
Abstract
In late 2013, chikungunya virus (CHIKV) was introduced to the New World and large outbreaks occurred in the Caribbean islands causing over a million suspected and over 20,000 laboratory-confirmed cases. Serological analysis is an essential component for the diagnosis of CHIKV infection together with virus isolation and detection of viral nucleic acid. Demonstrating virus neutralizing by serum antibodies in a plaque reduction neutralization test (PRNT) is the gold standard of all serological diagnostic assays. Prior to the testing, heat inactivation of serum at 56°C for 30 min is required for the inactivation of complement activity and adventitious viruses. The presence of adventitious contaminating viruses may interfere with the results by leading to a higher number of plaques on the monolayers and subsequent false-negative results. This procedure is widely accepted for the inactivation of flaviviruses and alphaviruses. In this study, the thermostability of CHIKV was evaluated. Heat inactivation at 56°C for 30 min was demonstrated to be insufficient for the complete removal of infectious CHIKV virions present in the samples. This thermotolerance of CHIKV could compromise the accuracy of serum tests, and therefore longer treatment for greater than 120 min is recommended.
Introduction
C
To inactivate serum complement activity and to remove the adventitious viruses prior to performing PRNT, heat inactivation at 56°C for 30 min is recommended by the World Health Organization (WHO) for the diagnosis of DENV (Roehrig et al. 2008). Although the duration of heat inactivation varies slightly between 30 and 40 min among different laboratories, this procedure has been proved to be sufficient for the inactivation of several arboviruses in the Flaviviridae family (Russell et al. 1967, Song et al. 2010). However, inactivation at 56°C for 30 min has been found to be incomplete for Western equine encephalitis virus (WEEV), a New World alphavirus (Fang et al. 2009). In the same study, the minimum time required for heat inactivation of WEEV at 56°C was demonstrated to be at least 90 min. Similarly, a significant fraction of infectious virus was also recovered when Sindbis virus was heated at 51°C for less than 45 min (Meyer et al. 1992). These results challenge the broad application of routine heat inactivation procedures that are widely used for serological diagnosis of arboviruses and other viral pathogens. Because CHIKV has rapidly spread into various locations worldwide, characterizing the thermostability of CHIKV is critical to ensure the accuracy of the results of the serological assays and the safety of laboratory personnel that perform serological diagnosis.
In the study reported here, the assessment of heat inactivation was performed at 56°C with various input quantities of CHIKV and incubation periods. Our results suggest that the currently accepted method of heat inactivation at 56°C for 30–40 min is insufficient for the complete removal of infectious virions of CHIKV.
Materials and Methods
Cell lines and virus
A cDNA infectious clone-derived CHIKV 2006OPY1 strain was used in this study. The virus stock was generated by electroporation of in vitro–transcribed viral RNA in BHK-21 cells and an additional passage in C6/36 cells, as previously described (Tsetsarkin et al. 2006). The titration of samples was performed with a 50% tissue culture infectious dose (TCID50) method on Vero cells using previously published procedures (Higgs et al. 2006).
Heat inactivation
The heat inactivation of CHIKV in this study was performed by placing 3 mL of serially diluted CHIKV in 15-mL conical tubes in a prewarmed water bath to ensure homogeneous heating. All serial dilutions were performed with L-15 medium containing 10% fetal bovine serum (FBS) and tryptose phosphate broth. Three independent sets of samples were generated to ensure the reproducibility of results. The initial aliquot was collected for the quantification of input titers. To monitor the thermostability of CHIKV at 56°C, virus inocula ranging from 7.95 logTCID50/mL to 5.35 logTCID50/mL were heated for 1 h at 56°C and sampled every 20 min. To investigate the minimum time required for complete heat inactivation of CHIKV relative to the recommended heat inactivation procedure by the WHO, virus stocks at 7.66 logTCID50/mL and 4.52 logTCID50/mL were heated for 40, 80, 120, and 160 min at 56°C. All of the samples were immediately aliquoted and frozen at −80°C prior to titration.
Statistical analysis
Linear mixed-effects models that can incorporate the association resulting from repeated measurements in the same tube were used to evaluate the effects of the initial viral titers and the incubation time for heat inactivation on the residual viral titers.
