
Abstract
Lobomycosis, a fungal disease of the skin and subcutaneous tissues caused by Lacazia loboi, is sometimes referred to as a zoonotic disease because it affects only specific delphinidae and humans; however, the evidence that it can be transferred directly to humans from dolphins is weak. Dolphins have also been postulated to be responsible for an apparent geographic expansion of the disease in humans. Morphological and molecular differences between the human and dolphin organisms, differences in geographic distribution of the diseases between dolphins and humans, the existence of only a single documented case of presumed zoonotic transmission, and anecdotal evidence of lack of transmission to humans following accidental inoculation of tissue from infected dolphins do not support the hypothesis that dolphins infected with L. loboi represent a zoonotic hazard for humans. In addition, the lack of human cases in communities adjacent to coastal estuaries with a high prevalence of lobomycosis in dolphins, such as the Indian River Lagoon in Florida (IRL), suggests that direct or indirect transmission of L. loboi from dolphins to humans occurs rarely, if at all. Nonetheless, attention to personal hygiene and general principals of infection control are always appropriate when handling tissues from an animal with a presumptive diagnosis of a mycotic or fungal disease.
Introduction

Dolphin with extensive lesions of lobomycosis from the Indian River Lagoon, Florida. Photograph courtesy of E. Howells, Harbor Branch Oceanographic Institution at Florida Atlantic University, 2007. Reprinted with permission.
Thus, the concept that L. loboi might be transmitted to humans from dolphins as a zoonosis seems logical upon initial consideration and indicates that appropriate precautions be used when handling presumptively infected dolphins. Furthermore, recent reports describing lobomycosis or lobomycosis-like disease (LLD) (based on photographic evidence) in dolphins from the Indian Ocean (Kiszla et al. 2009), the Japanese coast (Shirakihara et al. 2008), the western Pacific (Van Bressem et al. 2007, Van Bressem et al. 2009) and the North Carolina coast (Rotstein et al. 2009), as well as in humans from the African continent (Al-Daraji et al. 2008) and the European coast (Papadavid et al. 2012), may indicate that the ecological niche for the disease is expanding. Thus, the risk for transmission to humans, if real, may also increase. Alternatively, increased recognition and reporting may account for the apparent geographical expansion. Several authors have postulated that geographic expansion of lobomycosis in humans may be attributable to direct or indirect contact with infected dolphins that presumably migrated from endemic areas (Paniz-Mondolfi et al. 2012, Papadavid et al. 2012). Direct and indirect human contact with dolphins has expanded due to the increased number of dolphins under managed care in aquaria and commercial exhibitions, swim with dolphin programs for recreational or therapeutic benefit, and occupational exposures of veterinarians, marine biologists, and others who handle dolphins during health assessments, rescue operations, and rehabilitation efforts of stranded or injured animals.
Natural and Experimental Transmission of L. loboi
Support for the zoonotic potential of L. loboi is found in a report describing lobomycosis in a previously healthy 24-year-old aquarium attendant in France who worked in an aquarium pool with an infected bottlenose dolphin (Tursiops truncatus) (Symmers 1983). The dolphin had been caught in the Bay of Biscay off the French–Spanish coast and had multiple nodular lesions of the skin, with histologic evidence of yeast-like fungal cells on lesion biopsy. The patient developed a granuloma on the hand accompanied by supratrochlear lymphadenopathy approximately 3 months after regular exposure to the infected dolphin in the hospital pool in which it was being held. Histologically, the patient's lesions were characteristic of lobomycosis and contained short chains of fungal cells.
With the exception of this single case, further evidence that L. loboi can be transmitted from dolphins to humans under natural conditions is limited. Few cases have been reported in humans from coastal environments (Paniz-Mondolfi et al. 2012). The first was a patient from Surinam (Wiersema et al. 1965), an area where lobomycosis occurs in Guiana dolphins (Sotalia guianensis) that inhabit the Surinam River estuary (DeVries et al. 1973). A second case developed in a young man from South Africa who was an avid swimmer and diver (Al-Daraji et al. 2008). The third occurred in a fisherman from Venezuela who developed lesions on the ear after being pierced with a fishing hook in a coastal area where an affected dolphin was observed (Bermudez et al. 2009). Most recently, lobomycosis was diagnosed in a female farmer with hypoglobulinemia and common variable immunodeficiency and hepatitis who lived on the Greek coast (Papadavid et al. 2012). These observations suggest that the marine environment may occasionally be a source of infection for both species but do not implicate direct transmission from dolphins to humans (Bermudez et al. 2009).
