
Abstract
Central Queensland (CQ) is a large and isolated, low population density, remote tropical region of Australia with a varied environment. The region has a diverse fauna and several species of ticks that feed upon that fauna. This study examined 518 individual ticks: 177 Rhipicephalus sanguineus (brown dog tick), 123 Haemaphysalis bancrofti (wallaby tick), 102 Rhipicephalus australis (Australian cattle tick), 47 Amblyomma triguttatum (ornate kangaroo tick), 57 Ixodes holocyclus (paralysis tick), 9 Bothriocroton tachyglossi (CQ short-beaked echidna tick), and 3 Ornithodoros capensis (seabird soft tick). Tick midguts were pooled by common host or environment and screened for four genera of tick-borne zoonoses by PCR and sequencing. The study examined a total of 157 midgut pools of which 3 contained DNA of Coxiella burnetii, 13 Rickettsia gravesii, 1 Rickettsia felis, and 4 other Rickettsia spp. No Borrelia spp. or Babesia spp. DNA were recovered.
Introduction
C
The regional and rural nature, tropical climate, and remoteness of CQ place those living in the region at risk for zoonotic tick-borne diseases. The known tick-borne diseases of humans in the CQ region are Q fever (Coxiella burnetii) and Queensland tick typhus (Rickettsia australis) (Parola et al. 2013). However, multiple species of tick, as well as diverse and abundant wildlife are present in the region, which may act as undetected reservoirs of emergent tick-borne diseases described elsewhere in Australia. Pathogens such as Rickettsia honei subsp. marmionii have been detected in regions of Queensland north of CQ (Parola et al. 2013), while the putative human pathogen (Brown et al. 2001, Arraga-Alvarado et al. 2014), Anaplasma platys, has previously been reported in Rhipicephalus sanguineus ticks taken from domestic dogs in inland desert regions of the Northern Territory (Brown et al. 2001), west of the CQ region.
In Australia, there is controversy over the potential presence of two emergent tick-borne diseases, babesiosis and Lyme borreliosis. In 2012, a fatal, apparently locally acquired, case of a Babesia microti infection was reported from a patient in Canberra (Sanjaya et al. 2012). Babesia spp. of veterinary and agricultural importance is known to be present in Australia (Bock et al. 1999, Greay et al. 2016). B. microti has not been found in animals or ticks in Australia, nor have further human cases occurred. There are growing anecdotal reports of a “Lyme-like disease” manifesting following tick bites in Australia, including individuals in the CQ region (Chalada et al. 2016). While Borrelia burgdorferi s.l. are common tick-borne pathogens in the northern hemisphere, there is scant evidence supporting their presence in Australia (Chalada et al. 2016, Collignon et al. 2016). Investigations for Lyme-causing Borrelia in Australia have until now been limited to more densely populated southern coastal regions (Russell et al. 1994, Gofton et al. 2015a, 2015b, Graves et al. 2016). Sequences of a novel Borrelia species related to the Borrelia recurrentis group, Candidatus Borrelia tachyglossi, have been detected in Bothriocroton concolor ticks from Queensland echidnas, but the pathogenic potential of this organism remains unknown (Loh et al. 2016, 2017). Due to its remote and underpopulated nature, no survey of tick-borne pathogens in CQ has been previously published in the scientific literature.
Materials and Methods
Tick collection and identification
Ticks were actively collected from animals and questing ticks were collected by flagging. Live ticks were stored and transported at room temperature to the laboratory within 7 days and upon arrival, frozen at −20°C for 24 h to euthanize the tick. The tick collection catchment area for this study comprised the area between longitude 146° and 151°E, at latitude 24°S (Fig. 1). Five hundred sixteen ticks were collected from this region between January 2015 and November 2016.
