper hypothalamic–pituitary–thyroid (HPT) axis regulation. 16,17 Different neuropeptides synthesized by ARC neurons, such as alpha-melanocyte-stimulating hormone (α-MSH), neuropeptide Y (NPY), and agouti-related protein (AgRP), have been implicated in regulation of the HPT axis. 18 –22 We and others have proposed the melanocortin system as an important regulator of the HPT axis. The melanocortin 4 receptor (MC4R) is colocalized with TRH in the PVN, 23 and α-MSH and AgRP terminals have been described connecting to TRH neurons. 24 Furthermore, MC4R signaling activates the TRH promoter, 23 and administration of these arcuate peptides regulates the HPT axis. 21,25 Therefore, the melanocortin system can potentially control metabolism and energy expenditure through the regulation of TRH neurons and TH levels.
Herein, we addressed the causal connection between TRH neuron inhibition and HPT axis suppression during fasting; the role of ARC AgRP/NPY neurons in regulating the HPT axis; and the relevance of thyroid hormone receptor β (TRβ) signaling in MC4R-positive neurons in the PVN, which have been hypothesized as integrators of metabolic and TH signaling. Data presented here support an important role for ARC-PVN circuitry in regulating the HPT axis and challenge the current paradigm of how TRH neurons integrate melanocortin and thyroid signaling.
Materials and Methods
Methods are provided in further detail in the Supplementary Data.
Animals
All animal procedures were reviewed and approved by Beth Israel Deaconess Medical Center and Weill Cornell Medicine Institutional Animal Care and Use Committees (Nos. 001-2008, 008-2011, 098-2013, 099-2016; BIDMC/Harvard, Boston, MA, and No. 2018-029; WCM, New York, NY). Mice were housed at 22 ± 1°C, under a 12/12-hour light/dark cycle with standard mouse chow and water available ad libitum, unless otherwise stated. The following animals were used in this study: AgRP-IRES-Cre, 26 MC4R-2a-Cre, 27 and TRH-IRES-Cre 28 were provided by Brad Lowell (BIDMC); mTmG 29 provided by Laura Alonso (WCM); Thrb flox/flox 30 provided by Karine Gauthier (Université de Lyon, France).
Determination of circulating hormone levels
Blood samples were collected as above described. Enzyme-linked immunosorbent assay (Diagnostic Automation/Cortez Diagnostics, Inc.) was used to measure thyroxine (T4). Serum TSH was measured by Luminex Multiplex (EMD Millipore).
RNA extraction and quantitative reverse transcription-polymerase chain reaction
Brains were removed, and sagittal sections were taken for PVN microdissection. PVN, pituitary, and liver were quickly removed and frozen in liquid nitrogen and subsequently stored at −80°C. PVN and pituitary RNA were extracted using Direct-zol RNA Microprep Kit (Cat. No. R2062; Zymo Research). RNA from the liver was extracted using STAT-60 reagent (Tel-Test, Friendswood, TX). Then, total RNA was reverse transcribed using SuperScript VILO cDNA Synthesis Kit (Invitrogen, Carlsbad, CA). TaqMan gene expression assays for all messenger RNAs (mRNAs) were purchased from Life Technologies (Carlsbad, CA). Quantitative PCR was performed in duplicate using the 800 HT thermal cycler (Life Technologies, Foster City, CA). Relative mRNA levels were calculated using the standard curve method and normalized to the level of 18S ribosomal RNA
Transcardiac perfusion and immunofluorescence
Animals were deeply anesthetized by i.p. injection of ketamine (100 mg/kg) and xylazine (10 mg/kg), perfused transcardially with 0.9% phosphate-buffered saline (PBS), and followed by 4% neutral buffered formalin. The cerebral tissue was post-fixed in the same fixative overnight, equilibrated in 20% sucrose, and cut into 25-μm coronal sections on a cryostat. A series of 25 μm cryosections were processed for immunofluorescence for mCherry, green fluorescent protein (GFP), and cFos.
We performed immunofluorescence for mCherry and cFos to check the efficiency of viral infection and designer receptor exclusively activated by designer drugs (DREADD) 31 transduction and also to certify that our clozapine-n-oxide (CNO) injection can activate the DREADD. Briefly, the brain slices were equilibrated in phosphate-buffered saline with Tween 20 (PBST) buffer, the nonspecific binding was blocked with a solution of 5% normal goat serum and overnight incubated with a polyclonal rabbit anti-cFos antibody (1:100, Cat. No. sc-52; Santa Cruz) in PBST buffer. The next day, the tissues were rinsed with PBST and incubated for 1 hour with Alexa 488 donkey-anti-rabbit antibody (1:500, Cat. No. 711-545-152; Jackson ImmunoResearch). The sections were placed on slides and covered with mounting media. The endogenous fluorescence from mCherry can be observed without any immunostaining detection. GFP was detected using a similar protocol and a polyclonal goat anti-GFP conjugated to fluorescein isothiocyanate (ab6662; Abcam).
Statistical analyses
Statistical analyses were performed using GraphPad Prism version 9. Two- or three-way analysis of variance was performed, followed by Bonferroni's multiple comparison test. Unpaired t-test was used when comparing only two groups. The number of animals used in each study is described in the figure legends.
