
Abstract
Arboviral activity tracks vector availability, which in temperate regions means that transmission ceases during the winter and must be restarted each spring. In the northeastern United States, Culex restuans Theobald resumes its activity earlier than Culex pipiens L. and is thought to be important in restarting West Nile virus (WNV) transmission. Its role in WNV amplification, however, is unclear, because viral levels commonly remain low until the rise of Cx. pipiens later in the season. Because a vector's feeding habits can reveal key information about disease transmission, we identified early-season (April–June) blood meals from Cx. restuans collected throughout New Jersey, and compared them to published datasets from later in the season and also from other parts of the country. We found significantly higher avian diversity, including poor WNV hosts, and fewer blood meals derived from American Robins (17% versus over 40% found in later season). Critically, we identified blood meals from significantly more female than male birds in species where females are the incubating sex, suggesting that Cx. restuans is able to feed on such a wide variety of hosts in early spring because incubating birds are easy targets. Because WNV amplification depends on virus consistently reaching competent hosts, our results indicate that Cx. restuans is unlikely to be an amplifying vector of WNV in the early season. As the season progresses, however, changes in the availability of nesting birds may make it just as capable as Cx. pipiens, although at somewhat lower abundance as the summer progresses.
Introduction
I
In climates with cold winters that suppress vector activity, the cycle of transmission of arboviruses like WNV and SLE needs to be reinitiated each year. Because Cx. restuans are among the first bird biters to emerge from diapause in the northeastern United States, they are considered critical for initiating WNV transmission among birds in early spring, whereas Cx. pipiens amplifies viral levels later in the season (Andreadis et al. 2001). The same scenario has been proposed by Reiter (1988) for the relative roles of Cx. restuans and Cx. pipiens in the transmission of SLE. Although extrinsic factors such as temperature and rainfall affect the intensity of WNV transmission (Kramer et al. 2008), it is unclear why WNV levels remain low until summer, even after temperatures permissible for viral replication have been reached (Dohm et al. 2002, Kilpatrick et al. 2008). One way to investigate the early-season amplification of WNV is to examine the composition and diversity of Cx. restuans blood meal hosts during this time. Most passerine species are competent WNV hosts, whereas many other groups of birds, such as doves and shorebirds, are not (Komar et al. 2003, Blitvich 2008), and higher bird species diversity has been linked to lower WNV transmission (Ezenwa et al. 2006, Allan et al. 2009), a phenomenon known as the dilution effect (Ostfeld and Keesing 2000).
In this study, we examined the blood-feeding paterns of Cx. restuans collected in the early season in locations throughout New Jersey over a 10-year period. We analyzed the species diversity and sex ratio of blood meal hosts and characteristics of individual bird species, such as WNV competence, presence of migratory behavior, and egg incubation. We also compared our results to blood-feeding studies of Cx. restuans collected later in the season from the same region, as well as in Illinois and Tennessee. The latter locations were an effort to account for differences in bird assemblages and identify temporal patterns.
Methods
Collection of mosquitoes
Due to the difficulty of obtaining blooded mosquitoes in the early season, the specimens used in this analysis were collected by collaborators from a variety of locations throughout 12 New Jersey counties over 10 years, 2001–2011. Specimens were mostly collected as part of local surveillance efforts by mosquito control agencies using light traps, gravid traps, resting boxes, BioGents Sentinel traps (Biogents, AG, Regensburg, Germany), and backpack aspiration. All blooded specimens identified as Cx. restuans on the basis of morphology were immediately placed in a −80°C freezer and held for extraction and molecular identification. A subset collected between April 15 and June 15 across all years was analyzed in this study (n=113). We chose to not identify the blood meals of late-season Cx. restuans because specimens collected in the same manner and from many of the same sites as ours have already been analyzed and published (Apperson et al. 2004) and are included in the late season “Northeast” dataset (see below).
Identification of blood meals
Blooded abdomens were removed over ice, and DNA was extracted using a Qiagen DNeasy Blood and Tissue Kit (Qiagen, Valencia, CA). Only specimens confirmed to be Cx. restuans (105/113 specimens) using a molecular assay (Crabtree et al. 1995) were included in the final analysis. Specimens were first screened with BM primers (Kocher et al. 1989), and those that did not amplify were subsequently tested with Avian A (Cicero and Johnson 2001), then Rep/Amph (Cupp et al. 2004), and finally Mammal A primers (Ngo and Kramer 2003) following established protocols. Amplification products were sequenced with the forward primer of each pair. We identified the putative source of the blood using a GenBank BLAST search with a match ≥98% (Kent 2009). Mixed sequences were cloned and sent for sequencing. A total of 83 specimens (79.0%) were successfully identified using these methods.
