
Abstract
The importance of Dermacentor spp. in the transmission of tick-borne pathogens is not well recognized in Europe. To investigate the role of Dermacentor spp. in the transmission of tick-borne pathogens, questing ticks were collected in 9 sites from southern to northwestern France (Camargue Delta to Eastern Brittany) where Dermacentor spp. exist and tick-borne diseases had occurred previously. Three tick species were collected during the spring and autumn of 2009. Collected ticks (both males and females) included D. marginatus (n=377), D. reticulatus (n=74), and I. ricinus (n=45). All ticks were analyzed by PCR or reverse line blot for the presence of pathogens' DNA. Pathogens analyzed were based on veterinarian reports and included Anaplasma phagocytophilum, Coxiella burnetii, Anaplasma marginale, Borrelia burgdorferi, Bartonella spp., Babesia spp., Theileria spp., and Francisella sp. Francisella tularensis was not detected in any of the analyzed ticks. In D. marginatus, infection prevalence for A. phagocytophilum (3%) was similar to that found in I. ricinus in Europe. Other pathogens present in D. marginatus included A. marginale (0.5%), Bartonella spp. (9%), C. burnetii (12%), F. philomiragia (1.3%), and Theileria annulata/Babesia bovis (0.3%), which were detected for the first time in France. Pathogens detected in D. reticulatus included A. marginale (1%), Bartonella spp. (12%), C. burnetii (16%), Borrelia spp. (1.5%), and F. philomiragia (19%). Pathogens detected in I. ricinus included A. phagocytophilum (41%), Bartonella spp. (9%), C. burnetii (18%), A. marginale (1%), Borrelia spp. (4.5%), and Babesia sp. (7%). This study represents the first epidemiological approach to characterize tick-borne pathogens infecting Dermacentor spp. in France and that may be transmitted by ticks from this genus. Further experiments using experimental infections and transmission may be now conducted to analyze vector competency of Dermacentor spp. for these pathogens and to validate such hypothesis.
Introduction
The most widespread and abundant tick species in Europe is Ixodes ricinus. This ectoparasite is implicated in the transmission of several pathogens including Borrelia burgdorferi sensu lato (s.l.) (Smith and Takkinen 2006), Anaplasma phagocytophilum (Matsumoto et al. 2006, Woldehiwet 2010), Rickettia spp. (Socolovschi et al. 2009), Babesia spp. (Chauvin et al. 2009), Francisella tularensis (Foley and Nieto 2010), potentially Bartonella spp. (Cotté et al. 2008, Reis et al. 2011a), as well as some viruses such as tick-borne encephalitis virus (Kollaritsch et al. 2011).
The second most abundant tick species in Europe belongs to the genus Dermacentor, and is also important for public and veterinary health (Pérez-Eid 2007). Compared with I. ricinus, little data exist about the role of Dermacentor spp. in the transmission of pathogens in Europe. Dermacentor spp. are 3-host ticks (larvae, nymphs, and adults feed on different hosts, completing the life cycle in approximately 1 year) feeding on animals and accidentally on humans (Estrada-Peña and Jongejan 1999). Contrary to I. ricinus, Dermacentor larvae and nymphs are endophilic, i.e., they live in rodents and other micro-mammals burrows, thus limiting the contact with these stages (Pérez-Eid 2007). Also different from I. ricinus, Dermacentor males are partial bloodsuckers, with implications in the transmission of tick-borne pathogens (Pérez-Eid 2007). In France, spring and autumn are the main periods of activity for Dermacentor spp. ticks.
Two Dermacentor spp. are present in France—D. marginatus and D. reticulatus. D. marginatus infests ungulates whereas D. reticulatus feeds on dogs and horses; both species can bite humans (Estrada-Peña and Jongejan 1999). For example, Dermacentor spp. ticks accounted for 10% of the total number of ticks collected on humans in Spain (Estrada-Peña and Jongejan 1999), 0.9% in Italy (Manfredi et al. 1999), 3.25% in Turkey (Bursali et al. 2010), and 3.3% in Romania (Briciu et al. 2011). Dermacentor spp. are implicated in the transmission of Anaplasma ovis to sheep and goats (Crosbie et al. 1997, Friedhoff 1997), Babesia caballi and Theileria equi to horses (Kumar et al. 2009), Babesia canis to dogs (Cardoso et al. 2010), and Rickettsia slovaca to humans (Raoult et al. 2002). In addition, Dermacentor spp. are also suspected of transmitting several other pathogens, such as B. burgdorferi, F. tularensis, Coxiella burnetii, Rickettsia conori, and some viruses (Pérez-Eid 2007).
