
Abstract
Background:
The density of tumor-associated macrophages in the tumor microenvironment of anaplastic thyroid cancer (ATC) is associated with poor prognosis. However, the crosstalk between macrophages and ATC cells is poorly understood. This study aimed to examine the impact of macrophages on cancer cell phenotypes. We found a new mediator between M2 macrophages and ATC cells through proteomics analysis.
Methods:
The role of macrophages in proliferation, migration, and invasion of ATC cells was evaluated using coculture assay and conditioned medium (CM). Secretory factors in the CM from single or coculture were identified using liquid chromatography-tandem mass spectrometry proteomics analysis. We evaluated the role of the secretory factor in proliferation, migration, and invasion of cancer cells. In vivo xenograft model was used to evaluate the effect of the factor.
Results:
M2 macrophages significantly increased the proliferation, migration, and invasion of ATC cells, whereas M1 macrophages decreased the proliferation, migration, and invasion of ATC cells. Based on proteomic analysis of CM, we identify carboxypeptidase A4 (CPA4) as a mediator of the crosstalk between macrophages and ATC cells. CPA4 was only detected in the coculture media of M2 macrophage/8505C, and its expression in cancer cells increased by M2 macrophage. The expression of CPA4 protein was significantly higher in human thyroid cancers, particularly in ATCs, than normal and benign tissues. A bioinformatics analysis of public data revealed that CPA4 expression was associated with poor prognosis and dedifferentiation of thyroid cancer. Knockdown of CPA4 suppressed proliferation, colony formation, migration, and invasion of ATC cells, consistent with the decrease of STAT3, ERK, and AKT/mTOR phosphorylation and epithelial-mesenchymal transition (EMT) marker expression. In addition, the increased expression of CPA4 in cancer cells by M2 macrophage stimulation induced the polarization of macrophages to the M2 phenotype, which formed a positive feedback loop. Xenograft tumors did not develop after CPA4 knockdown.
Conclusions:
Our data suggest that CPA4 stimulates the progression of thyroid cancer by mediating between M2 macrophages and ATC cells. CPA4 can be a new therapeutic target for the treatment of patients with ATC.
Introduction
Anaplastic thyroid cancer (ATC), an undifferentiated aggressive cancer, accounts for 1–2% of all thyroid cancer diagnoses. 1,2 Multimodal treatments including surgery, radiation therapy, and cytotoxic chemotherapy are less responsive, which lead to short overall survival, with 6 months median survival of ATC. 1,2 New targeted therapies and immunotherapy have been developed to treat ATC. 3,4 The combination of dabrafenib and tramentinb for BRAF-mutated ATC has received the Food and Drug Administration approval and has shown a good response. 2,5,6 The use of immune checkpoint inhibitors for ATC is at the clinical trial stage. 4,7 However, there is limited evidence to show that this treatment will improve the survival of patients with ATC. The discovery of novel targets based on molecular mechanisms is required to improve overall survival rates in patients with ATC.
The tumor microenvironment is a biological complex composed of various types of stromal cells and extracellular matrix. 8 Tumor-associated macrophages (TAMs) are the core constituent cells of cancer-infiltrating immune cells and play an important role in cancer formation and progression. 9 Macrophages are classically divided based on polarization states into M1 and M2 macrophages. 9,10 M1 macrophages are known to act as cancer suppressors by recognizing and killing cancer cells, whereas M2 macrophages are known to promote angiogenesis, cancer cell proliferation, and metastasis. Recent evidence suggests that the function and characteristics of TAMs are much more complex than this classical differentiation, 11 but M2-like TAMs are believed to have associations with tumor progression and resistance to therapies, which leads to poor survival. 11
Macrophage infiltration increases as the degree of differentiation worsens in thyroid cancer. 12 In ATC, TAMs represent more than 50% of nucleated cells. High TAM density in thyroid cancer is associated with increased invasion and decreased patient survival. 10,12 Therefore, studying the regulation of macrophage polarization and their effect on thyroid cancer progression will open new areas of therapy. In this study, we analyzed the relationship between macrophage polarization states and ATC cell progression and identified carboxypeptidase A4 (CPA4) as a mediator of the positive feedback loop between macrophages and thyroid cancer cells through proteomic analysis of conditioned medium (CM). In this study, we evaluated the role of CPA4 in ATC tumorigenesis and macrophage polarization using in vitro assays and xenogeraft mouse model. Furthermore, we demonstrated clinical significance of CPA4 using public database analysis and human thyroid tissues. To the best of our knowledge, this is the first study to identify CPA4 as a potential biomarker and immunotherapeutic target for ATC.
Materials and Methods
Cell culture and macrophage-cancer cell cocultures
Three different ATC cell lines (BHT101, 8505C, and SW1736) with the BRAF mutation and a human acute monocytic leukemia cell line (THP-1) were used for experiments. For the macrophage/thyroid cancer cell coculture, THP-1 cells (7.5 × 105 cells/well) were seeded in the upper insert of a 6-well transwell apparatus (0.4 μm pore size, Corning, NY, USA) and differentiated into different stages of macrophages. Inserts containing macrophages were transferred to a 6-well plate seeded with thyroid cancer cells (5 × 104 cells/well) and cocultured. After 48–72 h of coculture, macrophages or thyroid cancer cells were harvested for analysis. Lentivirus-mediated knockdown or overexpression of CPA4 and siRNA transfection was performed to evaluate the role of CPA4. Detailed information regarding in vitro analysis is provided as Supplementary File S1. The primers used for real-time PCR are shown in Supplementary Table S1.
LC-MS/MS proteomic analysis
CMs were collected and centrifuged to remove cell debris. Proteins were digested with trypsin/Lys-C, and salts were removed by C18 reverse phase chromatography. Liquid chromatography-high-resolution mass spectrometry data were processed using SequestHT on Proteome Discoverer (version 2.2, Thermo Fisher Scientific), as described. 13
Analysis of public transcriptome data
Transcriptome data and its matched clinical patient data from The Cancer Genome Atlas (TCGA-THCA) and GSE33630 were used to evaluate biological and clinical significance related to CPA4. We divided the tumors in TCGA-THCA data according to the CPA4 mRNA expression status (negative or positive) and then compared important clinical prognostic predictors in thyroid cancer such as extrathyroidal extension (ETE), lymph node metastasis (LNM), and disease-free survival (DFS). In addition, thyroid differentiation scores (TDSs) 14 were used to identify the association between CPA4 expression and dedifferentiation. Moreover, we performed gene set enrichment analysis (GSEA) using Hallmark or KEGG gene sets in tumors with CPA4 expression, compared with tumors without CPA4 expression. 15 Among the ranked gene sets from the GSEA, those with a nominal p value <0.05 and an FDR q-value < 0.25 were considered statistically significant.