L. loboi can be transmitted directly from human to human under highly unusual circumstances. The first example occurred accidentally following resection of an auricular lesion and placement of a skin autograft obtained from the patient's shoulder where the lesion eventually developed. Transmission was believed to occur through a contaminated needle used to anesthetize the resection site on the ear and then for the skin graft site (Azulay et al. 1970). A second accidental transmission of lobomycosis occurred in a laboratory worker who collected skin biopsies from infected patients in Brazil, purified fungal cells from the biopsy material, and inoculated L. loboi into mice to propagate the organism in the laboratory (Rosa et al. 2009). Similarly, experimental inoculation of a laboratory scientist with yeast-like cells from a human patient shows that under unusual circumstances, Lacazia of human origin can be transmitted to other humans (Borelli 1961). L. loboi can also be transmitted by direct inoculation into mice, hamsters, tortoises, and armadillos (Paniz-Mondolfi et al. 2012). However, a report by a dermatologist who suffered an accidental laceration during a biopsy procedure of a dolphin with lobomycosis (Norton 2006) and anecdotal reports by a veterinary pathologist who was exposed to tissues from infected dolphins through wounds incurred during autopsy on several occasions (G.D. Bossart, personal communication, 2010) without subsequent development of lobomycosis suggest that transmission from dolphins to humans, even through direct inoculation, is unlikely in immunocompetent individuals.
Morphologic and Molecular Studies of L. loboi in Humans and Dolphins
Several lines of evidence support the contention that dolphin to human transmission of L. loboi is extremely rare. First, the organisms demonstrated in tissues from infected dolphins and humans differ morphologically (Fig. 2). The Lacazia cells seen in dolphin tissues are smaller than those in lesions from humans under light microscopy (Haubold et al. 2000). Morphometric analysis of tissue showed that human Lacazia have approximately twice the area and are 30% longer in their major axis than the organisms in dolphin tissues (Haubold et al. 2000). Furthermore, the ultrastructural characteristics of cell wall destruction differ between dolphins and humans. The authors postulated that the morphologic differences may result from environmental differences in salinity, temperature, and an aqueous environment, or, alternatively that the differences may represent the organism's cellular response to physiologic conditions within the host. A third possibility is that the organisms from humans and dolphins represent true phylogenetic divergence (Haubold et al. 2000).
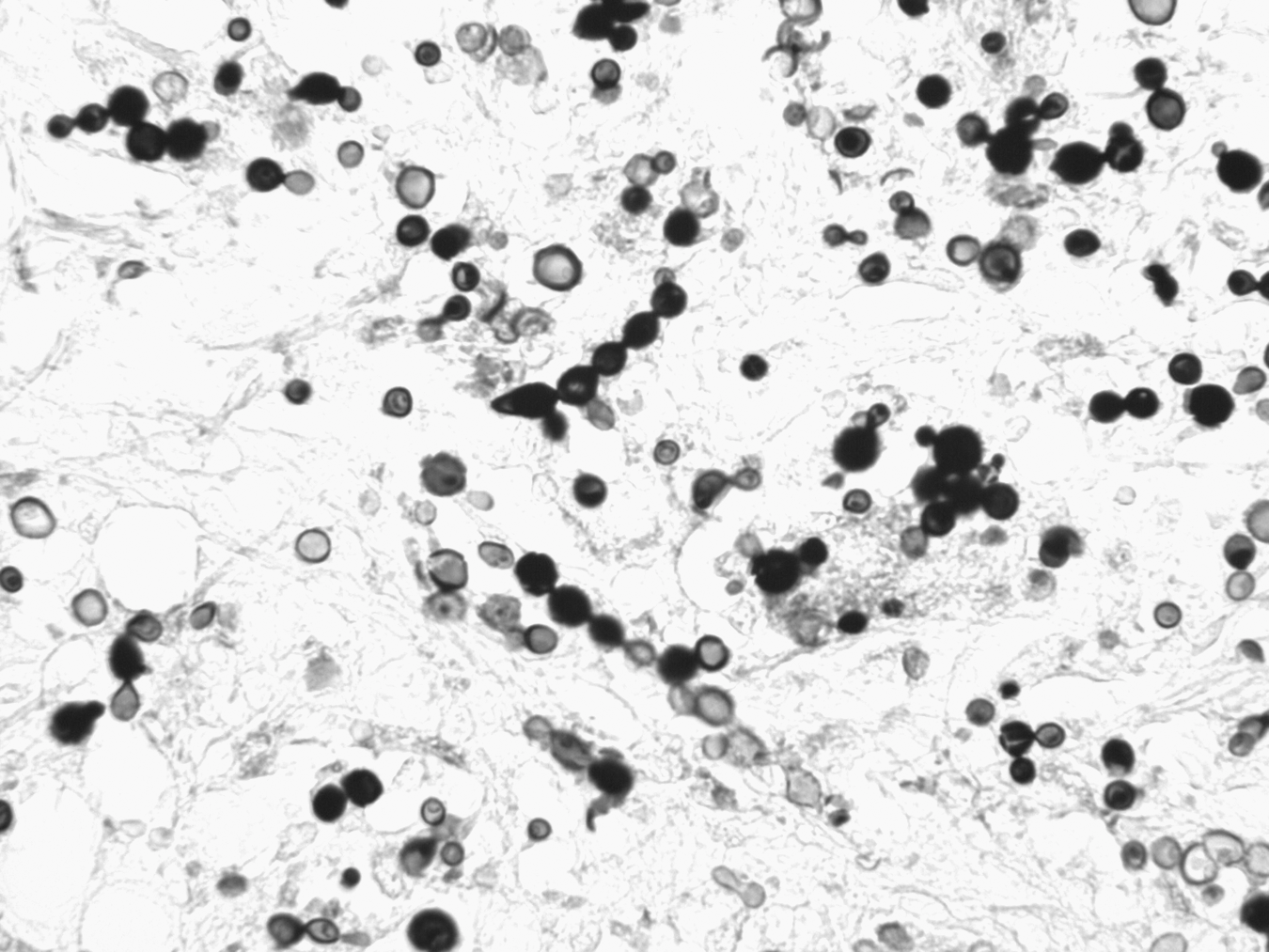
Photomicrograph of dermal tissue from a dolphin with lobomycosis. The yeast-like cells are seen singly and in chains connected by tube-like bridges, characteristic of Lacazia loboi. Gomori methenamine silver stain. Magnification, 400×.