Ticks can harbor 2 or more infectious agents and effectively transmit them simultaneously (Swanson et al. 2006). Consequently, it is important to characterize the prevalence of pathogen co-infections in ticks, which is significant for the correct diagnosis and prophylaxis of tick-borne diseases. In Europe, few studies have characterized tick co-infection with several pathogens (Toledo et al. 2009a, Cotté et al. 2010, Halos et al. 2010, Reye et al. 2010, Reis et al. 2011b, Torina et al. 2010, Satta et al. 2011), and there is a need to conduct studies estimating the risk of infection for animal and human populations.
Bovine granulocytic anaplasmosis and tick-borne fever (TBF) of ruminants due to A. phagocytophilum has been diagnosed in autumn 2007 in 3 alpine areas where its main vector, I. ricinus, is absent or rarely found but Dermacentor spp. are abundant (unpublished results). On the basis of these results, we hypothesized that Dermacentor spp. are implicated in the transmission of A. phagocytophilum and other pathogens. To test this hypothesis, the present study was performed by collecting ticks in different sites across France to characterize the prevalence of tick-borne pathogens in Dermacentor spp. and sympatric I. ricinus ticks in relation to cattle pathologies reported in the selected areas.
Materials and Methods
Study areas and tick collection
This study was undertaken at locations included in the area known for the presence of Dermacentor spp. ticks in France (Perez-Eid, 2007). According to the indications of veterinary practitioners, sites were selected in pastures where previous TBF, babesiosis, Q fever, Lyme disease, or anaplasmosis outbreaks were diagnosed as acute or subacute diseases identified through clinical signs and/or confirmed by PCR or positive serology. Collection sites were chosen in 9 French departments, corresponding to 11 veterinarian practices, and are presented in Figure 1. Questing adult ticks were collected using the flagging technique (Vassallo et al. 2000) in the spring (April and May) of 2009 for sites 1–3 and 7–9, and in the autumn (September and October) of 2009 for sites 4 and 5. Flagging was conducted from 10:00 AM to 6:00 PM during 2 days. All adult ticks (male and female) were individually preserved in 70% ethanol and identified to the species level using taxonomic keys (Pérez-Eid 2007), categorized by site of collection and sex, and frozen at −20°C until DNA extraction.
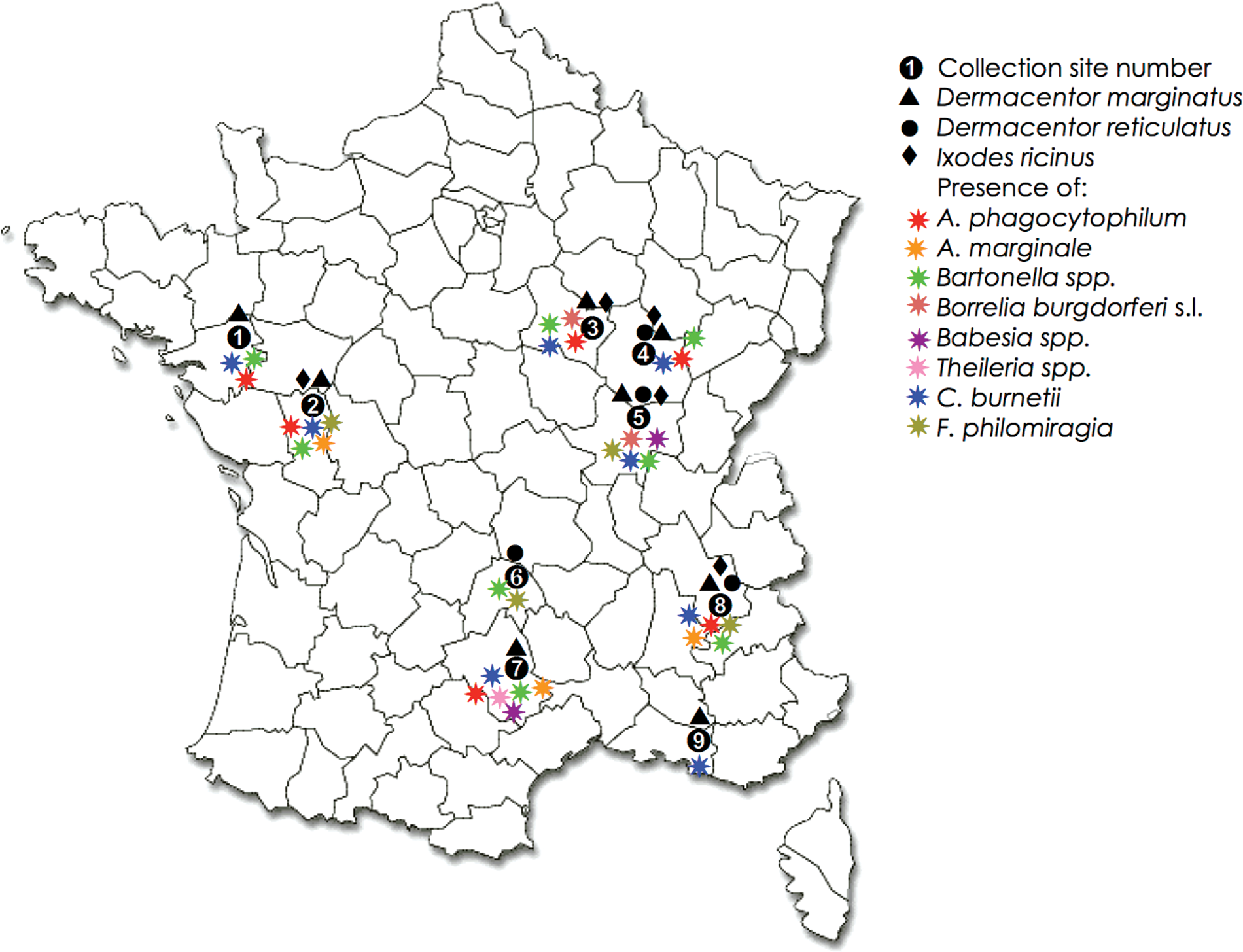
Tick collection sites in different French departments. Site 1: Loire Atlantique (department no. 44), Chateaubriand (47.24 N, 1.22 W), Soudan (47.44 N, 1.18W), Louisfer (47.4 N, 1.26 W); site 2: Deux-Sèvres (no. 79), St. Maurice la Fougeureuse (47.2 N, 0.3 W), St. Aubain du Plain (46.55 N, 0.28 W), Amailloux (46.44 N, 0.18 W), St. Julien de Vouvante (46.34 N, 0.46 W); site 3: Yonne (No. 89), St. Père (47.27 N, 3.45 E), Etaule (47.31 N, 3.55 E); site 4: Côte d'or (No. 21), Vic de Chassenay (47.28 N, 4.16 E), Chevigny (47.1 N, 5.28 E); site 5: Saône et Loire (no. 71), St Gervais/Couche (46.56 N, 4.56 E), Collonge la M (46.33 N, 4.47 E); site 6: Cantal (no. 15), Villedieu (44.59 N, 3.3 E); site 7: Aveyron (no. 12), Vezouillac (44.12 N, 3.5 E); site 8: Isère (no. 38), Nantes en Rattier (44.56 N, 5.49E), Notre Dame de Vaux (44.59N, 5.44E); site 9: Bouches du Rhône (no. 13), St Martin de Crau (43.38N, 4.48E). Color images available online at