Immunohistochemistry staining
Immunohistochemical staining was performed as described. 16 The tissue microarray (TMA) section includes 135 normal thyroid, 39 benign (nodular hyperplasia or follicular adenoma), 138 papillary thyroid cancer (PTC), and 21 ATC tissues. This study was conducted according to the guidelines of the Declaration of Helsinki. The use of TMA was approved by Institutional Review Board of Asan Medical Center, Seoul, Korea (IRB No. 2020-1219). Informed consent was obtained from all participants involved in the study.
Anti-CPA4 (Abcam, Cambridge, UK) and anti-CD163 (Abcam, Cambridge, UK) antibodies were used. The combinative staining results of CPA4 and CD163 were evaluated semi-quantitatively by an experienced pathologist (D.E.S): 0 (no staining), weak (<10% positive), moderate (10%–50% positive), and strong (>50% positive) expression; moderate-to-strong expression was considered positive for protein expression.
Animal experiments
Protocols for the animal experiment were reviewed and approved by the Institutional Animal Care and Use Committee of Asan Institute for Life Sciences (Project Number: 2022-12-132). For the xenograft experiment, 8505C cells were resuspended at 5 × 106 cells/200 μL in matrigel and phosphate-buffered saline and injected into female BALB/c nude mice.
Statistical analysis
Data are represented as mean ± standard deviation (SD). SPSS version 23.0 or GraphPad Prism software (San Diego, CA) was used to compare statistical differences. Continuous and categorical variables were compared by Student’s t-test and chi-square test, respectively. DFS was analyzed between negative and positive CPA4 tumor groups using a Kaplan–Meier estimator. Cochran–Armitage test was used for trend analysis. All p values were two-sided, and p < 0.05 was considered statistically significant.
Results
M2 macrophages stimulate proliferation, migration, and invasion of ATC cells
THP-1 monocytes were polarized into M1 and M2 macrophages. M1 macrophages showed higher CXCL11 and NOS2 expression (Supplementary Fig. S1A, B), and M2 macrophages showed higher CCL13 and MRC1 expression (Supplementary Fig. S1C, D). These data confirm the polarization of monocytes to macrophage subtypes.
To characterize the effect of macrophage subtypes on ATC cell progression, differentiated macrophages were cocultured with ATC cells. Coculture of M2 macrophages significantly increased the viability of 8505C and SW1736 cells compared with that of the control groups. Coculture with M1 macrophages significantly decreased the proliferation of 8505C and SW1736 cells (Fig. 1A, B). We evaluated the protein levels of key signaling pathways and EMT markers. The protein expression of p-AKT and N-cadherin increased in 8505C cells cocultured with M2 macrophages compared with the control (Fig. 1C and Supplementary Fig. S2A). The protein expression of p-AKT, p-ERK, and ZEB1 was decreased in 8505C cells cocultured with M1 macrophages, whereas the protein expression of p-AKT, ZEB1, and Slug increased in SW1736 cells cocultured with M2 macrophages (Supplementary Fig. S2B, C).
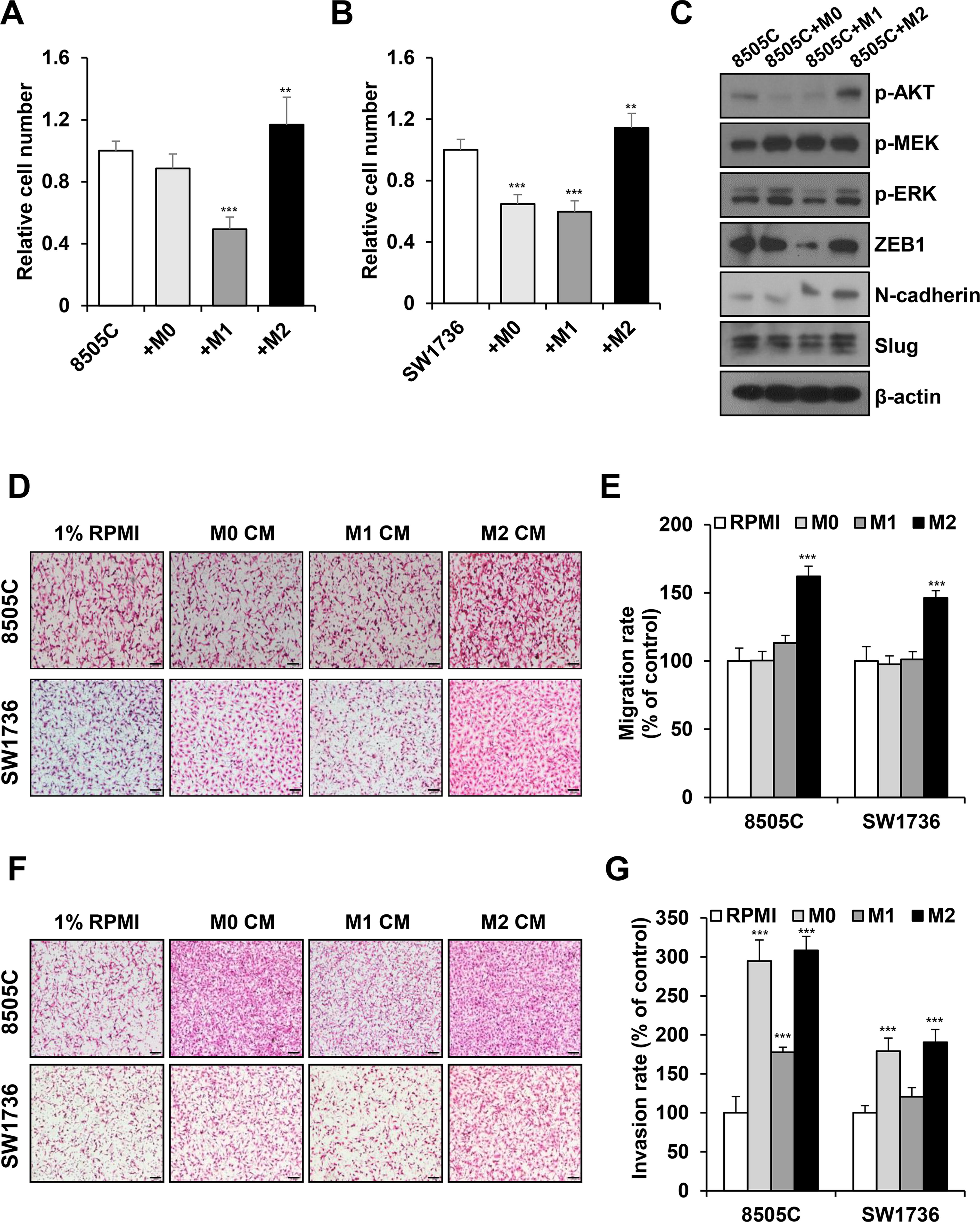
M2 macrophages induce proliferation, migration, and invasion of anaplastic thyroid cancer ATC cells.