To identify additional samples, a smaller 130-bp fragment was amplified using the primers CytbVertR1 (Egizi et al. 2013) and BMF1 (originally called H15149; Kocher et al. 1989). One mixed sequence amplified with these primers was cloned. This approach allowed the additional identification of 11 blood meals for a total of 94 (89.5%) identified blood meals in Cx. restuans.
Comparison with previous studies
Existing data on Cx. restuans avian blood-feeding habits was obtained from published studies and pooled by geographic location into three datasets. The Northeast data set encompassed four studies conducted in New Jersey, New York, and Connecticut (Apperson et al. 2002, Apperson et al. 2004, Molaei et al. 2006, Diuk-Wasser et al. 2010); the IL dataset was from two studies in southwest Chicago, Illinois (Hamer et al. 2008, Hamer et al. 2009); and the TN dataset was from Shelby County, Tennessee (Savage et al. 2007). Only studies that used molecular techniques to distinguish Cx. restuans from Cx. pipiens and that published a breakdown of all avian blood meals by species were included. These datasets were compared with the early-season New Jersey data from our study (called NJearly).
We analyzed a table of counts of each bird species in each dataset using a correspondence analysis with the ca package (Nenadic and Greenacre 2007) in the statistical software program R (R Core Team 2013). Proportion of American Robin blood meals was compared across datasets, as robins have been implicated as an important amplification host of WNV (Kilpatrick et al. 2006a). Because the presence of migrant birds may also affect WNV transmission, we calculated the proportion of migratory bird blood meals. A species was considered migratory for a particular dataset if it is not considered a year-round resident in the area where that dataset was collected (Poole 2005). Finally, because of their ability to dampen WNV transmission, we also calculated the proportion of nonpasserine blood meals in each dataset. Each proportion (robin, migratory, and nonpasserine) in the published datasets was compared to the early-season New Jersey data in a chi-squared test.
We calculated Rényi profiles, a graphical means of comparing species diversity between samples, in R (R Core Team 2013) using the packages BiodiversityR (Kindt and Coe 2005) and vegan (Oksanen et al. 2013). Rényi profiles plot Rényi entropy (Hα) over a range of alphas (α) using the formula:
where pi is the partial abundance of species i and S is the total number of species in the dataset (Rényi 1961, Tóthmérész 1995). For different values of α, this measure is related to several well-known measures of diversity including: α=0 for simple species richness, α=1 for Shannon diversity, α=2 for the Simpson index, and α=∞ for Berger–Parker diversity (Tóthmérész 1995). Therefore, Rényi profiles present a more comprehensive picture of diversity than looking at any measure individually and allow diversity to be ranked among sites (Tóthmérész 1995, Kindt and Coe 2005). Other diversity measures calculated were Shannon diversity (Shannon and Weaver 1949), inverse Simpson diversity (Simpson 1949), and the Pielou measure of evenness (Pielou 1966), using BiodiversityR and vegan (Kindt and Coe 2005, Oksanen et al. 2013).
Blood meal sexing
We used the assay described by Griffiths et al. (1998), which relies on a size discrepancy between the CHD-Z and CHD-W alleles (male birds are ZZ and females are ZW) (Griffiths et al. 1998) and works on a large variety of bird species. However, because in Cooper's Hawks the target fragments differ by only 2 base pairs (GenBank accession nos. JX460775 and JX460774) after amplification, we digested those PCR products with DraI (New England Biolabs, Beverly, MA). DraI cuts only the CHD-W allele, producing three bands in females and one in males. All PCR products and digestion products were run on a 3% agarose gel for 90 min to ensure separation of DNA bands.
Male and female blood meals were pooled across all avian species and tested in a chi-squared test against a null hypothesis of 1:1 male:female. A second chi-squared test omitted all species with male incubation, identified using the Birds of North America Online database (Poole 2005), as well as the Brown-headed Cowbird, a nest parasite that does not incubate its own eggs.