DNA extraction
DNA was extracted from individual ticks using the QIAamp® DNA mini kit (QIAGEN, Germany) following manufacturer's instructions. Total DNA was eluted in 100 μL of elution buffer. DNA samples were then distributed at −20°C in 4 96-well plates with 25 μL of DNA per plate to the various laboratories for pathogen DNA characterization.
Pathogen DNA characterization
Pathogens' DNA was characterized by PCR or reverse line blot (RLB) in tick samples using specific primers (Tables 1 and 2). All of the methodologies used here were highly specific for the target pathogen except the PCR performed to detect Bartonella sp. and Borrelia sp., for which we cannot exclude a cross-reaction with some tick symbiont DNA. In these cases, a sequencing step was performed, when possible, for positive PCR reactions.
Laboratories: (1) USC INRA Bartonella-Tiques, Maisons-Alfort, France; (2) Laboratoire d'Analyses Sèvres Atlantique, Niort, France; (3) IREC, Ciudad Real, Spain; (4) IZSS, Palermo, Sicily, Italy; (5) Laboratoire National de Référence Francisella, ANSES, Maisons-Alfort, France.
Bartonella spp., Borrelia burgdorferi s.l., and Anaplasma marginale
For B. burgdorferi s.l., and Bartonella spp., PCR reactions were performed in the MyCycler thermocycler (Bio-Rad, Strasbourg, France). Each reaction was carried out in a 25-μL volume containing 2 μL of tick DNA, 2 μL of 10 μmol/L of each primer, 2 μL of 2.5 mmol/L of each deoxyribonucleotide triphosphate (dNTP), 2.5 μL of 10×PCR buffer, and 1 U of Taq DNA polymerase (5 U/μL Takara Biomedical Group, Shiga, Japan). PCR products were sent for sequencing to GATC Biotech Company (Germany). Sequences were compared with known sequences listed in the GenBank nucleotide sequence databases by using the BLAST search option at the National Center for Biotechnology Information (
A. phagocytophilum
For A. phagocytophilum, pathogen detection was conducted by real-time PCR using the commercial kit ADIAVET® ANA PHA REALTIME (Adiagène, St. Brieuc France) targeting the msp4 gene. PCR amplification was carried out with 2 μL DNA in a total volume of 25 μL in a thermocycler CFX 96 (BioRad). Three A. phagocytophilum biovars could be detected with this kit, namely biovar phagocytophilum, biovar equi, and biovar EGH. Negative (DNase- and RNase-free sterile water) and positive controls were included in all experiments. For Borrelia spp., positive control DNA was kindly provided by E. Ferquel (CNR Borrelia, Institut Pasteur, Paris, France) and for A. phagocytophilum, positive control DNA included in the ADIAVET® ANA PHA REALTIME kit was used. Amplicons were analyzed by electrophoresis in 1.0% or 1.5% agarose gels containing ethidium bromide and DNA fragments were observed under ultraviolet light.