When we treated ATC cells with macrophage CM, M2 macrophage CM increased the number of migrated 8505C and SW1736 cells (Fig. 1D, E). M2 macrophage CM significantly increased cancer cell invasion; by contrast, 8505C and SW1736 cells treated with M1 macrophages exhibited decreased invasion ability (Fig. 1F, G). Our data indicate that M2 macrophages significantly induce ATC cell proliferation, migration, and invasion in vitro.
CPA4 expression is upregulated in M2 macrophage-thyroid cancer cell coculture
We hypothesized that secreted molecules in CM mediate crosstalk between M2 macrophages and thyroid cancer cells to induce proliferation, migration, and invasion. To identify the secretory factors, we analyzed CM from each condition: 8505C cancer cells, each subtype of polarized macrophages, M0-8505C coculture, M1-8505C coculture, and M2-8505C coculture (Supplementary Fig. S3A). Secretome analysis revealed that 32 proteins were specifically detected in M2/8505C coculture CM compared with those in 8505C, M0-8505C, and M1-8505C (Fig. 2A and Supplementary Table S2). When M0, M1, M2, M0-8505C, M1-8505C, and M2-8505C were compared, 15 proteins were specifically detected in M2/8505C coculture CM (Supplementary Fig. S3B and Supplementary Table S3). Based on the literature review, we selected five secreted proteins, of which three (IGFBP2, CPA4, and TFP1) were expressed only in M2/8505C coculture and two proteins (FBN2, IGFBP1) were upregulated in M2/8505C coculture compared with M0/8505C and M1/8505C cocultures. CPA4 showed the highest increase in mRNA expression in M2/8505C cells cocultures than in M0/8505C or M1/8505C cocultures (Supplementary Fig. S3C-F). Therefore, we further focused on CPA4. Figure 2B shows the abundance results from the proteomic analysis of CPA4.
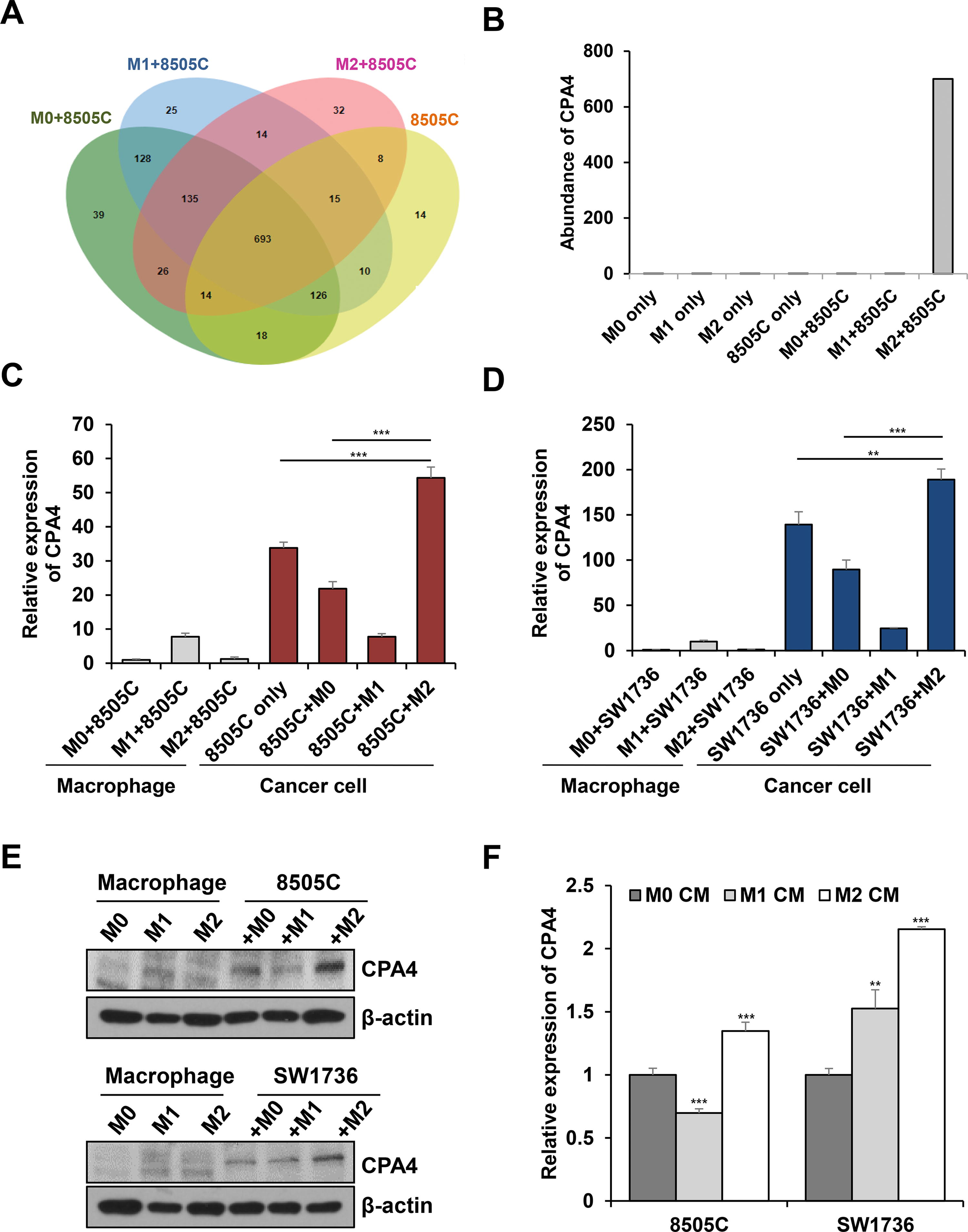
Secretome analysis of conditioned media revealed that CPA4 is specifically secreted under M2/8505C coculture conditions.