Results
We identified blood meals from 94 Cx. restuans mosquitoes to species. Two specimens (∼2%) contained mixed blood meals—one a two-way mix between human and House Finch and the other a three-way mix between American Robin, Tufted Titmouse, and Cooper's Hawk. All other mixes were cases where the primers amplified mosquito DNA instead of that of the vertebrate host, a phenomenon we have observed when the vertebrate DNA is highly degraded (A.E. Egizi, personal observation). The human blood identified in the mix with House Finch was the only mammalian meal detected in our studies from Cx. restuans. The remaining blood meals contained 96 bird sequences from 23 avian species (Table 1).
“Code” is the four-letter code used for that species in Fig. 1.
Species found in three-way mixed meal.
Species found in a mixed meal with human.
The two datasets in the northeast United States (NJearly and Northeast) had similarly high species richness compared to IL and TN, although sample sizes were significantly smaller (Table 2). Importantly, however, the NJearly dataset had a more even distribution of species compared to the other three resulting in the highest Shannon and Simpson diversity indices (Table 2). These patterns are reflected in the Rényi diversity profiles in Figure 1, where the NJearly profile has a flatter slope, indicating greater evenness than the other communities. Because the TN dataset is consistently lower across all values of α, it has the lowest overall diversity, followed by IL; however, the top two profiles cannot be definitively ordered using this method because they intersect (Fig. 1).
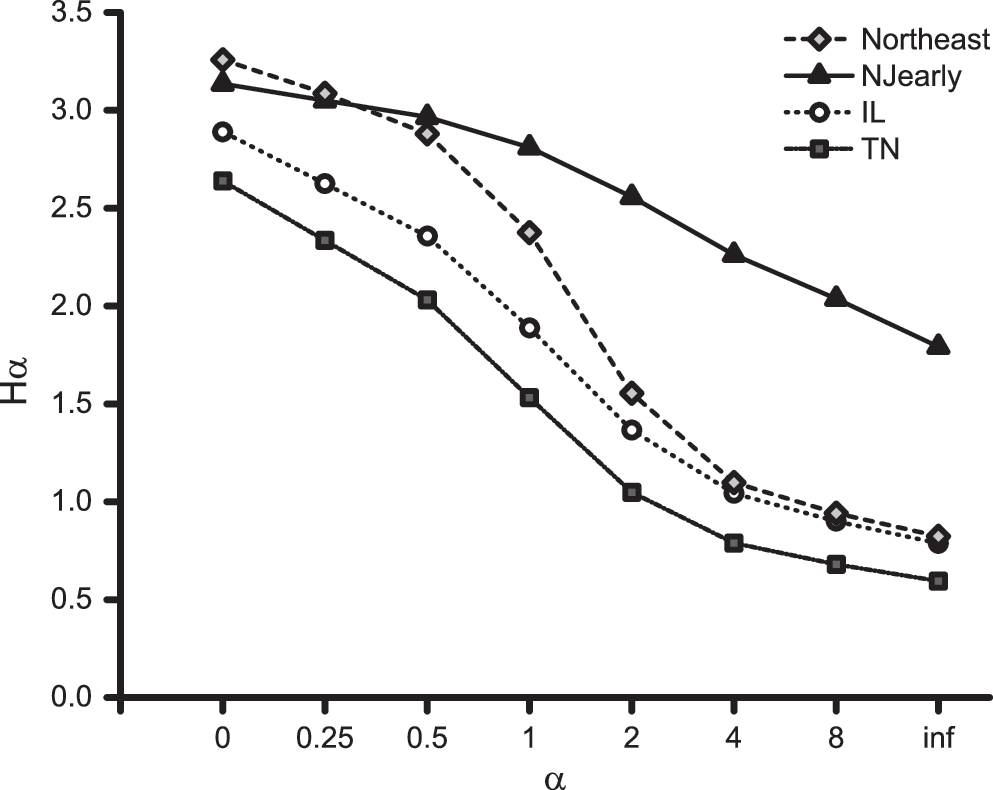
Rényi diversity profile values (Hα) plotted over a range of α for each blood meal dataset. Smaller values of α tend to weigh species richness more heavily, whereas larger values weigh evenness more heavily. Shannon and Simpson diversity, which include both factors in their calculation of diversity, fall into the middle (α=1 and α=2, respectively). Northeast=the dataset comprising five studies conducted in New York, New Jersey, and Connecticut; NJearly=the dataset from this paper; IL=dataset comprising two studies in and around Chicago, IL; TN=dataset from Shelby County, TN.