Francisella sp
Real-time PCR assays were performed using primers and probes that target the tul4, fopA, and ISFtu2 genes of F. tularensis, as previously described (Hollis et al. 1989, Versage et al. 2003). The fluorogenic hybridization probes were synthesized by Applied Biosystems (France) with a 6-carboxy-fluorescein reporter molecule (FAM) attached to the 5′ end and a quencher (tetramethylrhodamine, TAMRA) attached to the 3′ end. Amplification and data analysis were carried out on an ABIPRISM 7000 (Applied Biosystems) thermocycler. Real-time PCR was performed on a 20-μL final volume using TaqMan Universal PCR Master-Mix, 0.4 μM forward and reverse primers, 0.1 μM fluorogenic probe, and 5 μL of template DNA. For each reaction, both negative (no DNA template and Escherichia coli DNA) and positive (2 pg of purified F. tularensis subsp. holarctica FSC 200 strain) controls were included. To verify if amplified products were the correct size, amplification products were run on 2% agarose gels and visualized by staining with ethidium bromide. This assay is species specific and able to differentiate F. tularensis and F. philomiragia. Identification of F. tularensis occurs when all 3 target sequences (ISFtu2, fopA, and tul4) give a positive result, whereas identification of F. philomiragia occurs when the ISFtu2 assay is positive and the tul4 assay is negative (Versage et al. 2003).
C. burnetii
C. burnetii was detected using the ADIAVET® COX REALTIME kit (Adiagène, St Brieuc, France). PCR amplification was conducted in a 25-μL volume containing 2 μL of tick DNA using a CFX 96 Thermocycler (BioRad). A control DNA included in the PCR kit was used as positive control, and DNase- and RNase-free water was used as negative control.
Babesia and Theileria spp
PCR amplifications were performed to amplify the hypervariable V4 region of the 18S rRNA gene of Babesia and Theileria species (Nagore et al. 2004). Reactions were carried out in 50 μL with 5 μL of tick DNA using a thermocycler 2720 (Applied Biosystems). PCR products were then used for RLB hybridization, as previously described (Gubbels et al. 1999, Georges et al. 2001, Schnittger et al. 2004). For each piroplasm, specific oligonucleotide probes were used (Table 2) to detect Babesia/Theileria spp., Babesia bigemina, B. bovis, B. divergens, B. major, B. motasi, B. ovis, B. crassa, Theileria annulata, T. velifera, T. taurotrago, T. mutans, T. buffeli/orientalis, T. ovis, T. lestoquardi, Theileria all sp2 (China), Theileria all sp1 (China), Babesia all sp1 (Turchey), and Babesia all sp2 (Lintan). After hybridization, the membrane was exposed to a chemiluminescent detection film (Amersham) for 60 min to 24 h and then developed on Develop X-ray film (AGFA) and Fixed X-ray film (AGFA). A black spot in the sample–probe cross in the hyperfilm demonstrated a positive signal for that pathogen.
Statistical analysis
A 2×2 chi-squared test or Fisher exact test (when n<10) was performed using the SPSS 11.0 statistical program (SPSS Inc., Chicago, IL) to compare prevalence between tick species for a given pathogen or between pathogens for a given tick species. The differences were considered statistically significant at p≤0.05.
Results
Tick collection
A total of 495 adult ticks (60% females and 40% males) were collected (Table 3). Three species of ticks were identified on different collection sites: D. marginatus (n=377; 76%), D. reticulatus (n=74; 15%), and I. ricinus (n=45; 9%) (Fig. 1). Most ticks (57%; n=284) were collected in western France in collection site 2 (department no. 79). Despite the importance of this sample, D. reticulatus was not collected in this site. A similar result was obtained for site 3 (n=11). The second site in terms of the number of ticks collected was site 8 (n=62) in southeastern France where the 3 tick species were recovered in sites 4 and 5, in spite of the small number of ticks collected in these 2 sites (n=6 and n=17 ticks, respectively). In sites 1 (n=60), 7 (n=38), and 9 (n=9), only D. marginatus specimens were found, whereas in site 6 (n=8) only D. reticulatus specimens were found.