Results
Chemogenetic stimulation of TRH neurons activates the HPT axis
It has long been known that the hypothalamic TRH neuron plays a critical role in regulating the HPT axis. 32 Indeed, Kondo et al. demonstrated that deleting Trh specifically from the PVN results in reduced T4 levels despite increased circulating TSH levels. 6 We used a chemogenetic approach (Fig. 1A) to directly address the role of PVN TRH neurons in regulating TSH and T4 levels, by stereotaxically introducing in the PVN an M3Dq receptor linked to mCherry that can be expressed only in the presence of the Cre recombinase (Fig. 1A). We hypothesized that TRH neuronal stimulation would quickly increase TSH and T4 levels. Therefore, we activated this receptor by the exogenous ligand CNO and euthanized the mice 2 hours later.
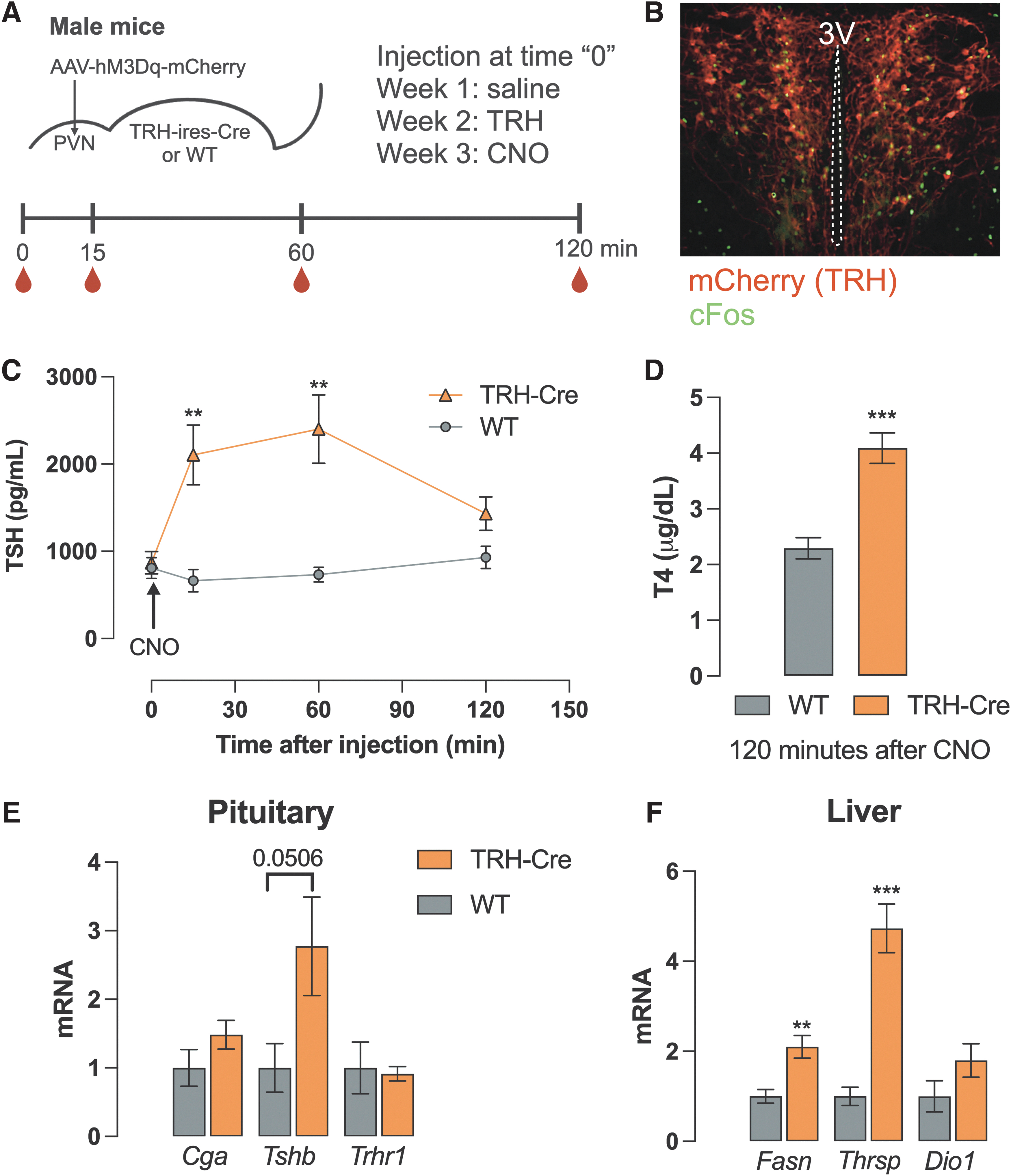
Effects of chemogenetic stimulation of TRH neurons in the PVN. Diagrammatic description of the experimental protocol, where the red drops indicate blood collection times (
The appropriate location of the hM3Dq receptor and effective neuronal activation by CNO were determined using mCherry and cFos expression, as shown in Figure 1B. Plasma TSH levels were similar between TRH-IRES-Cre mice or wild-type (WT) controls before CNO administration. Fifteen minutes after CNO administration, TSH levels were more than 300% higher in TRH-IRES-Cre mice when compared with the control group at the same time point. TSH levels remained elevated for at least 60 minutes and then fell to control levels after 2 hours (Fig. 1C). After 2 hours of CNO injection, T4 levels were significantly elevated in TRH-IRES-Cre mice (4.09 ± 0.27 vs. 2.29 ± 0.19 μg/dL, p < 0.001) (Fig. 1D), as well as the expression of TH targets in the liver, such as Thrsp and Fasn (Fig. 1F). Interestingly, Dio1 levels did not rise, which may reflect the kinetics of its induction by TH.