Moreover, we confirmed that CPA4 is derived from cancer cells using CPA4 mRNA expression analysis. The CPA4 expression increased in 8505C cells of the M2-8505C group compared with 8505C cells of the M0-8505C group or 8505C cells only group (Fig. 2C). Figure 2D also demonstrated high CPA4 expression in SW1736 cells from M2-SW1736 coculture, in contrast to SW1736 cells from either the M0-SW1736 group or the SW1736 only group. Western blot analysis revealed consistent results (Fig. 2E and Supplementary Fig. S4A). When cancer cells were treated with CMs from each polarized macrophage subtypes, CPA4 expression significantly increased in 8505C or SW1736 cancer cells treated with M2 macrophage CM (Fig. 2F). Because AKT was activated in cancer cells cocultured with M2 macrophages, we hypothesized that the AKT pathway regulates CPA4 expression in cancer cells. Treatment with the AKT inhibitor perifosine led to significant reduction in CPA4 expression in 8505C cells induced by M2 macrophage (Supplementary Fig. S4B, C). Taken together, thyroid cancer cells stimulated by M2 macrophage specifically secrete CPA4 by AKT activation.
CPA4 is associated with dedifferentiation and poor clinical outcomes in thyroid cancer
We evaluated the clinical significance and tumor biology of CPA4 expression by analyzing public datasets. Supplementary Table S4 depicts the baseline clinicopathological characteristics of the subjects analyzed. Of 505 differentiated thyroid cancers, 387 (76.6%) showed CPA4 mRNA expression (0.11 ± 0.26). CPA4 mRNA expression was significantly elevated during dedifferentiation of human thyroid cancer (Fig. 3A), which was supported by lower TDS 14 in tumors with CPA4 expression, compared with CPA4-negative tumors (Fig. 3B). ATC had markedly higher CPA4 expression than normal thyroid and PTC (Fig. 3C). In GSEA analysis, CPA4-positive tumors possessed the fundamental cellular processes that are required for cancer progression, such as cell proliferation (e.g., cell cycle, DNA replication, and E2F targets), invasion (EMT, cell adhesion molecules, and axon guidance), and tumor inflammation (TNFα signaling, JAK-STAT3) (Fig. 3D, E). Moreover, the CPA4-positive tumors exhibited more aggressive and invasive features (Fig. 3F, G) and shorter DFS of patients (Fig. 3H).
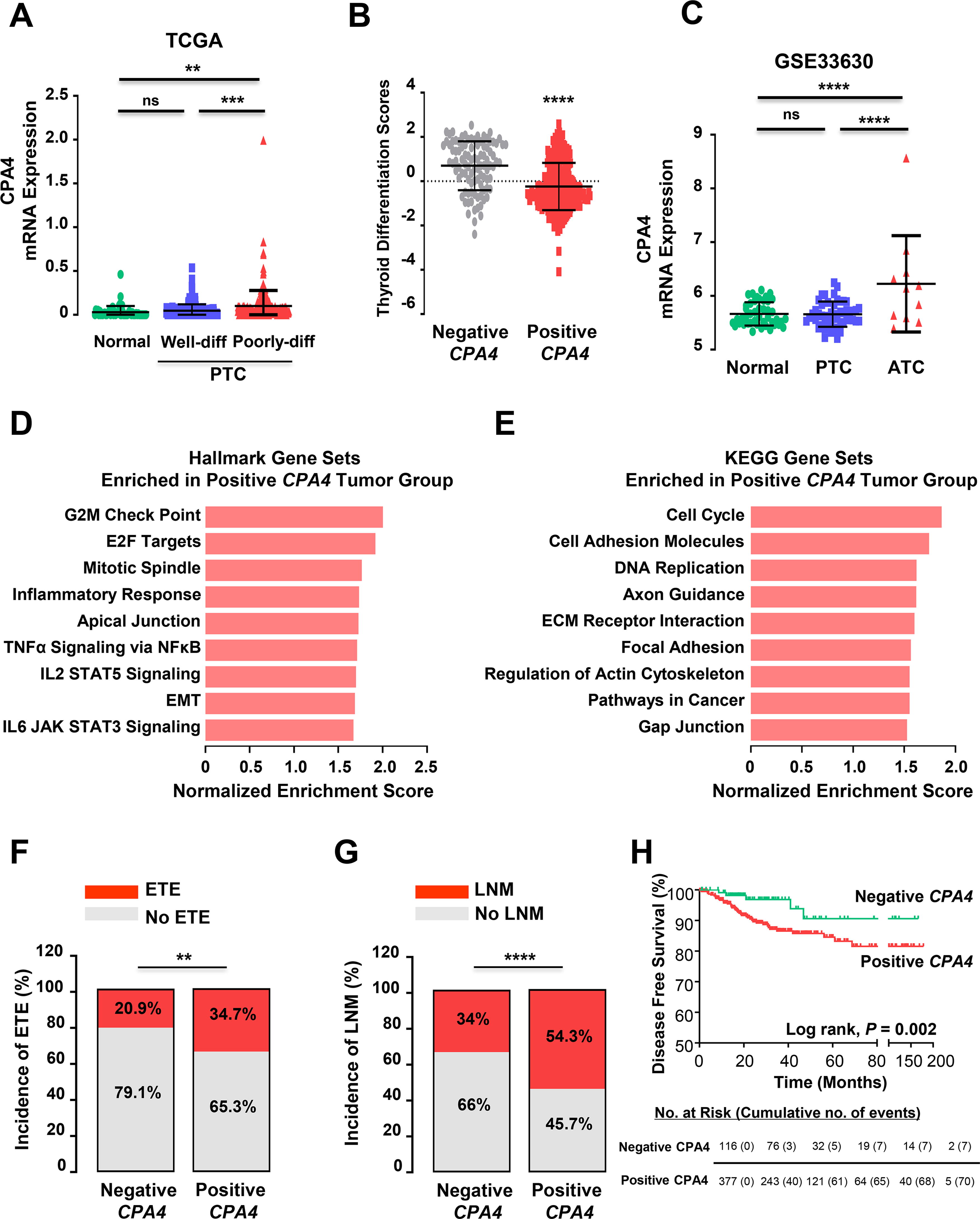
CPA4 expression contributes to poor clinical outcomes in thyroid cancer.