N, number of blood meals; S, avian species richness; H′, Shannon information index; 1/D, inverse Simpson diversity index; J′Pielou's measure of evenness, calculated as H′/ln(S);
The NJearly dataset has the highest evenness because of a significantly lower proportion of American Robins (Tables 2 and 3). Both the NJearly and northeast datasets had significantly higher proportions of migratory birds and nonpasserine birds, compared to the TN or IL data (Tables 2 and 3). In a correspondence analysis, NJearly is separated from the other three datasets along Axis 1, which makes up 50% of the variation (Fig. 2). Note that American Robin (AMRO) falls near the other datasets along Axis 1 (Fig. 2).
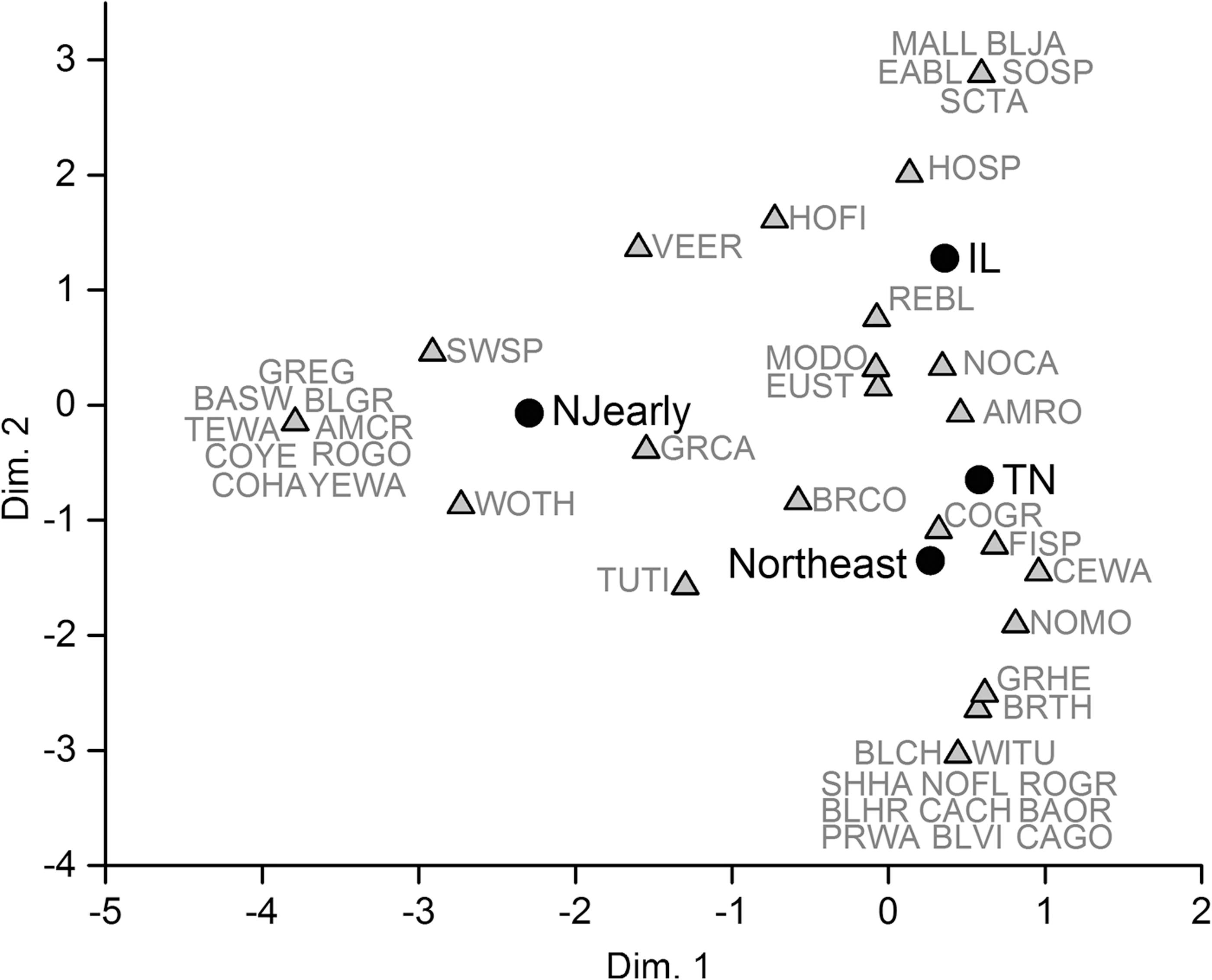
Correspondence analysis biplot of host species (grey triangles) and location dataset (black circles) scores in dimensions 1 and 2 explaining 49.69% and 26.74% of the variance, respectively. Species codes follow Table 1, with additional acronyms for those not in this dataset but in others as follows: BAOR, Baltimore Oriole; BLCH, Black-capped Chickadee; BLHR, Black-crowned Night Heron; BLJA, Blue Jay; BLVI, Blue-headed Vireo; BRTH, Brown Thrasher; CACH, Carolina Chickadee; CAGO, Canada Goose; CEWA, Cedar Waxwing; EABL, Eastern Bluebird; FISP, Field Sparrow; GRHE, Green Heron; MALL, Mallard; NOFL, Northern Flicker; NOMO, Northern Mockingbird; PRWA, Prairie Warbler; ROGR, Rose-breasted Grosbeak; SCTA, Scarlet Tanager; SHHA, Sharp-shinned Hawk; SOSP, Song Sparrow; WITU, Wild Turkey. Species that are strongly associated with a particular dataset are clustered near it on the plot.
p<0.001.
The sex of the blood meal host was determined in 80/94 samples (Table 4). The overall sex ratio was not significantly different from 1:1 in a chi-squared test (χ2=1.25, degrees of freedom [df ]=1, 0.50<p<0.25). However, the sex ratio was significantly female-biased in species with female-only incubation (χ2=6.12, df=1, p<0.05).
Discussion