In support of the latter hypothesis, molecular sequencing of ribosomal DNA from an infected dolphin showed a novel sequence related more closely to Paracoccidiodes brasiliensis than to L. loboi of human origin (Esperón et al. 2012). This recent evidence is supported by an earlier study that showed that ribosomal RNA gene sequences from an infected dolphin were 97% homologous with P. brasiliensis (Rotstein et al. 2009). However, sera from infected dolphins recognized the immuno-dominant ≈193-kDa antigen from an extract of human L. loboi more strongly than the gp43 antigen of P. brasiliensis in western blotting analyses (Mendoza et al. 2008). Serological cross-reactivity between serum from human patients with lobomycosis and P. brasiliensis has also been demonstrated (Camargo et al. 1998). However, direct molecular and phylogenetic comparison of Lacazia of dolphin and human origin has not been reported.
Epidemiologic Characteristics of Human and Dolphin Lobomycosis
The endemicity and natural history of lobomycosis in dolphins and humans are strongly contrasted. Lobomycosis was first reported in a Florida bottlenose dolphin (T. truncatus) in the Gulf of Mexico in 1971 (Migaki et al. 1971) and is endemic in south Florida's coastal estuaries. Early reports describe sightings of apparently infected dolphins in the Florida Keys, and the Indian River Lagoon (IRL) dating to the 1950s (Caldwell et al. 1975). The IRL, an estuary that extends 156 miles along approximately 40% of Florida's eastern coastline, is a hyperendemic locale for the disease in dolphins. Capture–release health assessment studies showed that the prevalence in the IRL is approximately 10% and that the disease aggregates in the southern reaches of the estuary (Reif et al. 2006). In this zone, an area with slightly higher water temperatures and lower salinity than the northern IRL, the prevalence is higher, approaching 17% (Murdoch et al. 2008). Geographic differences in the prevalence of lobomycosis and LLD (Burdette Hart et al. 2011) suggest that specific features of the marine environment create a suitable niche for Lacazia and potential transmission to dolphins or humans. However, human lobomycosis has not been reported among approximately 1.7 million persons who inhabit the coastal counties bordering the IRL. Extensive human exposure to the IRL through recreational activities suggests that the organism could be introduced into the dermis through cuts, punctures, and abrasions, if, in fact, the Lacazia of dolphins was a human pathogen under natural circumstances. Lobomycosis is extremely rare in industrialized countries, diagnosed almost exclusively among persons with a history of exposure to tropical environments (Elsayed et al. 2004). For example, the first case reported in the United States occurred in 2000 in a traveler who had visited Venezuela frequently and was likely exposed through contact with a waterfall (Burns et al. 2000).