CPA4 knockdown inhibits proliferation, migration, and invasion of ATC cells
Next, we generated CPA4 knockdown 8505C and SW1736 cells. RT-PCR and western blot analysis revealed that siRNA treatment significantly reduced CPA4 expression (Fig. 4A, B). CPA4 suppression inhibited the growth of 8505C and SW1736 cells as well as their colony-forming ability (Fig. 4C, D). These results were confirmed using another siRNA in a low concentration (Supplementary Fig. S5A-C). Moreover, we established stable CPA4 knockdown (8505C and SW1736 cells) or overexpression (8505C and BHT101 cells). ATC cell proliferation significantly decreased in CPA4 stable knockdown cells compared with controls (Supplementary Fig. S6A, B). CPA4 overexpression enhanced cell proliferation compared with controls (Supplementary Fig. S6C, D). The transwell migration and invasion ability of both 8505C and SW1736 cells significantly decreased in siCPA4 transfected cells compared with controls (Fig. 4E, F). These results were confirmed using another siRNA (Supplementary Fig. S5D). CPA4 stable knockdown also significantly decreased the migration of ATC cells compared with controls (Supplementary Fig. S6E, F). These results suggest an important role of CPA4 in thyroid cancer cell proliferation, migration, and invasion.
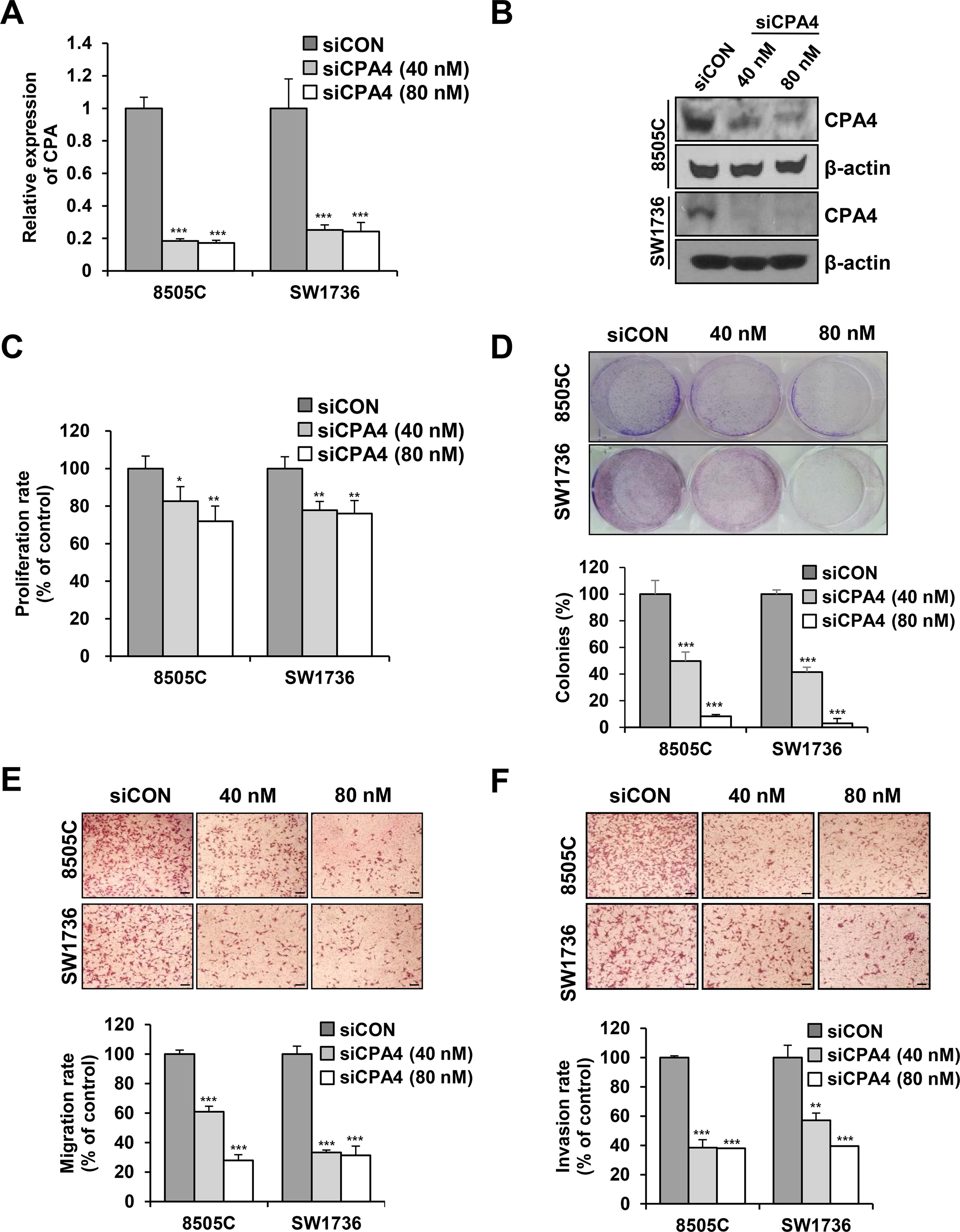
CPA4 knockdown inhibits proliferation, migration, and invasion of ATC cells.
CPA4 regulates EMT, STAT3, ERK, and AKT/mTOR signaling
Western blot analysis showed that CPA4 knockdown downregulated EMT markers such as N-cadherin and vimentin in 8505C cells (Fig. 5A and Supplementary Fig. S7A) and ZEB1, N-cadherin, vimentin, and Snail in SW1736 cells (Fig. 5B and Supplementary Fig. S7B). Conversely, CPA4 overexpression in 8505C cells resulted in the upregulation of ZEB1 and Snail (Fig. 5C and Supplementary Fig. S7C) and ZEB1, N-cadherin, vimentin, and Snail in BHT101 cells (Fig. 5D and Supplementary Fig. S7D). Furthermore, CPA4 knockdown significantly decreased STAT3, ERK, and AKT/mTOR phosphorylation in 8505C and SW1736 cells (Fig. 5A, B). CPA4 overexpression increased STAT3, ERK, and AKT/mTOR phosphorylation compared with controls in 8505C and BHT101 cells (Fig. 5C, D). We propose that CPA4 plays a role in thyroid cancer progression by activating the STAT3, ERK, and AKT/mTOR signaling pathways, as well as EMT.