Pathogen detection in ticks
Detection of Bartonella spp
Of the 495 tick samples tested, 47 (9.5%) were positive for the 356-bp fragment of the Bartonella spp. citrate synthase (gltA) gene (Table 4). The presence of Bartonella spp. was similarly distributed among the 3 tick species as well as between females and males (Table 4). Bartonella spp. were found in almost all collection sites, with the exception of site 9, where only 9 ticks were collected (Fig. 1). Eight sequenced amplicons were homologous to Bartonella spp. Four of them showed 100% identity with the uncultured Bartonella spp. isolate 10158 BART citrate synthase (gltA) gene (GenBank accession no. EF662055) that was isolated from Ixodes scapularis in the United States and for which the closest species is Bartonella rochalimae (76% identity). The other amplicons showed 97% identity with the uncultured Bartonella sp. clone 162 isolated from Ixodes tasmania in Australia (accession No. JQ228398), 76% identity with Bartonella melophagi strain K-2C (accession No. JQ228399), 77% identity with Bartonella sp. pn 1564ga isolated from a rodent in United States, the closest species being Bartonella grahamii (accession no. JQ228400), and 77% identity with Bartonella sp. B29044 that was isolated from bats in Guatemala and for which the closest known species is Bartonella elizabethae (accession No. JQ228401).
The number of positive/tested samples is shown for female and male ticks with percent prevalence shown for all (female + male) ticks. Infection prevalence were compared between host tick species for a given pathogen (significant differences compared against a D. reticulatus and b I. ricinus; p<0.05) and between pathogens in a given tick species (significant differences are shown with symbols i to viii for A. phagocytophilum, A. marginale, B. burgdorferi s.l., Bartonella spp., C. Burnetii, Babesia spp., Theileria spp., and F. philomiragia).
Collection sites are described in Figure 1.
The laboratories where the experiments were conducted are described in Table 1.
Detection of B. burgdorferi s.l
Of the 495 ticks analyzed, 3 (0.6%) were positive for the 357-bp amplified fragment of B. burgdorferi s.l 16S rDNA (Table 4). Two of them corresponded to I. ricinus females (collected on sites 3 and 5 in Eastern France) and 1 corresponded to a D. reticulatus male collected on site 5 (Fig. 1). Only 1 of the amplified fragments obtained from an I. ricinus, was sequenced and showed a 100% identity to B. burgdorferi strain Titov gaj 16S rDNA gene that was isolated from I. ricinus in Serbia (accession No. JQ228402).
Detection of A. marginale
Only 0.6% prevalence was found for A. marginale in collected ticks, with 2 D. marginatus females collected on sites 2 and 7 and 1 D. reticulatus male from site 8 positive for pathogen DNA. A. marginale was not detected in I. ricinus ticks.
Detection of A. phagocytophilum
Of the 495 ticks analyzed, 30 (6%) were positive for the 130-bp fragment of A. phagocytophilum msp4 gene when tested by real-time PCR. A. phagocytophilum was found with similar prevalence in all collection sites, with the exception of sites 5, 6, and 9, where fewer ticks were collected (Table 3 and Fig. 1). None of the D. reticulatus collected were positive for A. phagocytophilum and only 3% of D. marginatus (5% males and 2% females) were positive for A. phagocytophilum, whereas 41% of I. ricinus collected (60% males and 31% females) were positive for A. phagocytophilum (Table 4).
Detection of F. tularensis and F. philomiragia
All tick samples tested were negative for tul4 and 19 of them were positive for fopA and ISFtu2, indicating the absence of F. tularensis and a prevalence of 3.8% for F. philomiragia. This bacterium was found in 4 different collection sites (Fig. 1). Only D. marginatus and D. reticulatus showed positive results for F. philomiragia, with a higher prevalence of 18.9 % in D. reticulatus, especially in females (33 % prevalence) (Table 4).
Detection of C. burnetii
C. burnetii showed a prevalence of 12.9% in analyzed ticks (Table 4). C. burnetii DNA was recovered from ticks at all collection sites, except from site 6 in central France, where only 8 ticks were collected (Fig. 1). The bacterial DNA was found both in males and females of the 3 tick species collected, with a higher prevalence of 18% in I. ricinus, followed by D. reticulatus (16%) and D. marginatus (12%) (Table 4).
Detection of Babesia/Theileria spp
Prevalences of 0.8% and 0.2% were found for Babesia and Theileria spp. parasites, respectively (Table 4). Three I. ricinus female ticks collected on site 5 were positive for Babesia spp., with 2 of them positive for B. divergens and 1 that did not correspond to any of the Babesia spp. analyzed (i.e., B. bovis, B. divergens, B. major, B. motasi, B. ovis, and B. crassa). One D. marginatus female collected on site 2 was positive for Babesia and Theileria spp. that did not correspond to any of the Babesia spp. and Theileria spp. (T. annulata, T. velifera, T. taurotragi, T. mutans, T. hirsi, T. buffeli, T. ovis, and T. lestoquardi) analyzed. Finally, 1 D. marginatus female collected on site 7 was positive for both B. bovis and T. annulata. None of the samples from D. reticulatus were positive for Babesia or Theileria spp. (Table 4).