For instance, stimulation of Thrsp expression by a TRβ agonist is quicker than Dio1 in a rat model. 33 Additionally, we cannot exclude the presence of other factors contributing to this quick rise in Thrsp and Fasn mRNA expression, such as increased food intake. Indeed, activation of TRH neuronal circuitry has recently been demonstrated to activate food intake. 34 Despite increased T4 levels, pituitary Tshb expression was also elevated by almost three times in TRH-IRES-Cre mice (p = 0.05) (Fig. 1E). Similar effects were observed in female mice (Supplementary Fig. S1A). Saline administration had no effect in TRH-IRES-Cre mice, and TRH administration itself had comparable effects on control and TRH-IRES-Cre groups in terms of increasing TSH levels (Supplementary Fig. S1B). Taken together, the activation of TRH neurons in the PVN directly leads to the activation of the HPT axis.
Activation of TRH neurons restores TSH and T4 levels in fasted mice
Mechanisms controlling the HPT axis response to a negative energy balance remain to be elucidated. It has been recently demonstrated that TH action remains unaltered in the hypothalamus of fasting mice and does not account for HPT axis suppression. 35 Here, we tested whether inhibition of TRH neurons in the PVN is the primary cause of fasting-induced suppression of TSH and T4 levels. We hypothesized that other metabolic inputs inhibit TRH neuronal activity and thus used stimulatory DREADDs, which can be induced by Cre expressed in TRH-IRES-Cre mice to counteract that effect (Fig. 2A). A control experiment injecting saline shows that both WT and TRH-IRES-Cre mice respond normally to fasting with a fall in both their TSH and T4 levels after DREADD injection (Fig. 2B).
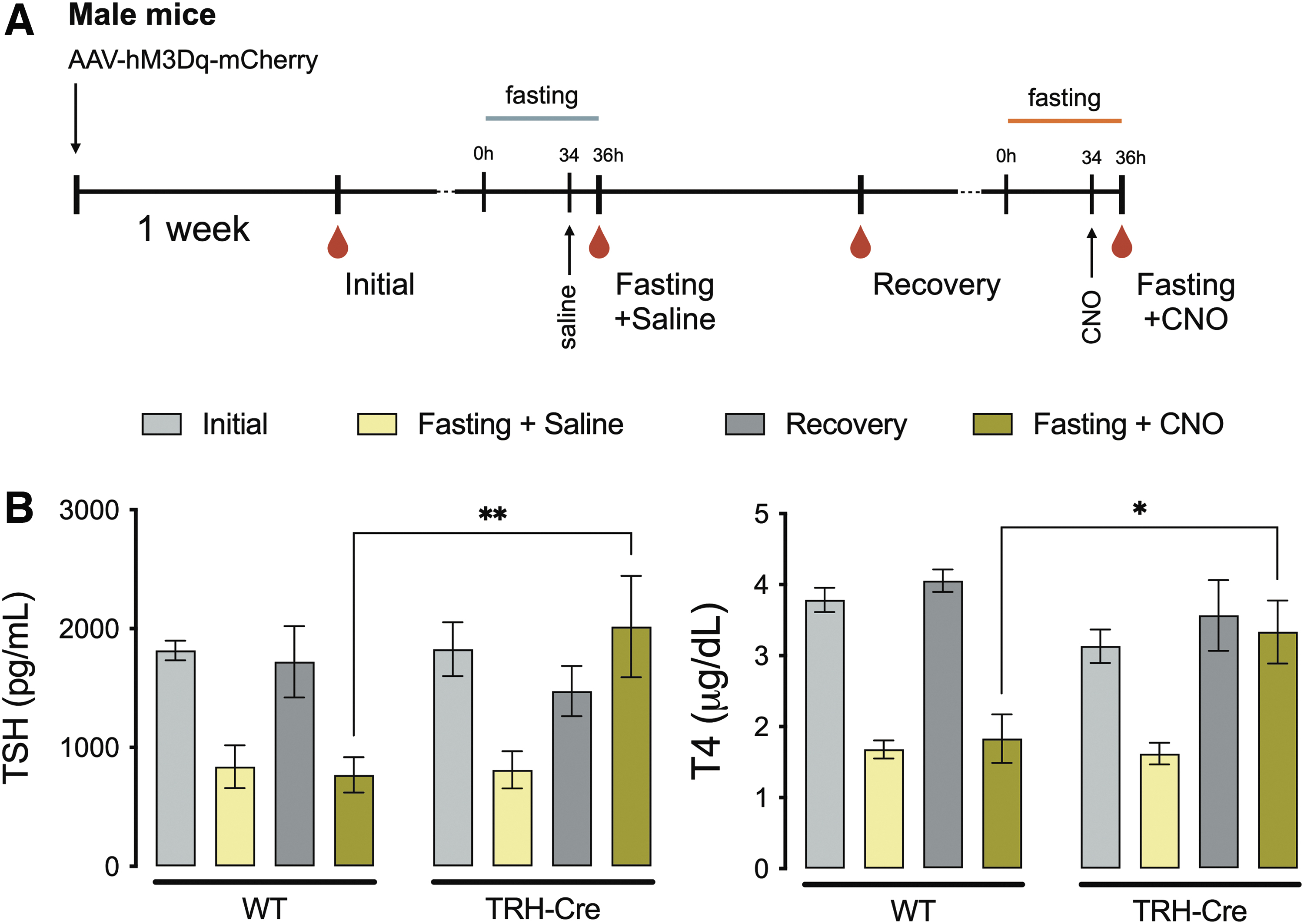
Influence of TRH activation on fasting-induced HPT axis suppression. Diagrammatic description of the experimental protocol, where the red drops indicate blood collection times (
Two weeks after the first fasting experiments, both groups showed similar TSH and T4 levels (recovery bar) and were submitted to a second fasting experiment where they received CNO 2 hours before euthanasia (34 hours into fasting). Remarkably, CNO injection restored TSH and T4 levels in TRH-IRES-Cre mice (TSH: 2017.2 ± 425.8 vs. 769.4 ± 149.4 pg/mL, p < 0.01; T4: 3.33 ± 0.44 vs. 1.83 ± 0.34 μg/dL, p < 0.05) (Fig. 2B), demonstrating that TRH neuronal inhibition is the primary cause of reduced TH levels in energy-depleted mice.