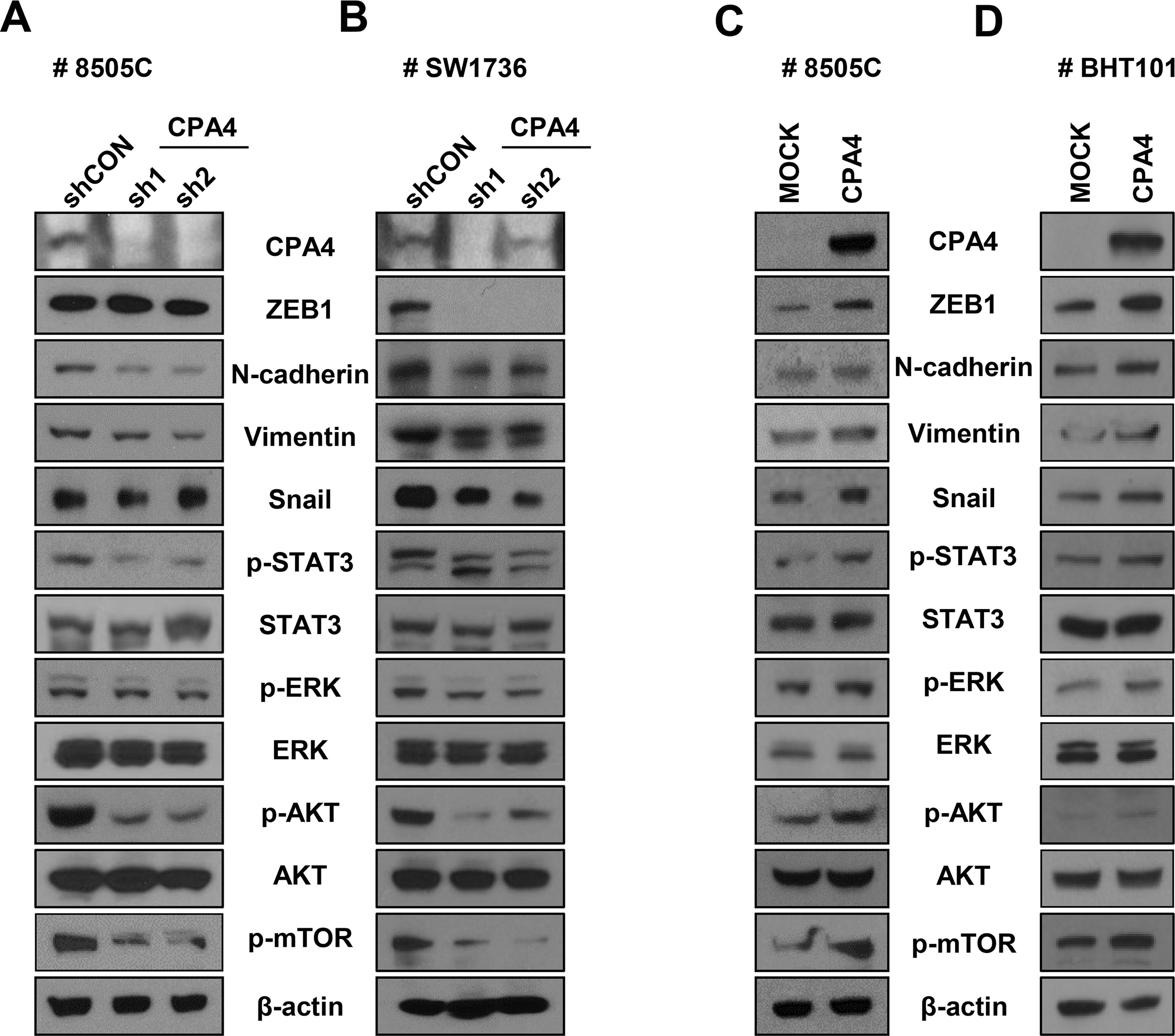
Lentiviral knockdown or overexpression of CPA4 changes signaling pathway and expression of EMT markers.
CPA4 induces M2 macrophage polarization
Next, we evaluated macrophage polarization according to CPA4 expression of ATC cells. M0 macrophages were cultured for 48 hours with CM from 8505C cells with CPA4 knockdown, overexpression, or control. The expression of M1 markers, such as IL-6 and CXCL11, significantly increased in macrophages treated with CPA4 knockdown cells CM (Fig. 6A). By contrast, IL-6 and CXCL11 expression significantly decreased in macrophages treated with CPA4 overexpression cells CM (Fig. 6B). Treatment with recombinant CPA4 protein also significantly decreased IL-6 expression and increased Arg1 and CCL13 expression (Supplementary Fig. S8). Protein expression of CD163 was higher in macrophages cultured with CPA4 overexpressing cells CM compared with mock CM. Conversely, the expression of CD163 significantly decreased in M2 macrophages treated with CPA4 knockdown cells CM (Fig. 6C). Flow cytometry analysis revealed that the percentage of CD163+ cells significantly decreased in macrophages cultured with CPA4 knockdown cells CM compared with the control (Fig. 6D). We found that STAT3 phosphorylation 17,18 increased in M2 macrophages cultured with CPA4 overexpressing cancer cells CM compared with control (Fig. 6C). Thus, CPA4 expression in cancer cells increased by M2 macrophages induced macrophage polarization to M2.