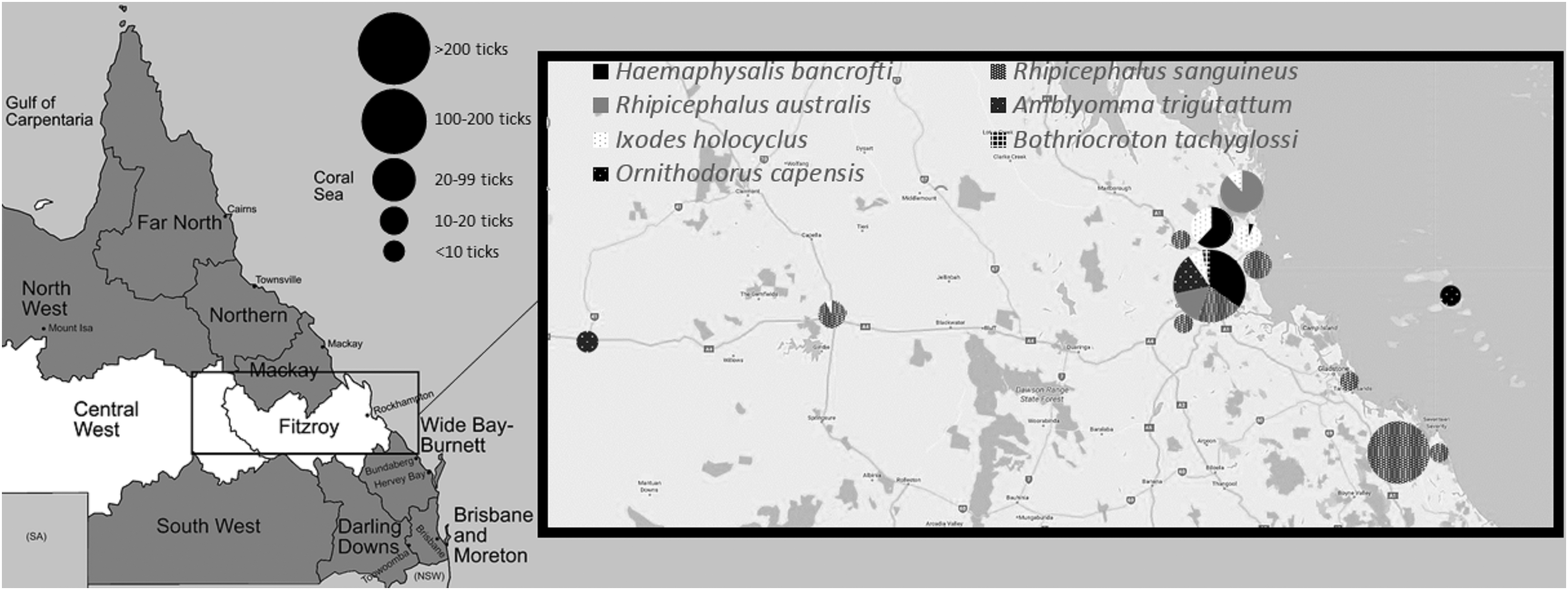
Map of Queensland showing the CQ region (white), study catchment area (box), sites where ticks were recovered (inset), number, and proportion of tick species recovered at that site (pie charts). CQ, Central Queensland.
Tick identification
The source hosts and identification of all ticks included in this study are shown in Table 1. Several references were consulted for tick identification purposes (Roberts 1962, 1970, Andrews et al. 2006, Barker et al. 2014). The identification of the tick gender and life stages followed the criteria described as follows: male adults were identified by the scutum covering the whole dorsal body and the presence of a genital aperture and female adults by the scutum only partially covering the dorsal body, plus the presence of a genital aperture. Nymphs were identified by the scutum partially covering them, but not having a genital aperture. Larvae were identified by possession of only six legs; species was based on what the concurrent species of nymphs and adults on the same animal at the same time was. No host animal harbored more than one species of tick. Ticks recovered were as follows: 177 R. sanguineus, 123 Haemaphysalis bancrofti, 102 Rhipicephalus australis, 47 Amblyomma triguttatum, 57 Ixodes holocyclus, 9 Bothriocroton tachyglossi, and 3 Ornithodoros capensis. Pooling of midgut samples into groups by identical tick species and common individual host animal resulted in a total of 157 pooled tick midguts being tested. Two ticks from outside of CQ catchment area were processed: a Haemaphysalis humerosa from a bandicoot (species not determined, either Perameles spp. or Isoodon spp.) from North Queensland and an A. triguttatum from the environment in the Darling Downs/South West region.
Sub-species not determined.
Tick dissection
Ticks were thawed, dipped, and agitated in a 10% hypochlorite solution for 3 s and then agitated in 70% ethanol for 3 s. Tick mid-guts were removed aseptically and stored at −80°C. Midguts of ticks of the same species from the same animal were pooled, with a maximum of 10 tick midguts per pool. Tick exoskeletons were stored in ethanol and assigned a unique identifying number in the Barker Laboratory collection at the University of Queensland (Table 1).