Results
The test for the thermostability of CHIKV at 56°C was performed with four different titers of viral stocks at 7.95, 7.13, 6.19, and 5.35 logTCID50/mL, respectively. The results demonstrated the tolerance of CHIKV for heat inactivation, as summarized in Figure 1. With the average initial titer at 7.95 logTCID50/mL, the heat inactivation at 56°C for 20, 40, and 60 min led to the average residual viral titers at 5.33 logTCID50/mL, 5.14 logTCID50/mL, and 3.62 logTCID50/mL, respectively. The average residual viral titers in the group receiving inoculum at 7.33 logTCID50/mL after the heat inactivation for 20, 40, and 60 min were 4.28 logTCID50/mL, 3.62 logTCID50/mL, and 3.62 logTCID50/mL, respectively. In the group with average initial viral titer at 6.19 logTCID50/mL, the average viral titers declined to 3.00 logTCID50/mL, 2.19 logTCID50/mL, and 2.58 logTCID50/mL after 20, 40, and 60 min of heat inactivation, respectively. The heat inactivation of the inocula at 5.35 logTCID50/mL resulted in the decline of average titers to 2.81 logTCID50/mL, 1.16 logTCID50/mL, and 0.86 logTCID50/mL at 20, 40, and 60 min from the start of heat inactivation respectively. Overall, the residual titers after 60 min of heat inactivation showed significantly negative association with incubation time (p < 0.05).
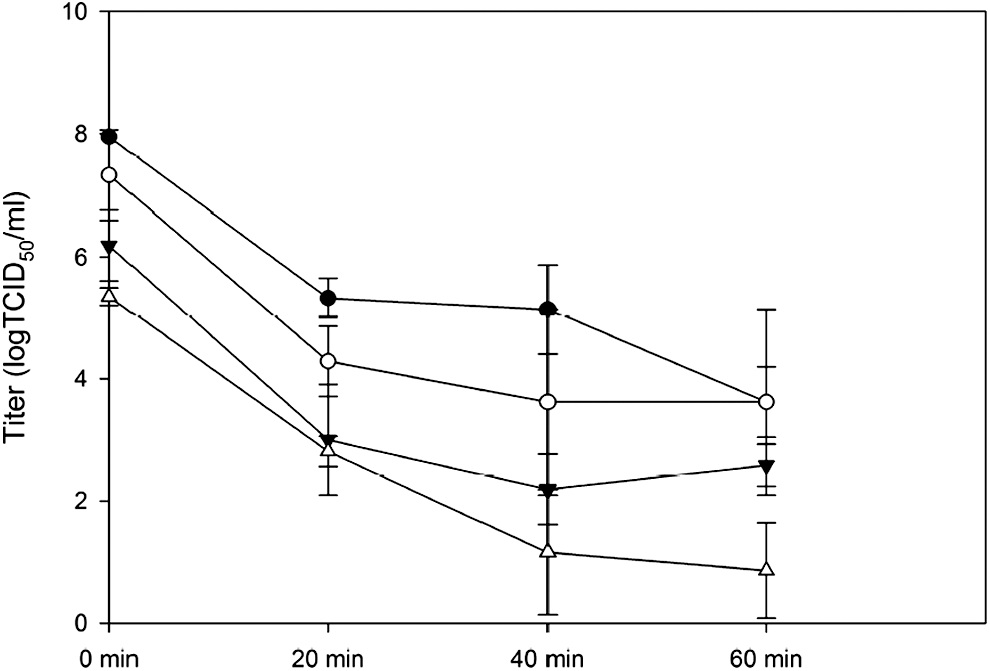
Thermostability of chikungunya virus (CHIKV) at 56°C. Four aliquots of CHIKV with different titers were heat inactivated at 56° for 60 min and sampled every 20 min. Average input titers of each aliquot were 7.95 log 50% tissue culture infectious dose (TCID50)/mL (solid circle), 7.33 logTCID50/mL (empty circle), 6.19 logTCID50/mL (inverted triangle), and 5.35 logTCID50/mL (empty triangle).
It is clear that the heat inactivation procedure recommended by the WHO for DENV is not sufficient for the complete inactivation of infectious chikungunya virions. The minimal time required for the complete heat inactivation was further determined with inoculum at two different average viral titers, 7.66 logTCID50/mL and 4.52 logTCID50/mL, and summarized in Figure 2. In the group receiving 7.66 logTCID50/mL of initial input viral titer, the residual average titers were 3.81 logTCID50/mL, 1.11 logTCID50/mL, and 1.00 logTCID50/mL after 40, 80, and 120 min from the start of heat inactivation. No infectious virions were detected after 160 min of heat inactivation. With the average initial viral titers at 4.52 logTCID50/mL, no infectious virions were detected after 40, 80, 120, and 180 min of heat inactivation.