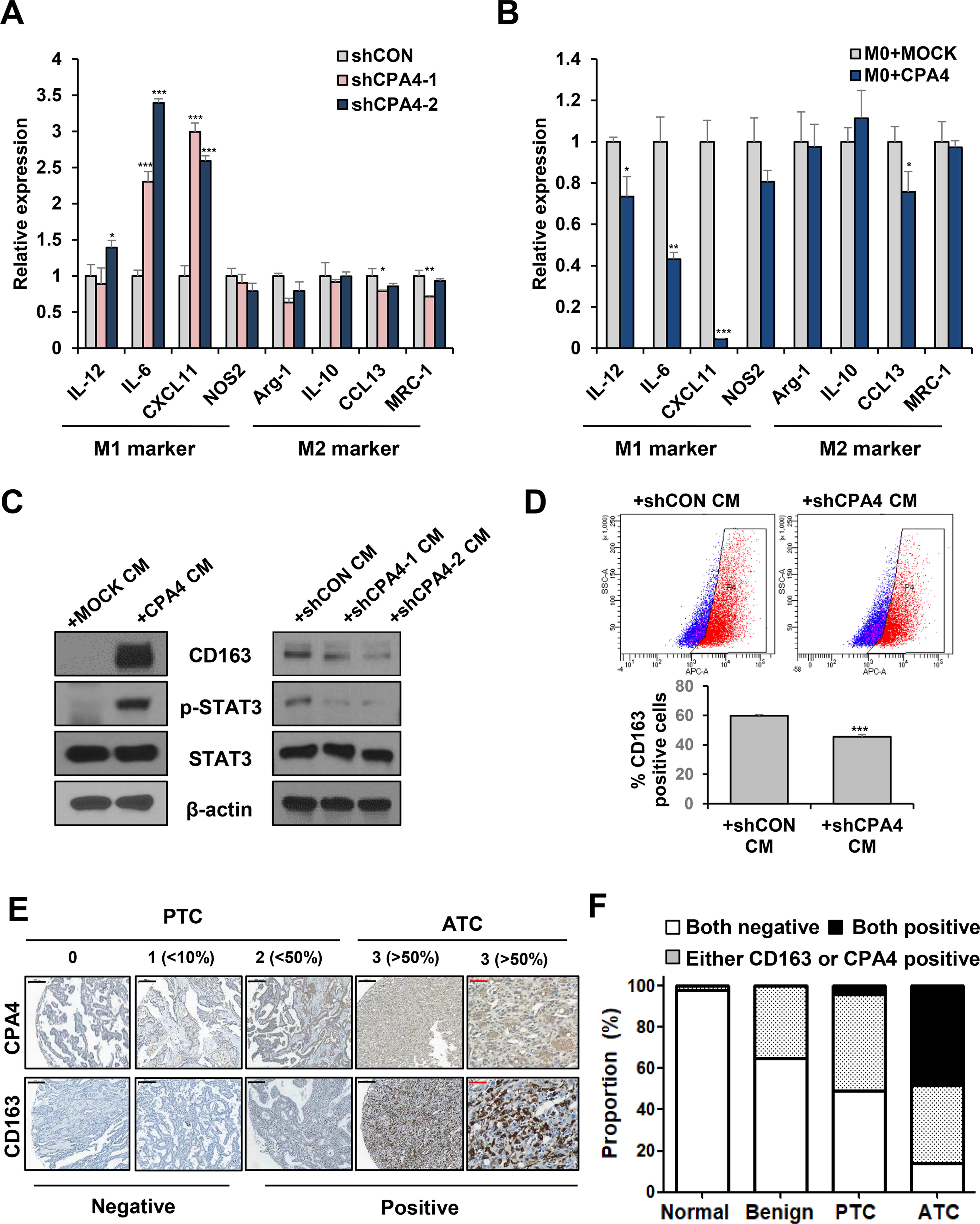
CPA4 regulates macrophage polarization.
We investigated the protein expression of CPA4 and CD163 in human thyroid tissues. The expression of CD163 was specifically observed in ATC tissues. CPA4 expression was significantly higher in human thyroid cancers, especially in ATC, than in normal and benign tissues (Supplementary Fig. S9 and Fig. 6E), consistent with the bioinformatics data. The positivity for CPA4 and CD163 protein expression was significantly correlated (Supplementary Table S5, p= 0.002), and both CD163 and CPA4 positive rates were the highest in ATC tissue (Fig. 6F). Collectively, these results indicate that CPA4 regulates the crosstalk between thyroid cancer cells and TAMs and is a potential therapeutic target in ATC.
Knockdown of CPA4 inhibits tumor growth in an in vivo 8505C xenograft model
To investigate the effect of CPA4 on tumorigenicity in vivo, we injected CPA knockdown and control 8505C cells into mice. On day 30 after injection, subcutaneous tumors were observed in the control group. However, tumor formation was not observed until the end of the experiment in the CPA4 knockdown group. Representative images of tumor size and tumor incidence are shown in Figure 7A. Tumor growth and weight in the control group are shown in Figure 7B and C.
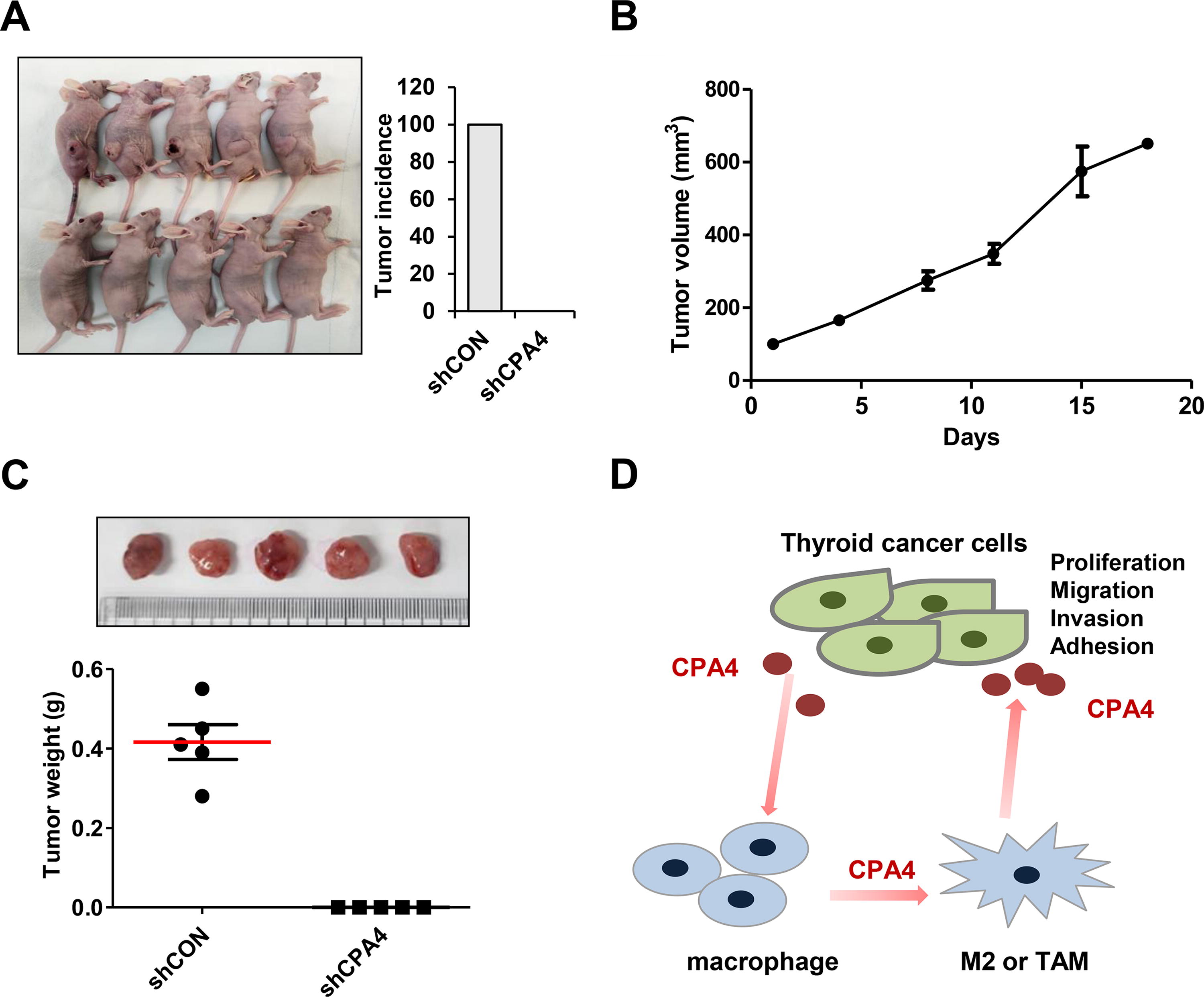
Knockdown of CPA4 inhibits tumor growth in the 8505C xenograft model.
