
Abstract
Candidatus Bartonella mayotimonensis was detected in 2010 from an aortic valve sample of a patient with endocarditis from Iowa, the United States of America. The environmental source of the potentially new endocarditis-causing Bartonella remained elusive. We set out to study the prevalence and diversity of bat-associated Bartonella in North America. During 2015, mist nets and harp traps were used to capture 92 bats belonging to two species: little brown myotis (Myotis lucifugus Le Conte 1831, n = 73) and the gray myotis (M. grisescens A.H. Howell 1909, n = 19) in Kentucky, Michigan, Pennsylvania, and Tennessee. DNA preparations of peripheral blood samples from bats were subjected to a three-marker (gltA, rpoB, and intergenic spacer region [ISR]) multilocus sequence analysis. Sequence-verified gltA-positive PCR amplicons were obtained from nine samples. Three sequences were 99.7–100% identical with the gltA sequence of the Iowa endocarditis patient strain. Analysis of rpoB and ISR sequences demonstrated that one little brown myotis sample from the Upper Peninsula of Michigan contained Bartonella DNA, with 100% sequence identity with the Iowa endocarditis patient strain DNA. It appears possible that bats are a reservoir of Candidatus Bartonella mayotimonensis in North America.
Introduction
B
Veikkolainen et al. (2014) recently detected and isolated Bartonella strains from peripheral blood samples of Daubenton's myotis (Myotis daubentonii Kuhl 1817) in Southwestern Finland. Multilocus sequence analysis (MLSA) demonstrated that rpoB and ftsZ markers of the isolated strains had up to 96–97% sequence identity with the Iowa Candidatus B. mayotimonensis patient strain sequences (Veikkolainen et al. 2014). However, one frequently used Bartonella MLSA marker, the gltA (Norman et al. 1995), displayed low values of sequence identity (91.4–92.9%) (Veikkolainen et al. 2014). This casts doubt on the possibility that the Bartonella isolates from Finnish bats were strains of Candidatus B. mayotimonensis.
Here, we set out to study the prevalence and diversity of bat-associated Bartonella in North America, where Candidatus B. mayotimonensis was first detected from the endocarditis patient sample.
Materials and Methods
During 2015, mist nets and harp traps were used to capture bats at hibernacula or summer roosts in Kentucky, Michigan, Pennsylvania, and Tennessee. Ninety-two bats belonging to two species, the little brown myotis (Myotis lucifugus Le Conte 1831, n = 73) and gray myotis (M. grisescens A.H. Howell 1909, n = 19), were included in this study (Fig. 1 and Supplementary Table S1; Supplementary Data are available online at
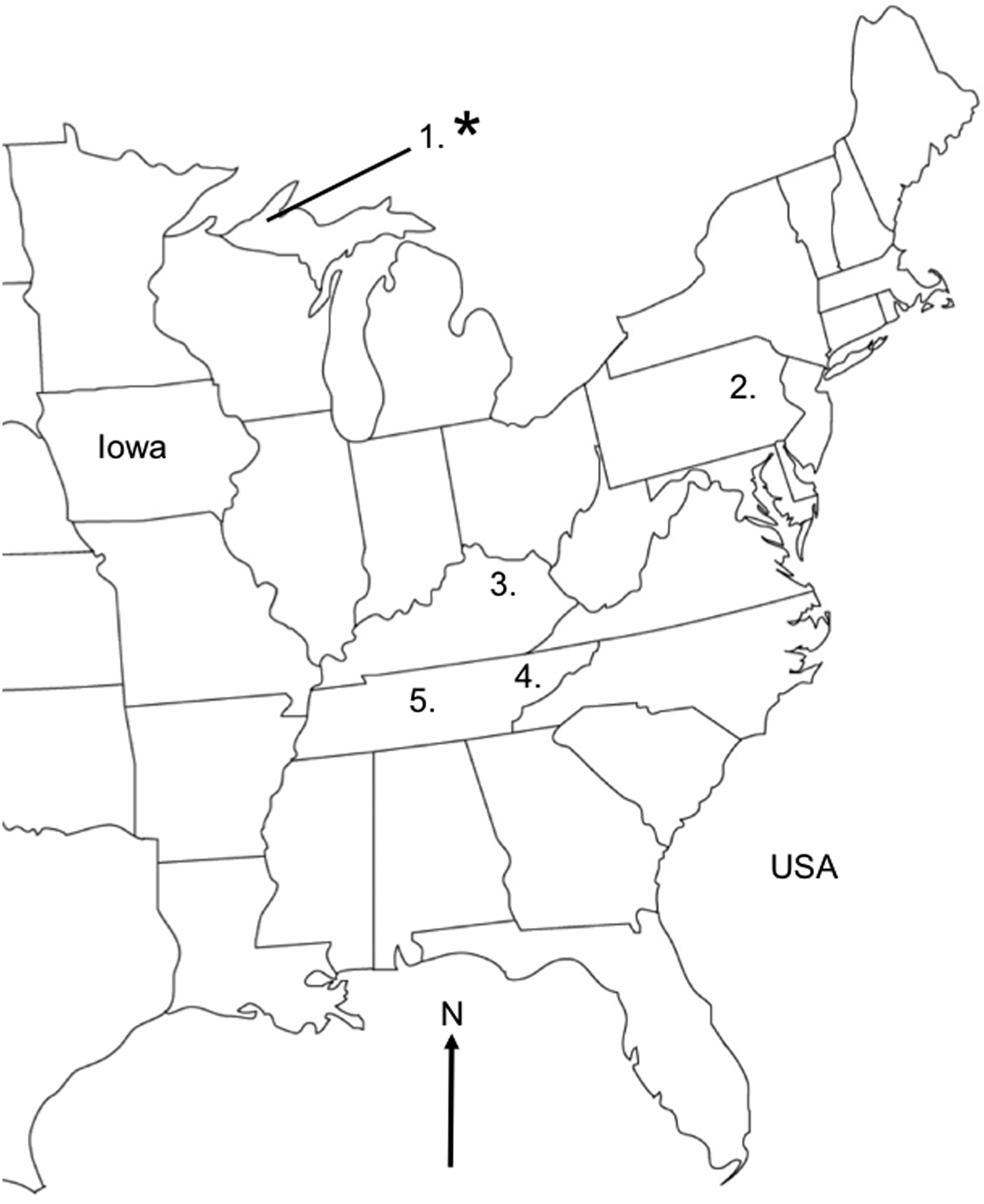