Our study shows that early-season Cx. restuans collected in New Jersey feed almost exclusively on birds, although we did find a single human meal present in a mix. Previous studies of Cx. restuans in New York (Patrican et al. 2007) and Connecticut (Molaei et al. 2006, Diuk-Wasser et al. 2010) found exclusively avian meals, whereas other studies found as much as 13–32% mammalian (Apperson et al. 2002, Apperson et al. 2004, Savage et al. 2007, Hamer et al. 2008, Hamer et al. 2009). The frequency of mixed meals we observed (2%) is within the range found in previous studies (1–4%) (Apperson et al. 2002, Savage et al. 2007, Hamer et al. 2009). The three-way mixed meal we identified is a unique find and may have been possible because our primers targeting a smaller DNA fragment (130 bp) facilitated detection of degraded DNA. Our dataset and the Northeast dataset had a significantly greater proportion of migratory birds (19% and 17%, respectively) than either the IL or TN data (4% for both). This is probably due to geography, because the mid Atlantic coast (particularly the Cape May and Delmarva peninsulas) is considered a major stopover for neotropical migrants (Mabey et al. 1993). In New Jersey, only 22 out of 350 avian species detected annually in the state are permanent residents (Dunne et al. 1989), so the majority of the 210 birds observed breeding in the state each year (Boyle 2011) are migrants.
Numerous studies from multiple parts of the country have found that Culex mosquitoes derive a large portion of their meals from American Robins (Apperson et al. 2002, Apperson et al. 2004, Kilpatrick et al. 2006 , Molaei et al. 2006, Savage et al. 2007, Hamer et al. 2008, Hamer et al. 2009, Kent et al. 2009, Diuk-Wasser et al. 2010, Montgomery et al. 2011). They also seem to prefer robins in laboratory choice tests (Simpson et al. 2009) and feed in greater frequency than predicted by its abundance in the field, leading researchers to postulate that robins are major amplification hosts of WNV (Kilpatrick et al. 2006a). In our study of early-season Cx. restuans, although robins are still the most abundant species at 17%, this number is significantly lower than in the other three datasets (Tables 2 and 3).