Stimulation of AgRP/NPY neurons suppresses TSH levels despite increased food intake
Importantly, arcuate nucleus (ARC) ablation disrupts the TRH neuronal response to fasting. 16 It is known that metabolic challenges, such as fasting, affect ARC neurons. Since AgRP and NPY expression increase in the ARC during fasting and AgRP/NPY neurons innervate TRH neurons in rodents and humans, 24,36 we addressed whether increased AgRP/NPY neuronal activity leads to HPT axis suppression through TRH neurons. To induce AgRP/NPY neurons over a 36-hour period, similar to what happens during fasting, we designed experiments where a stimulatory DREADD can be expressed in AgRP/NPY neurons. This was accomplished by injecting the AAV-MD3q-mCherry stereotaxically into mice that express Cre under control of the AgRP locus (AgRP-IRES-Cre).
To test for the effects of DREADD expression in the ARC and the serial injections performed, we conducted a control experiment injecting saline only (Fig. 3A) while all mice were freely eating. Both WT and AgRP-Cre mice showed similar responses (gray bars on panels C, D, E, and F) with their TSH and T4 levels remaining unchanged. Next, using CNO, we induced a sustained stimulation of AgRP/NPY neurons (Fig. 3A) in the ARC (Fig. 3B). AgRP/NPY neuronal activation resulted in increased food intake (9.93 ± 0.46 vs. 5.87 ± 0.49 g/25 g, p < 0.001) (Fig. 3C) and over 2 g of body weight gain in 36 hours (p < 0.0001) (Fig. 3D). Despite these effects, both TSH (Fig. 3E) and T4 (Fig. 3F) levels were suppressed following AgRP/NPY neuronal activation. Thus, independent of energy balance, activation of AgRP/NPY neurons leads to the suppression of the HPT axis. Similar effects were observed in female mice (Supplementary Fig. S2).
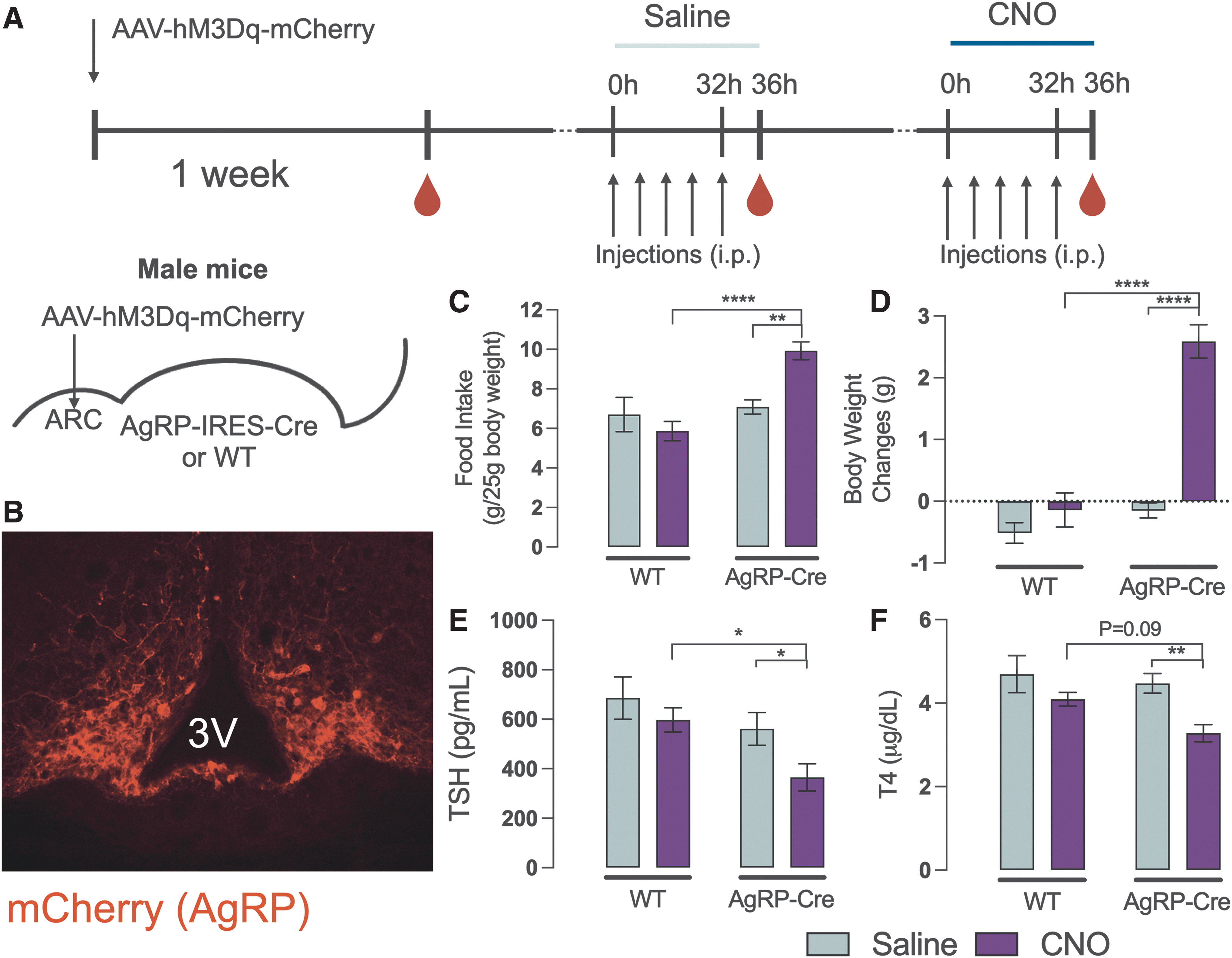
Consequences of chemogenetic activation of AgRP neurons. Diagrammatic description of the experimental protocol, where the red drops indicate blood collection times (
Inhibition of AgRP/NPY neurons blocks fasting-induced suppression of the HPT axis
Next, we wanted to determine if inhibition of AgRP/NPY neurons in the presence of a fast would prevent the reduction in TH levels. To test that, we injected an inhibitory DREADD in the ARC of AgRP-Cre mice (Fig. 4A) to inhibit AgRP/NPY neurons during fasting. An initial experiment was conducted to test the effects of the injected DREADD on the HPT response to fasting. Both control (WT) and AgRP-Cre groups responded as expected to a 36-hour fast and were similar to one another in the context of the fall in TSH and T4 levels 4 hours after the last injection of saline (Fig. 4C, first two columns).