Study locations in the eastern United States of America. Bats were captured in five locations. Asterisk refers to the location where strains of Candidatus Bartonella mayotimonensis were detected. The initial endocarditis patient with Candidatus B. mayotimonensis infection was diagnosed in the state of Iowa (Lin et al. 2010). Numbers after locations represent Bartonella-positive bats/total number of bats caught: (1) Greenland, Upper Peninsula, Michigan (7/43); (2) Allentown, Pennsylvania (0/24); (3) Frankfort, Kentucky (0/6); (4) Warren County, Tennessee (1/7); (5) Montgomery County, Tennessee (1/12).
All methods were approved by the Institutional Animal Care and Use Committee at Bucknell University (protocol DMR-016). The bats were collected under Scientific Collector's Permits from Michigan (SC1475) and Kentucky (SC1411147), Pennsylvania Game Commission Special Use Permit (permit #33085), Tennessee Wildlife Resource Agency Scientific Collection Permit (#3742), and USFWS permit # TE35313B.
DNA was isolated from 100 μL of the PBS-diluted packed cell sample by using a QIAamp DNA mini kit (Qiagen; 51304) for a final elution of 60 μL. The rest of the packed cell samples (200 μL) were used for bat ecophysiology studies that are neither related to nor reported in this study. Therefore, we did not make attempts to cultivate Bartonella strains.
Subsequently, 5 μL of the isolated DNA solution was subjected to a master mix-based PCR protocol that was specific for the citrate synthase gene (gltA) (Norman et al. 1995), for the RNA polymerase B-subunit gene (rpoB), and for the 16S-23S rRNA intergenic spacer region (ISR) (García-Esteban et al. 2008) with DyNAzyme II DNA polymerase (Thermo Fischer Scientific; F501S) (Table 1). Primers specific for rpoB (Table 1) were custom-designed to match the sequence of the strain from the patient from Iowa (Lin et al. 2010), and they are applicable, for example, as a nested oligo pair when the frequently used rpoB-specific 1400F/2300R oligo pair (Renesto et al. 2001) has been utilized in primary PCR. Distilled water and 10 ng of chromosomal DNA of B. quintana strain Toulouse were used as the negative and positive controls, respectively.
Results
Sequence-verified gltA-positive PCR amplicons (GenBank KX807171–KX807179) were obtained from nine samples (Supplementary Table S1). Searches using BLAST (August 2016) with the gltA sequences demonstrate that samples from three little brown myotis from the Upper Peninsula of Michigan contained DNA of Bartonella that was most similar (97.5%) to DNA of Bartonella previously detected in a bat fly, Trichobius corynorhini, in the United States of America (GenBank JX41623; unpublished). One sample from the little brown myotis from the Upper Peninsula of Michigan and one from the gray myotis from Tennessee contained DNA of Bartonella that was most similar (98.0%) to DNA of Bartonella previously detected in Costa Rican bats (GenBank KJ816689; unpublished). One sample from a gray myotis from Tennessee contained DNA of Bartonella that was only distantly related to any entry in GenBank. This sample contained DNA that was closest (91%) to the DNA of Bartonella DNA that was previously detected in samples from Costa Rican bats (GenBank KJ816665; unpublished).