Co-infection with different pathogens
Among the 495 ticks tested, 153 (31%) were positive for at least 1 pathogen, 18 (12%) were positive for 2 pathogens, and none of them carried 3 or more pathogens. C. burnetii DNA was detected in association with all pathogens tested, except for B. burgdorferi s.l. and Theileria spp. Theileria spp. parasites were found only in a tick also positive for Babesia spp. Bartonella spp. DNA was found in association with all the pathogens tested with the exception of A. marginale, which was detected only together with C. burnetii in 1 tick. B. burgdorferi s.l. was found in 1 tick in association with Bartonella spp. and in another tick together with Babesia spp. Finally, F. philomiragia was detected in conjunction with C. burnetii in 4 ticks and with Bartonella spp. in 2 ticks.
Discussion
In this study, we report the results of a survey conducted in 9 study sites in France with the aim of evaluating the prevalence of tick-borne pathogens in Dermacentor spp. and sympatric I. ricinus ticks. We were interested in Dermacentor spp. because these ticks represent the second genus of medical and veterinary importance after Ixodes spp. in Europe; however, few studies have characterized pathogen prevalence in these ticks (Kahl et al. 1992, Sixl et al. 2003, Sting et al. 2004, de la Fuente et al. 2004a, de la Fuente et al. 2005a, Toledo et al. 2009a, Torina et al. 2010, de Carvalho et al. 2011, Satta et al. 2011).
Nine collection sites were chosen from southern to northwestern France where Dermacentor spp. are abundant and tick-borne diseases have occurred (unpublished results). D. marginatus was the most abundant tick species in collected samples, followed by D. reticulatus and I. ricinus. Dermacentor spp. ticks lack host specificity and could infest and transmit different pathogens during their life cycle to several vertebrate hosts, including humans (Estrada-Pena and Jongejan 1999). Therefore, it is important to investigate the prevalence of pathogens of medical and veterinary importance in these ticks. The choice of analyzed pathogens was made according to the pathologies reported by the veterinarians practitioners in the concerned zones and includes A. phagocytophilum, A. marginale, B. burgdorferi s.l., Bartonella spp., C. burnetti, Babesia spp., Theileria spp., and Francisella sp.
The most prevalent pathogen recovered in Dermacentor spp. was F. philomiragia in D. reticulatus, particularly in female ticks. Vector-borne transmission of F. philomiragia has never been suspected, and its detection, for the first time in ticks, was not initially planned in our study. While looking for F. tularensis, this bacterium was revealing in the ticks. This bacteria appears to be an opportunistic pathogen, primarily causing serious diseases associated with 2 risk groups of chronic granulomatous disease and immunocompromised patients (Hollis et al. 1989). F. philomiragia has been isolated from humans with a febrile syndrome compatible with bacterial infection in Europe, North America, and Australia (Hollis et al. 1989). Knowing whether this bacterium can be transmitted by a vector like a tick must now to be clarified. F. tularensis has been suspected to be transmitted by both mosquitoes and ticks (Eliasson et al. 2002, and 3 cases of transmission associated with Dermacentor spp. ticks have been described in Spain (Morner 1992, Alkorta et al. 2000, Teijo-Nunez et al. 2006). Furthermore, it was reported that 0.7% of the D. marginatus ticks analyzed in another area of Spain carried this pathogen (Toledo et al. 2009a). In Portugal, a Francisella-like endosymbiont with significant identity with F. tularensis was detected in 39% of the D. reticulatus analyzed (de Carvalho et al. 2011). Taken together, these results suggest that Dermacentor spp. ticks could play a role in the maintenance and transmission of Francisella spp.
C. burnetii was the second most prevalent pathogen recovered from all collection sites, with similar prevalence in all 3 tick species. C. burnetii is responsible for Q fever, a zoonotic disease endemic worldwide (Maurin and Raoult 1999). Goats are probably the main reservoir host, and humans become infected mainly by inhalation of contaminated aerosols or dusts containing C. burnetii shed by infected animals (Tissot-Dupont et al. 2004). However, although previously considered as negligible, the role of ticks in bacterial transmission to wildlife and pets and in maintaining C. burnetii in wild and peridomestic cycles is now clearly recognized (Toledo et al. 2009b). In addition, C. burnetii infects several tick species. Other authors have found PCR evidence of C. burnetii in Dermacentor spp. collected in Spain (Toledo et al. 2009b) and Germany (Beytout et al. 2007) and in Rhipicephalus spp. and Haemaphysalis spp. collected in Sardinia, Italy (Satta et al. 2011). On the other hand, C. burnetii was not detected in I. ricinus collected in The Netherlands (Sprong et al. 2012) nor in D. marginatus collected in Sardinia, Italy (Satta et al. 2011). These results suggested that several tick species might vector C. burnetii in different regions to wild and domestic animals and eventually humans.