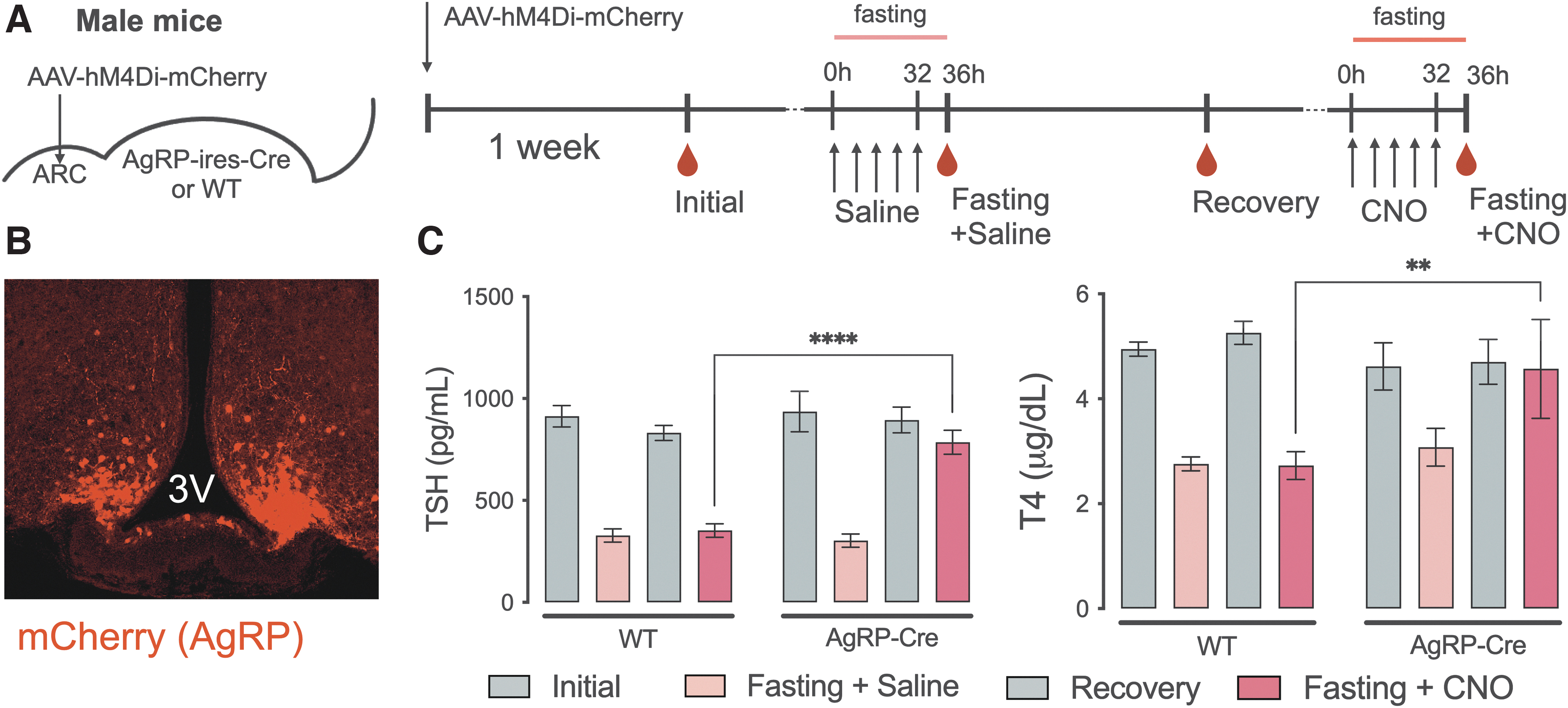
Effects of the chemogenetic inhibition of AgRP neurons during fasting. Diagrammatic description of the experimental protocol, where the red drops indicate blood collection times (
Subsequently, animals received CNO injections throughout fasting, allowing for sustained inhibition of AgRP/NPY neurons. Strikingly, AgRP-Cre mice receiving CNO showed no reduction in either TSH or T4 levels, which remained significantly higher than the control group (TSH: 786.0 ± 58.9 vs. 303.2 ± 32.5 pg/mL, p < 0.0001; T4: 4.57 ± 0.94 vs. 3.08 ± 0.36 μg/dL, p < 0.01; n = 8–10) Thus, inhibition of AgRP/NPY neurons disrupts the effects of energy depletion on the HPT axis. Similar effects were observed in female mice (Supplementary Fig. S3).