It is possible this difference is a consequence of our broad sampling approach, collecting mosquitoes from 12 different counties and in many different types of environments. Most studies focus their efforts in a smaller area, often in urban and suburban neighborhoods where WNV has been detected. In New Jersey, residential areas contain significantly more robins and House Sparrows than adjacent wetland areas (Johnson et al. 2012), so sampling in residential areas might yield more robin blood meals. However, the Northeast dataset comprised four different studies in three different states, with a range of environments (of note, several of the specimens were obtained from the same sources as ours), and in fact had similar species richness and sample size to our study, whereas TN and IL had lower species richness despite larger sample sizes. This indicates that at least for the comparison between our data and the Northeast data, collection bias does not explain the difference in diversity patterns.
More likely, then, these differences result from divergent host behavior in the early season. Changes in the behavior of robins at the conclusion of the breeding season have been invoked to explain why Cx. pipiens shift their feeding away from robins in late summer (Kilpatrick et al. 2006b, Diuk-Wasser et al. 2010). Conversely, it is possible that the early-season behavior of robins makes them less available as hosts, or more importantly, that early-season behavior of other bird species makes them more available. Behaviors likely to influence avian host availability in the early season relate to reproduction, such as egg incubation and brooding of young. In Alabama, mosquito blood meals identified from herons peaked during the height of heron nesting season (Burkett-Cadena et al. 2011), whereas in Tennessee, the number of blood meals from robins oscillated up and down over the season, possibly in response to this species' multiple broods per year (Savage et al. 2007). Nestlings have been considered especially susceptible to mosquitoes due to their decreased defensive behaviors, feathers, and motility; however, comparisons of landing and feeding rates on nestlings versus adults did not find evidence for this, possibly because adults shield them from bites during brooding (Griffing et al. 2007, Burkett-Cadena et al. 2010).
Instead, adult birds sitting on the nest for long periods of time may themselves become easy targets for mosquitoes. For example, Cooper's Hawks have an incubation time of over a month during which the female is on the nest day and night, leaving only when fed periodically by the male (Baicich and Harrison 1997, Poole 2005). In New York, this typically occurs from late April to early May (Meng 1951). Our samples collected during this time period showed a large number of Cooper's Hawk blood meals compared to other studies, the majority of which (8/10 sexed meals) are female. In addition, we found a significant bias toward female blood meals across all species in which the female alone performs incubation duties, suggesting that Cx. restuans targets incubating birds for blood feeding. This could explain why we observed a greater diversity of hosts in the early season, as the assortment of nest-bound birds in early spring could act like a “buffet for mosquitoes,” whereas hosts are more difficult to locate and target once nestlings fledge and nests are abandoned.
The dilution effect hypothesis predicts that greater host diversity should result in lesser disease incidence (Ostfeld and Keesing 2000). This has been shown for WNV and nonpasserine diversity (Ezenwa et al. 2006, Allan et al. 2009) because nonpasserines tend to be poor WNV hosts (Komar et al. 2003, Blitvich 2008). Our dataset contains a large number of meals from Cooper's Hawks and Mourning Doves, which as Accipitriformes and Columbiformes, respectively, are poor hosts of WNV (Komar et al. 2003). We also detected blood meals from wetland birds (e.g., Great Egret, Ross's Goose), and areas with large colonies of wetland birds have been associated with lower mosquito infection rates (Reisen et al. 2009).
Conclusions
Our study reveals that specific bird behaviors associated with the time of year, such as egg incubation in the spring, may be critical drivers of WNV amplification dynamics. This is in contrast to current hypotheses that WNV amplification hinges on the occurrence of specific vector species, such as Cx. pipiens. In fact, Cx. restuans is similar to Cx. pipiens in its late season host usage and therefore their different roles in WNV transmission may result from differences in seasonal timing rather than innate species characteristics.
Footnotes
Acknowledgments
We thank Robert Fleischer (Smithsonian Institution) for information about sexing birds. We thank the following for sample collection and identification: Linda McCuiston, Rutgers University; Tony Acquaviva, Monmouth County Mosquito Extermination Commission; Peter Bosak and Diane McNelly, Cape May Mosquito Control; Christine Musa and Sara May, Warren County Mosquito Extermination Commission; Bill Reinert and Dana Chort, Atlantic County Mosquito Control; Rich Candeletti and Mike Romanowski, Ocean County Mosquito Extermination Commission; Sean Devaney, Union County Mosquito Control; and the Salem County Mosquito Extermination Commission. This work was funded by USDA Hatch Grant no. NJ08194, NE-1043 Multistate funds, and start-up funds to D.M. Fonseca.
Author Disclosure Statement
No competing financial interests exist.