Remarkably, blood from three little brown myotis from the Upper Peninsula of Michigan contained DNA of Bartonella, with up to a 100% sequence identity with the strain of Candidatus B. mayotimonensis from Iowa (Lin et al. 2010) (Table 2). We further subjected these three samples to master mix-based PCR protocols that were specific for the rpoB and ISR (García-Esteban et al. 2008) (Table 1). Analysis of rpoB and ISR sequences (GenBank KX807182–KX807187) demonstrated that one sample from a little brown myotis from the Upper Peninsula of Michigan contained DNA of Bartonella, with up to a 100% sequence identity with the strain of Candidatus B. mayotimonensis from the patient in Iowa (Lin et al. 2010) (Table 2).
ISR, intergenic spacer region.
Discussion
Based on our results, we suggest that bats are a reservoir of Candidatus Bartonella mayotimonensis in North America. We found out that the gltA, rpoB, and ISR MLSA marker sequences of some Bartonella strains associated with North American bats are 100% identical with the Bartonella detected in the patient with endocarditis from Iowa, the United States of America (Lin et al. 2010). This patient lived on a farm, and it is possible that this person could have come into contact with bats roosting in one or several farm buildings. The little brown myotis often roosts in attics or barns during summer, and the size of their maternity colonies can reach hundreds of individuals. During weaning, pups often stray inside houses and may come into contact with humans.
One possible transmission mechanism of hemotrophic Bartonella from bats to humans may be bat ectoparasites or their feces. We have detected Bartonella gltA sequences (GenBank KX807180–KX807181) in Canada from bat-associated Cimex adjunctus (unpublished). These sequences were most similar (98.2% and 97.3%) to DNA of Bartonella previously detected in Costa Rican bats (GenBank KJ816689; unpublished) and Bartonella DNA previously detected in a bat fly Trichobius corynorhini in the United States of America (GenBank JX416236; unpublished). A previous report describes the detection of Bartonella sp. DNA in one bat bug, C. adjunctus, from South Carolina, the United States of America (Reeves et al. 2005). The presence of human blood in an engorged bat tick, Carios kelleyi, from Iowa has also been reported (Gill et al. 2004), indicating that this bat ectoparasite may occasionally feed on human blood. Remarkably, Bartonella sp. DNA has been detected in C. kelleyi in Iowa, the United States of America (Loftis et al. 2005).
More studies on the presence of viable Bartonella in bat-associated ectoparasites are needed to critically evaluate whether bat ectoparasites or their feces are involved in the transmission of hemotrophic Bartonella to humans. Moreover, the cultivation of North American isolates of Candidatus Bartonella mayotimonensis from bat peripheral blood samples should be attempted, for example, similar to our previous study conducted in Finland (Veikkolainen et al. 2014). The isolated strains would allow detailed multilocus sequence analytics, genome sequencing, as well as the development of serological tests for the assessment of the clinical importance of Candidatus Bartonella mayotimonensis.
Following the pioneering work by the Birtles laboratory in the United Kingdom (Concannon et al. 2005), our group in Finland (Veikkolainen et al. 2014, Lilley et al. 2015), among others, has shown that a diversity of Bartonella genotypes are associated with bats and their ectoparasites on a global scale, including Kenya (Kosoy et al. 2010), Guatemala (Bai et al. 2011), Peru (Bai et al. 2012), and Taiwan (Lin et al. 2012). In fact, the non-Candidatus B. mayotimonensis gltA sequences detected in this study have less than 95% sequence identity with any of the Bartonella type strain gltA sequences. This indicates that bats in North America may host a plethora of new species of Bartonella. Given the possible link between bat-associated Bartonella and endocarditis, more studies similar to Mannerings et al. (2016) are needed to assess the zoonotic potential of bat-associated Bartonella.
Footnotes
Acknowledgments
Tennessee Wildlife Resource Agency: Josh Campbell, Daniel Istvanko, Chris Ogle, Chris Simpson, and Dustin Thames; University of Tennessee: Grace Carpenter, Scott Hollis, Devin Jones, Reilly Jackson, and Mallory Tate; Clemson University: Leanne Burns. The authors also thank Craig Willis and members of the University of Winnipeg bat lab that helped collect samples.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.