Melanocortin-sensitive neurons are not regulated by TRβ
We and others have proposed that metabolic and TH/TRβ signaling systems regulate the HPT axis via the same TRH neurons in the PVN. Evidence shows that this regulation might be integrated at the transcriptional level. 14,23,37,38 To interrogate how thyroid and ARC signaling pathways are integrated by PVN neurons, we generated MC4R-specific TRβ knockout mice utilizing MC4R-Cre mice crossed to TRβ floxed mice (TRβKO MC4R ) given the high expression of the MC4R on TRH neurons in the PVN. 23 Using this approach, recombination is detected in the PVN when using a reporter mouse line and it targets neurons projecting to the median eminence (Fig. 5A). To confirm deletion of TRβ in MC4R neurons, we performed PCR using DNA extracted from microdissected hypothalamus. In DNA extracted from the PVN, we saw clear genomic recombination in floxed mice.
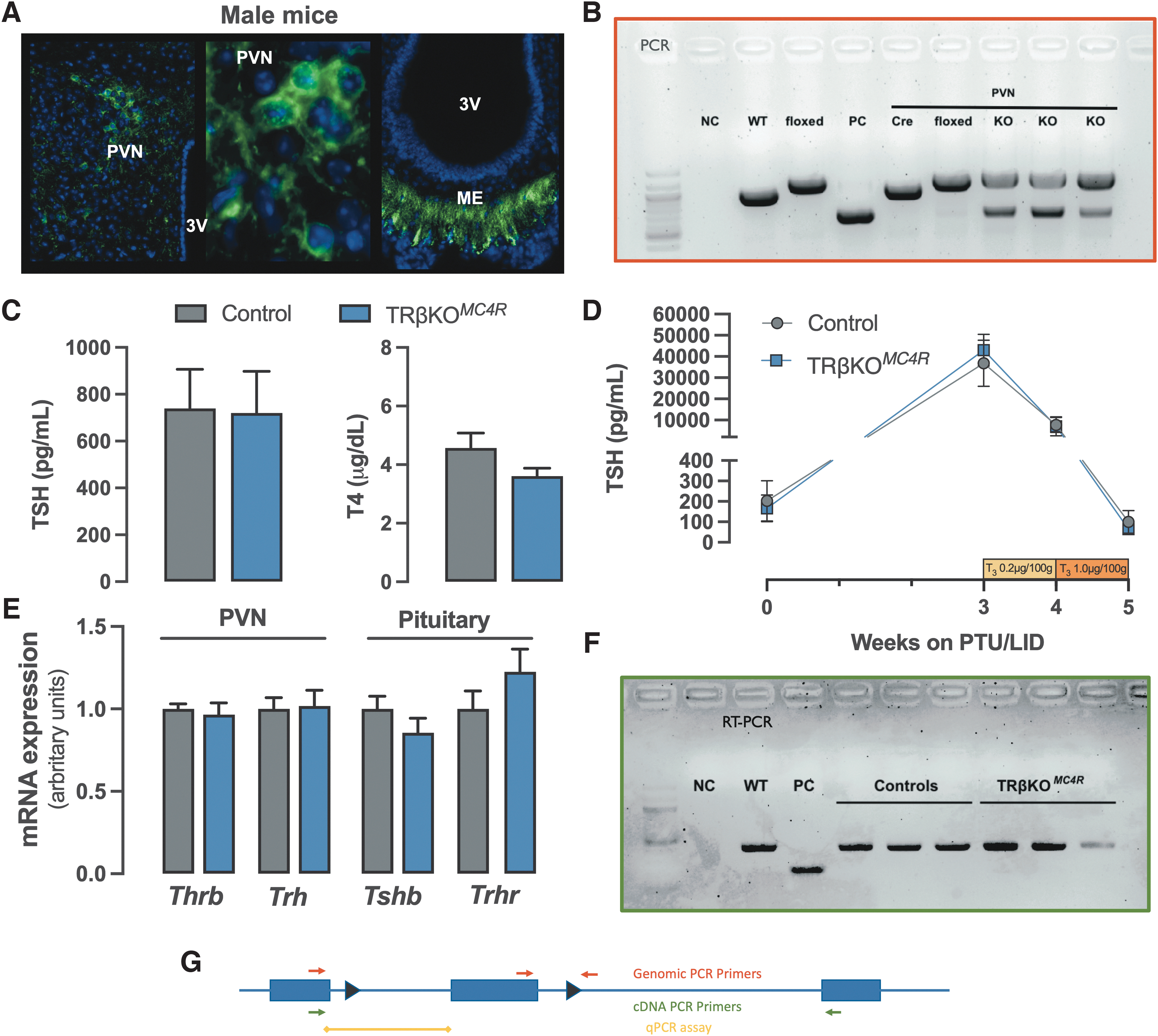
Deleting Thrb gene from MC4R-positive neurons (TRβKO
MC4R
mouse). GFP labeling is identified in the PVN and ME of a reporter mouse bred to MC4R-2a-Cre. The third ventricle is indicated (3V) (
As expected, two bands are present (floxed and recombined alleles) consistent with the fact that not every cell in the PVN expresses MC4R (Fig. 5B). Strikingly, deletion of the Thrb gene did not affect the HPT axis as TSH and T4 levels were not different between control and knockout (KO) groups (Fig. 5C). Similarly, the expression of the TH targets Trh in the PVN and Tshb, and Trhr expression in the pituitary were indistinguishable between the KO and control groups (Fig. 5E). To further interrogate the central sensitivity to triiodothyronine (T3) in the TRβKO MC4R mice, we treated hypothyroid mice with increasing doses of T3 to suppress circulating TSH. T3 was administered daily over the last 2 weeks of the 5-week protocol (Fig. 5D). T3 effectively suppressed TSH levels in both groups, with no detectable difference between them. Surprisingly, both quantitative PCR and reverse transcription-polymerase chain reaction data show no change in TRβ mRNA from microdissected PVN (Fig. 5E, F), despite genomic recombination. Therefore, these data suggest that MC4R neurons do not significantly express the Thrb gene.