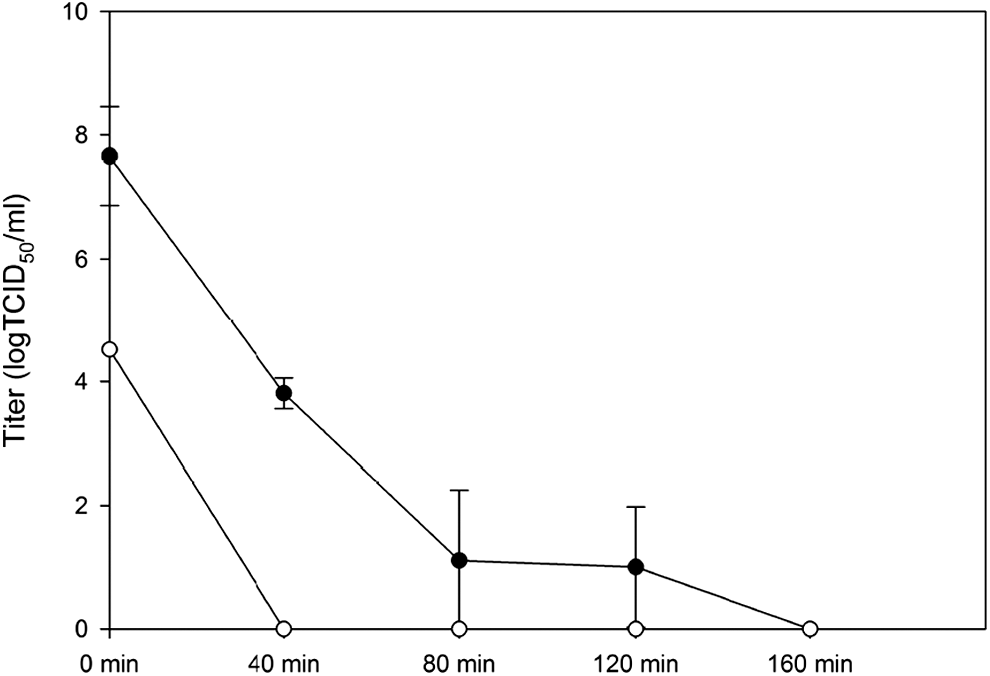
Determination of minimal incubation time required for complete heat inactivation of chikungunya virus (CHIKV). Two aliquots with different titers were tested in this experiment, 7.66 log 50% tissue culture infectious dose (TCID50)/mL (solid circle) and 4.52 logTCID50/mL (empty circle).
Discussion
These results and those of others indicate that the routinely used heat inactivation procedure that is effective for flaviviruses fails to achieve the complete inactivation of CHIKV and WEEV. On the basis of the results of this study, it is reasonable to conclude that such insufficient inactivation is likely to be common for at least the members of Semliki Forest serological complex and WEEV serological complex regardless of the biological differences, such as the choice of vector species, the preference of vertebrate hosts, and the symptoms in infected humans. It is obvious that a longer heat-inactivation procedure should be recommended for the laboratory diagnosis of CHIKV and possibly other alphaviruses to ensure the elimination of false-negative results in neutralization tests and to improve safety levels in the laboratory. This is especially important given the sympatric distribution of CHIKV with other viruses, including DENV, and the concurrent circulation in many endemic areas.
Although relatively higher temperatures have been reported to inactivate CHIKV and other arboviruses, heat inactivation at 56°C of serum samples remains the most acceptable practice in the laboratory due to the denaturation and gelation of plasma proteins, which commences at 55°C and increases at 60–65°C (Vermeer and Norde 2000, Gorinstein et al. 2002, Pialoux et al. 2007). The incomplete heat inactivation of CHIKV in serum or plasma samples might potentially create false-negative results in neutralization tests by the unintentional addition of infectious virions that saturate the neutralizing activities of antibodies.
An appropriate control for eliminating such confounding factors is to perform serum-only controls (Fang et al. 2009). The incorporation of a serum-only control into the neutralization tests is particularly important for serological diagnosis performed in areas where CHIKV co-circulates with DENV, which has been previously found sensitive to heat inactivation at 56°C for 30 min, to ensure the more heat-resistant CHIKV is not present as residual viruses in the samples. Several of the published seroepidemiological studies of CHIKV adopted neutralization methods from DENV and performed heat inactivation at 56°C for 30 min (Kuniholm et al. 2006, Ayu et al. 2010, Nakgoi et al. 2014, Kam et al. 2015). Our results also highlight the possibility that the prevalence of CHIKV from the results of these studies can potentially be underestimated due to the false-negative results generated by the failure of removing residual viruses after heat inactivation. More importantly, the current heat inactivation protocol for the serological diagnosis of alphaviruses should be modified on the basis of the higher thermostability of the viruses.
Footnotes
Author Disclosure Statement
Stephen Higgs is the Editor-in-Chief of Vector-Borne and Zoonotic Diseases. No conflicts of interest exist for the remaining authors.