Conversely, lobomycosis in humans is endemic in tropical regions of Central and South America and is most frequently associated with exposure to fresh water habitats, vegetation, and soil (Rodriguez-Toro 1993). Lobomycosis associated with occupational exposures to farmers, hunters, mineral explorers (gold panning), rubber workers, fishermen, and others has been reported widely in endemic areas (Fuchs et al. 1990, Rodriguez-Toro 1993). Transmission in occupational and residential settings appears to occur by cutaneous introduction of the organism through trauma, including cuts, snakebites, ray stings, insect bites, and abrasions on exposed surfaces of the body. Several high-risk occupational groups have regular exposure to fresh water aqueous environments. However, lobomycosis has not been reported in either of the two freshwater species of dolphins that inhabit the Amazon and Orinoco river basins of Brazil and Venezuela where the human disease is endemic, botos (Inia geoffrensis) or tucuxis (Sotalia fluviatilis) (Paniz-Mondolfi et al. 2009).
The Immune System in Dolphin and Human Lobomycosis
Defects in the immune system may constitute a common pathway for infection of humans and dolphins with Lacazia. Lobomycosis has been described in a dolphin with hypogammaglobulinemia and chronic-active hepatitis (Bossart 1984), and similarly in a woman with hypoglobulinemia and cholestatic hepatitis (Papadavid et al. 2012), suggesting that the disease may occur in immunocompromised dolphins and humans. However, only a single case of lobomycosis has been described in a patient infected with human immunodeficiency virus (HIV) (Xavier et al. 2006).
In human lobomycosis, impairment of cellular immunity is characterized by failure to become sensitized to dinitrochlorobenzene, negative hypersensitivity skin tests to common bacterial and fungal pathogens, and a delayed skin allograft rejection (Pecher and Fuchs 1988). Depressed cellular immunity may be responsible for the chronicity of the lesions and failure to resolve spontaneously (Rodriguez-Toro 1993). Mononuclear cells obtained from patients with lobomycosis exhibit cytokine profiles predominantly of the Th2 profile (Vilani-Moreno et al. 2004a). A higher frequency of immunosuppressive cytokines transforming growth factor-β (TGF-β1) and interleukin-10 (IL-10) found in the skin lesions of patients suggests that immunoregulatory disturbances may be responsible for the large number of fungal cells present in the granulomas and lack of pathogen containment (Vilani-Moreno et al. 2005, Vilani-Moreno et al. 2011).
Similarly, dolphins with lobomycosis have multiple defects in adaptive immunity (Reif et al. 2009). Compared to healthy dolphins, affected dolphins had reduced numbers of circulating helper T cells and mature and immature B cells as well as a reduction in the number of cells expressing major histocompatibility complex (MHC) class II molecules. Mitogen-induced lymphocyte proliferation of T and B cells was reduced substantially. Changes in lymphocyte surface markers and proliferation were accompanied by reduced antibody titers to common marine microorganisms. No evidence for impaired phagocytosis in monocytes and granulocytes obtained from dolphins with lobomycosis was found, compatible with a report that phagocytosis of L. loboi by monocytes from human patients did not differ from controls (Vilani-Moreno et al. 2004b). Despite evidence that humans and dolphins have substantial defects in cell-mediated immunity, it remains unclear whether infection with Lacazia represents an opportunistic infection or whether infection leads to secondary changes in immune function (Reif et al. 2009, Honda and Raugi 2013).
Conclusions
In summary, morphological and molecular differences between the human and dolphin organisms, differences in geographic distribution, the lack of human cases in an endemic, high-prevalence area of dolphin lobomycosis, the existence of only a single documented case of presumed zoonotic transmission, and anecdotal evidence suggest that direct transmission of L. loboi from dolphins to humans is highly unusual. The risk of acquiring a Lacazia infection from a dolphin with clinical evidence of lobomycosis appears to be very low. Nonetheless, appropriate hygiene and care are always appropriate when handling tissues from an animal with a presumptive diagnosis of an infectious disease.
Footnotes
Acknowledgment
The work was supported in part by Florida's Protect Wild Dolphins specialty license plate fund, through Harbor Branch Oceanographic Institution at Florida Atlantic University.
Author Disclosure Statement
No competing financial interests exist.