Discussion
Regulation of TH levels is a complex multilevel phenomenon. 17,39 The PVN is a key regulator of TH levels, integrating multiple signaling systems. Here, we provide direct causal evidence of PVN TRH neuronal activity determining TH levels for the first time. We also demonstrate that ARC AgRP/NPY neurons are major regulators of the HPT axis and crucial mediators of the physiological response to a negative energy balance.
TRH is a potent stimulator of TSH synthesis, secretion, and bioactivity, ultimately being an important regulator of thyroid activity and TH levels. 4 –6 In this study, we demonstrate that the specific stimulation of TRH neurons in the PVN activates the whole HPT axis, including the expression of target genes in the liver. However, because Cre is expressed broadly in the PVN under the control of the TRH genomic locus, it is not clear which or how many of these neurons are required for this effect. In addition, work by a variety of groups has established that TRH neurons in the PVN co-express the MC4R and the NPY1 and 5 receptors as well TRβ isoforms. 23,40 –42 Still, it is not yet clear whether every TRH neuron that projects to the median eminence to regulate pituitary TSH levels has the capability to respond to all of these signaling pathways simultaneously.
It is well known that TRH mRNA is suppressed by TH and its expression increases in hypothyroidism. Nevertheless, during fasting, Trh expression remains low despite reduced TH levels. This suggests that TRH neurons have a primary role in the response to negative energy balance. Indeed, when activating TRH neurons in fasting mice, we were able to restore TSH and T4 levels. Hence, suppression of TRH neurons is a key regulatory step leading to the HPT axis suppression during fasting.
A possible explanation for suppressed TRH in face of reduced circulating TH is increased hypothalamic synthesis of T3. It has been proposed that fasting increases hypothalamic type 2 deiodinase (Dio2) expression and activity, increasing hypothalamic T3 production despite reduced circulating TH levels. 43,44 However, fasting-induced suppression of TRH expression is preserved in TRβ2KO mice, 15 and fasting does not increase hypothalamic TH action, 35 likely dismissing the hypothesis of increased local synthesis of T3. Additionally, 5′deiodinase activity is not required for fasting-induced suppression of TH levels. 45
Previously, we demonstrated that NPY and the MC4R regulate the HPT axis such that the response to fasting is lost in mice that lack both the MC4R and NPY. 19,22 Strikingly, as we show here, chemogenetic activation of ARC AgRP/NPY neurons suppresses TSH and T4 levels, even though the animals showed increased food intake and body weight. 46 Since a positive energy balance does not inhibit the HPT axis, the direct action of AgRP/NPY neurons must be regulating the HPT axis through suppression of TRH neurons.
Since fasting activates AgRP/NPY neurons, we wanted to determine if inhibiting their activation during fasting would prevent the fall in TH levels seen during a fast by blocking their downstream effects on TRH neurons. Indeed, our results confirm this hypothesis, showing that inhibition of ARC AgRP/NPY neurons disrupts the normal response to fasting, preventing HPT axis suppression. Therefore, in a state of negative energy balance, it is likely that metabolic signals drive the activation of AgRP/NPY neurons in the ARC, which project to the PVN and suppress TRH neurons through MC4R and NPY receptors, inhibiting TSH secretion and reducing TH levels.
However, it is still not clear why PVN TRH neurons do not respond to the low TH levels seen in fasting as the presumption has always been that all hypophysiotropic TRH neurons express MC4R, NPY receptors, and the TRβ and thus should respond to all signals. Indeed, it is reasonable to propose that TRH neurons integrate this multitude of inputs, initiating a metabolic response through the regulation of the HPT axis. To test this, we aimed to disrupt TH signaling in melanocortin-responsive neurons. Since MC4R has been described to colocalize with TRH in the PVN, we generated MC4R-specific TRβKO mice (TRβKO MC4R ). Surprisingly, TRβ mRNA expression was not reduced in TRβKO MC4R mice in the PVN, and they also possessed a normal HPT axis at baseline and responded normally to both low and high levels of TH. This result indicates that MC4R-positive neurons may not express TRβ, suggesting that distinct subpopulations of hypophysiotropic TRH neurons respond to different stimuli (Fig. 6). Further studies are necessary to address this hypothesis, but undoubtedly, the data presented challenges the current paradigm of the HPT axis regulation.