Discussion
We herein confirmed the different roles of macrophages in ATC progression using a coculture model. M2 macrophages promoted proliferation and invasion in ATC cells. To understand how ATC cells and M2 macrophages communicate, we evaluated the secretome using CM by LC-MS/MS proteomics analysis. We discovered that CPA4 is specifically secreted in ATC cells cocultured with M2 macrophages. Moreover, M2 macrophages induced CPA4 expression in ATC cells. CPA4 regulates the proliferation, migration, and invasion of ATC cells and induces macrophage differentiation toward M2 subtype, leading to a positive feedback loop (Fig. 7D). Our results demonstrate the important role of CPA4 in ATC progression and indicate that CPA4 can be a therapeutic target by regulating crosstalk between ATC cells and macrophages.
TAMs from circulating monocytes or tissue-resident cells are the most abundant immune cells in the tumor microenvironment of most cancers. 8,12,19 However, the role of infiltrating macrophages in cancer is poorly understood. Macrophages in cancer tissues were first believed to exert anticancer effect through phagocytosis. However, subsequent studies revealed that TAMs have tumor-promoting function, and increased infiltration of TAMs decreased patient survival in many cancers. 11 Owing to this diversity and complexity in the function of TAMs, the binary classification of TAMs as having the M1/M2 phenotype has a limitation. However, using polarized macrophages for in vitro experiments was essential. In this study, M1 macrophages decreased the proliferation, migration, and invasion of ATC cells, whereas M2 macrophages accelerated the aggressiveness of ATC cells. These results are consistent with studies that showed poor prognosis of patients with thyroid cancer with high TAM infiltration. 12
Given the significant role of secretory factors in the crosstalk between ATC cells and macrophages, we analyzed the CM and identified secreted proteins. After a literature review, we selected IGFBP2, TFP1, FBN2, IGFBP1, and CPA4 as possible crosstalk mediators. IGFBP2 expression has been associated with the accumulation of M2 macrophages in pancreatic cancer tissues and the induction of polarization into M2 macrophages through increased IL-10 production in pancreatic cancer cells. 20 FBN2 expression has been associated with an increase in immunosuppressive cells in bladder cancer. 21 In this study, we selected CPA4 based on mRNA expression analysis. Other potential mediators must also be further analyzed.
CPA4 belongs to the carboxypeptidase family and catalyzes the release of carboxy-terminal amino acids. 22 Several studies have shown that CPA4 promotes the growth of colorectal and liver cancer cells. 23,24 Shao et al. reported that CPA4 promotes the migration and invasion of pancreatic cancer cells and regulates EMT markers. 25 However, the clinical significance and role of CPA4 in thyroid cancer have not been studied. This study found that CPA4 expression was associated with aggressive thyroid cancer subtypes, low TDS, higher incidence of ETE and LNM, and low DFS. This association of CPA4 with aggressiveness of thyroid cancer is consistent with studies in lung, triple-negative breast, bladder, and pancreatic cancer. 26 –29 Our experiments confirmed the functional role of CPA4 in promoting thyroid cancer proliferation, migration, and invasion.
How CPA4 promotes cancer progression remains unclear; however, CPA4 mediates the activation of multiple signaling pathways and EMT. CPA4 knockdown in ATC cells showed decreased protein expression of activated AKT, mTOR, and STAT3. CPA4 knockdown decreased the expression of ZEB1, N-cadherin, and vimentin, whereas CPA4 overexpression increased ZEB1 and Snail expression. With respect to ERK signaling, all cancer cells had BRAF mutation and showed ERK activation from the beginning. M2 coculturing could not further increase the expression of activated ERK in thyroid cancer cells, even though the expression of the activated ERK increased with CPA4 overexpression and decreased with CPA4 knockdown.
In addition to regulating signaling pathways in thyroid cancer, CPA4 regulates other cells in the cancer microenvironment as a secretory factor. Studies have reported that high levels of CPA4 in the serum are positively linked to the progression of lung cancer and pancreatic cancer. 26,29 Our data showed that CPA4 has an important role in macrophage differentiation. CM secreted from CPA4 knockdown cancer cells induced M0 macrophage toward M1 polarization, whereas CM from CPA4 overexpressing cancer cells inhibited M1 polarization. CPA4 protein expression was associated with CD163 protein expression in human thyroid tissues. Consistently, a positive association was observed between CPA4 expression and macrophage infiltration in bladder urothelial carcinoma. 28 Taken together, CPA4 plays an important role in ATC progression by influencing the behavior of cancer cells and TAMs.
In conclusion, CPA4 appears to simulate thyroid cancer progression by mediating the crosstalk between macrophages and ATC cells. CPA4 expression was significantly higher in ATCs than in differentiated thyroid cancers and associated with poor clinical outcomes. Thus, CPA4 may be a novel immunotherapeutic target for ATC.
Footnotes
Author’s Contributions
Y.-S.C. and M.J.J.: Conceptualization, data acquisition, and analysis, writing original draft, review, and editing. W.K.L.D., D.E.S., and K.K.: data acquisition and analysis, article review and editing. W.B.K. and W.G.K.: Conceptualization, interpretation of the data, article review and editing, funding acquisition, and administration. All the authors had full access to the data, took responsibility for the accuracy of the data analysis, and approved the final version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the National Research Foundation (NRF) of Korea Research Grant (
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Figure S9
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary File S1