Bartonella spp. DNA was detected with similar prevalence in the 3 tick species analyzed and in all collection sites except 1. Sequence results suggested the existence of new Bartonella spp. or strains and/or the amplification of DNA from an unknown endosymbiont as was previously reported (Tijsse-Klasen et al. 2011). The presence of Bartonella spp. has been reported in ticks from all over the world, including Europe (Angelakis et al. 2010a). However, the tick role in the transmission of Bartonella spp. has been debated for many years, despite several reports of indirect evidence (Billeter et al. 2008, Angelakis et al. 2010a, Telford and Wormser 2010). Recent studies demonstrated the transmission of Bartonella spp. by I. ricinus both in vitro (Cotté et al. 2008) and in vivo (Reis et al. 2011a). In Italy, Bartonella spp. were not detected in D. marginatus, whereas pathogen DNA was detected in Rhipicephalus spp. (Satta et al. 2011). Recently, a study reported the detection of B. henselae infection in a patient following a bite by a Dermacentor spp. tick that was infected with the same bacteria (Angelakis et al. 2010b). Bartonella spp. prevalence reported here in I. ricinus was similar to that reported in ticks collected form northern France (Halos et al. 2005) and higher than that reported in western France (0.2%) (Cotté et al. 2010) and near Paris (0.1%) (Reis et al. 2011b).
A high prevalence of A. phagocytophilum was found in I. ricinus ticks (41% by real-time PCR), whereas 3% prevalence was found in D. marginatus. This obligate intracellular bacterium is the causative agent of granulocytic anaplasmosis in several hosts, including humans, horses, dogs, and ruminants (Woldehiwet 2006). This pathogen is widely distributed in France, where it has been identified in 84 Departments (Matsumoto et al. 2006; unpublished results), beyond the limits of the presence of its main vector I. ricinus. This fact suggests the implication of other tick species such as Dermacentor spp. in the transmission of A. phagocytophilum. Some studies conducted in Spain reported the detection of A. phagocytophilum DNA in D. marginatus questing ticks (Toledo et al. 2009a) and in ticks feeding on deer and wild boar (de la Fuente et al. 2005a, de la Fuente et al. 2004a), when none was detected in studies conducted in Italy (Satta et al. 2011). In our study, D. reticulatus was not infected with A. phagocytophilum as previously reported in Austria (Sixl et al. 2003). As expected, I. ricinus was confirmed infected as the principal vector of A. phagocytophilum with prevalences that were higher than those reported from other studies conducted in different French regions (0.35–10.7%) (Parola et al. 1998, Cotté et al. 2010, Halos et al. 2010, Reis et al. 2011b). However, these results may be affected by differences in the experimental methods used to determine infection prevalence. In fact, the comparison of 2 detection methods used in our study to detect A. phagocytophilum has shown that real-time PCR had a better sensitivity than PCR (data not shown).
Some A. marginale-positive samples were detected in both D. marginatus and D. reticulatus, but not in I. ricinus, suggesting that Dermacentor spp. may be vectors of A. marginale in France. This bacterium, responsible for bovine anaplasmosis, is suspected to be transmitted by several hard tick species in subtropical regions (de la Fuente et al. 2005a) and also mechanically by certain hematophagous dipterans such as tabanid horse flies (de la Fuente et al. 2005b). Although Dermacentor spp. ticks are the biological vectors of A. marginale in North America, the main tick vector in Europe seems to vary depending on the region (Kocan et al. 2010). The results of a study conducted in 2005 in Sicily showed that among 8 collected tick species, including D. marginatus and I. ricinus, only Rhipicephalus turanicus and Haemaphysalis punctata were found to be infected with A. marginale (de la Fuente et al. 2005a). In Spain, H. marginatum and Rhipicephalus bursa were identified as potential biological vectors for A. marginale (de la Fuente et al. 2004a). However, a study performed in Hungary in 2008 reported the presence of A. marginale in Tabanus bovis and not in D. marginatus, D. reticulatus, I. ricinus, and Haemaphysalis concinna ticks, suggesting that mechanical transmission by tabanids may be more important than the biological vector role of hard ticks in this region (Hornok et al. 2008).
Theileria spp. were not identified in this study, except for 1 D. marginatus female that was found positive for T. annulata with a possible co-infection with B. bovis. This protozoan parasite is implicated in tropical theileriosis and is transmitted by ticks of the genus Hyalomma (Jongejan et al. 1983). Tropical cattle theileriosis is distributed in the Mediterranean and Middle East regions from Morocco to western parts of India and China. This geographical distribution may explain the fact that T. annulata was recovered here in southern France only. To our knowledge, this is the first report of T. annulata in France and suggested that D. marginatus ticks are susceptible to infection with this parasite.