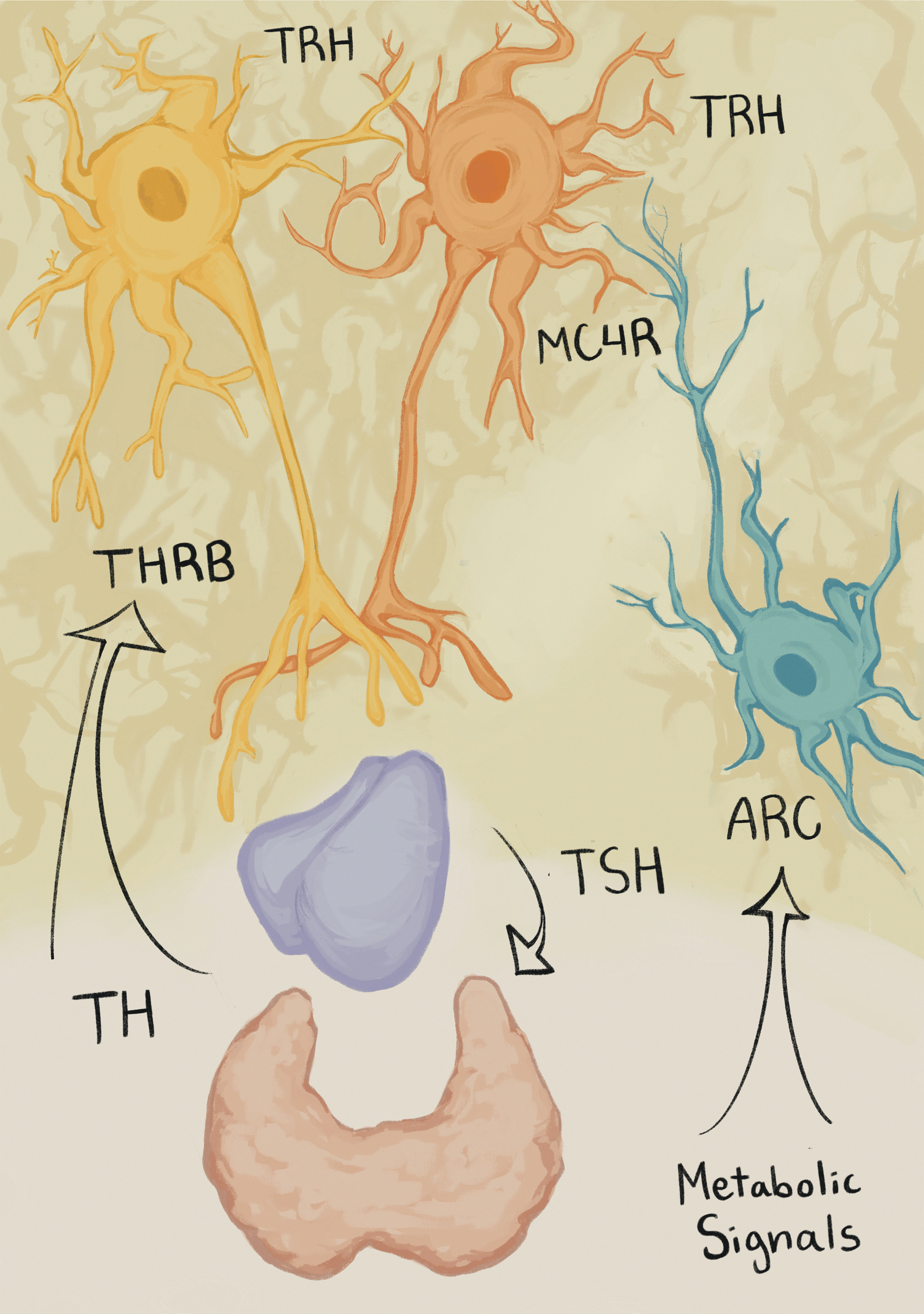
Diagrammatic description of proposed TRH regulation. The HPT axis is regulated by ARC neurons, and present data suggest that MC4R neurons do not significantly express THRB. Therefore, we hypothesize that TRH subpopulations are responsive to distinct stimuli. Here, two TRH neurons are depicted representing the possible existence of multiple TRH subpopulations. However, it is unclear if these are indeed distinct populations and, if so, how they affect each other. THRB, thyroid hormone receptor beta.
In summary, the present work shows a direct relationship between an ARC-PVN circuitry using chemogenetic techniques, and this circuitry is directly responsible for the HPT adaptive response to negative energy balance. Indeed, fasting-induced HPT axis suppression requires inhibition of TRH neurons and that is mediated, at least in part, through ARC AgRP/NPY neurons. It remains to be determined if the lack of response to low levels of TH by TRH neurons is because there are specific subsets of TRH neurons as our data suggest.
Footnotes
Acknowledgments
We thank Bradford Lowell (BIDMC/Harvard) for the Cre lineages (AgRP, MC4R, and TRH), Laura Alonso (WCM) for the mTmG mouse, and Karine Gauthier (ENSL) for the floxed TRβ mouse. The authors are also thankful for Lara Spitz Sousa artistic representation connecting key concepts of the present study in ![]() .
.
Authors' Contributions
R.H.C.-e-S.: Performed the experiments, analyzed the data, and wrote the article. R.R.: Performed the experiments and reviewed the article. A.N.H.: Conceived the study, and reviewed and edited the article. K.R.V.: Conceived the study, performed the experiments, analyzed the data, and reviewed the article.
Author Disclosure Statement
The authors have nothing to disclose.
Funding Information
This work was supported in part by K01 DK091403 and RO1 grants DK056123 and DK117940 from the NIDDK and Research Grant from the American Thyroid Association to K.R.V. R.R. was supported by a research fellowship 2014/20704-5 from FAPESP (Brazil).
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3