Babesiosis is a worldwide tick-borne hemoprotozoosis affecting many mammalian species (Chauvin et al. 2009). In France, the most prevalent Babesia species corresponds to B. divergens, a bovine parasite that may infect humans and is transmitted by I. ricinus (L'Hostis and Chauvin 1999). In this study, B. divergens was detected in I. ricinus ticks collected from 2 study sites with a prevalence lower than that previously reported in northern France (20.6%; Halos et al. 2005) but similar to that found in western France (9.8%; Cotté et al. 2010). Although D. marginatus is considered a potential vector of B. divergens (Estrada-Peña and Jongejan 1999), this parasite was not recovered from Dermacentor spp. ticks in France. The fact that our study was performed in bovine pastures may explain why Babesia sp. EU1 was not identified in collected ticks. This Babesia species was recovered with high prevalence from ticks collected in French forests, where roe deer and not cattle are suspected as the main reservoir host (Duh et al. 2005, Bonnet et al. 2007, Reis et al. 2011b). B. bovis was identified in a D. marginatus female tick collected in southern France. B. bovis is a tick-borne protozoan parasite transmitted by Rhipicephalus spp. ticks that infects cattle in tropical and subtropical regions (Bock et al. 2004). As previously discussed, B. bovis was identified in the same tick infected with T. annulata, a pathogen also found in more tropical regions (Genis et al. 2008). This result suggested the introduction of cattle persistently infected with these pathogens in the study site where these parasites were recovered for the first time in France. However, as in previous cases with other ticks/pathogens, the finding of a D. marginatus infected with these 2 parasites does not imply that they are transmitted by this tick but maybe simply that the tick acquired infection after immatures feed on an imported and infected animal.
In this study, 7% of collected I. ricinus females were found infected with B. burgdorferi s.l. Borrelia prevalence in this tick species range from 0% to 36% in France (Randolph 2001, Halos et al. 2005, Ferquel et al. 2006, Beytout et al. 2007, Cotté et al. 2010, Reis et al. 2011b), demonstrating a high variation in pathogen prevalence between different regions. Ticks from the I. ricinus complex are considered the main vectors of B. burgdorferi s.l. (Nadelman and Wormser 1998), the causative agent of Lyme borreliosis, which is the most significant human vector-borne disease in Europe (Smith and Takkinen 2006). However, other tick species have been suspected to transmit these bacteria, and among them are Dermacentor spp. such as D. marginatus (Angelov et al. 1996). Furthermore, studies performed in Germany detected viable Borrelia spp. in D. reticulatus questing ticks with a 11.3% prevalence (Kahl et al. 1992). These results agreed with the finding of B. burgdorferi s.l. DNA in 1 D. reticulatus male analyzed in our study. As previously reported in Spain (Toledo et al. 2009a), Borrelia spp. DNA was not found in D. marginatus. Again, the possible role of D. reticulatus and D. marginatus in the transmission of B. burgdorferi s.l. needs to be demonstrated because other Dermacentor spp. ticks such as D. silvarus (Sun and Xu 2003), D. andersoni (Dolan et al. 1997), D. variabilis (Dolan et al. 1997), and D. occidentalis (Lane et al. 1994) are not vectors of Borrelia spp.
The results presented here corresponded to the first systematic study of tick-borne pathogens in Dermacentor spp. ticks in France. These results suggest a role for Dermacentor spp. as vectors of tick-borne pathogens that affect human and animal health. Several pathogens including A. phagocytophilum, A. marginale, B. burgdorferi, Bartonella sp., C. burnetii, B. bovis, T. annulata, and F. philominagia, were detected in D. marginatus and/or D. reticulatus, suggesting a possible role of these tick species in the life cycle and transmission of these pathogens in France. However, without experiments demonstrating the vector competence of these tick species, the epidemiological significance of these findings must be taken with caution, because the presence of a pathogen in ticks does not necessarily mean that they are capable of transmitting it to susceptible hosts. Nevertheless, this information is important for epidemiological studies of tick-borne pathogens in France and to prevent the risks associated with pathogen transmission by Dermacentor spp. ticks to humans and animals. Last, the list of pathogens studied here is not exhaustive, and other microorganisms like Rickettsia spp. or other species of Babesia sp. and Theileria sp. that could be carried and possibly transmitted by Dermacentor spp. should be studied in the future.
Footnotes
Acknowledgments
We thank veterinary practitioners J-M. Nicol, J-R. Clidière, F. Piffoux, A. Chauzy, J-L. Laurent, J. Manière, F. Tonnelle, T. Perrot, and J-F. Rudant for their contribution to the selection of collection sites by G. Joncour, and A. Jolivel, E. Alboussière, A. Senkowski, and M. Joncour-Lostanlen for helping him to collect ticks. We also thank F. Simonnet, K. MacCoy, G. Uilenberg, A. Senkowski, C. Perez-Eid, E. Ferquel, N. Boulanger, D. Raoult, B. Davoust, J-L. Marie, C. Chastel, and the “Tiques et Maladies à Tiques” group (REID- Réseau Ecologie des Interactions Durables) for stimulating discussions and help in the preparation of the project.
Author Disclosure Statement
No competing financial interests exist.