DNA extraction
DNA extraction was performed using the PowerSoil DNA isolation kit (MO BIO Laboratories, Inc., Carlsbad, CA). A sterile distilled water control was included in each extraction batch. All DNA extracts were subjected to a tick 18S rRNA gene PCR (Black and Piesman 1994) as a DNA extraction and PCR inhibition control.
PCR targets and reaction conditions for Borrelia and Babesia
Babesia spp. detection was by an 18S rRNA gene PCR (Hilpertshauser et al. 2006), detecting the Babesia genus, with extracted DNA from a veterinary isolate of Babesia gibsoni used as a positive control. Borrelia spp. detection was by two genus-specific PCRs targeting the Borrelia 16S rRNA gene (Rys 1993) and OspC gene (Crowder et al. 2010). DNA extracted from cultures of B. burgdorferi, Borrelia afzelii, and Borrelia garinii were used as controls. Conventional PCR mixtures contained 0.5 μM each of forward and reverse primer (Geneworks, Adelaide, SA) with a total reaction volume of 25 μL. Thermocycler conditions were; initial denaturation at 95°C for 10 min; then 40 cycles of 95°C for 30 s; 50°C (tick 18S rRNA and Babesia 18S rRNA) or 45°C (Borrelia 16S rRNA and OspC) for 30 s; and 72°C for 35 s, with final extension at 72°C for 7 min. Products were visualized by agarose gel electrophoresis.
PCR targets and reaction conditions for Rickettsia and Coxiella genes
Real-time PCR was performed using a RotorGene Q (Qiagen, Germantown MD) cycler. Targets were the Rickettsia citrate synthase (gltA) gene (Stenos et al. 2005), the Coxiella heat shock (htpAB) operon (Bond et al. 2016), and the C. burnetii outer membrane protein (com1) gene (Lockhart et al. 2011). PCR was run with 400 nM each of forward and reverse primer and 200 nM of probe (Biosearch Technologies, Buenos Aires, Argentina). Extracted DNA of clinical C. burnetii, Rickettsia conorii, and Rickettsia typhi isolates was used as controls. Cycler conditions were as follows: initial hold at 50°C for 3 min, second hold at 95°C for 5 min, and 40 cycles of denaturation at 95°C for 20 s with annealing/extension at 60°C for 40 s. Samples were considered positive if their fluorescence passed the threshold (0.02) in less than 38 cycles and equivocal between 38 and 40 cycles.
Samples with an equivocal or positive real-time PCR result for the Rickettsia gltA gene were subjected to a conventional hemi-nested PCR for sequencing using primers targeting the Rickettsia gltA gene and the Rickettsia 17 kDa antigen gene (Ishikura et al. 2003) with 0.5 μM each of forward and reverse primers (Geneworks) in a 25 μL reaction. Cycler conditions were as follows: initial denaturation at 95°C for 5 min, 40 cycles of denaturation at 95°C for 30 s, annealing for 30 s, and extension at 72°C for 90 s, followed by a final extension at 72°C for 7 min. The first of the hemi-nested PCRs for both targets used an annealing temperature of 44°C and the second reactions used an annealing temperature of 48°C. All primer and probe sequences are shown in Table 2.
F, forward primer; P, probe; R, reverse primer.
Sequencing and analysis
DNA bands from the conventional PCRs were cut from the agarose gel using sterile disposable scalpel blades and purified using the FavorPrep Gel/PCR Purification Kit (Favorgen, Taiwan) according to the manufacturer's protocol. Purified DNA was stored at −20°C until use.
Sequences of the Rickettsia 17 kDa and gltA products were determined by Sanger sequencing at the Australian Genome Research Facility, Melbourne. Forward and reverse sequences were then formed into contiguous sequences using the HeracleBioSoft DNA Baser v4.16.0.25. Subsequent sequences were analyzed for identification using the NCBI BLAST and phylogenetic relationships in comparison to published Rickettsia 17 kDa and gltA sequences carried out using MEGA7 software (Kumar et al. 2016). The neighbor-joining algorithm was used to generate the initial tree. Evolutionary history was inferred using the minimum evolution method with a bootstrap consensus tree inferred from 500 replicates. Branches corresponding to partitions reproduced in less than 50% bootstrap replicates were collapsed. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (500 replicates) is shown next to the branches. Evolutionary distances were computed using the maximum composite likelihood. The molecular evolution tree was searched using the close-neighbor-interchange algorithm at a search level of 1. Codon positions included were first+second+third+noncoding. All positions containing gaps and missing data were eliminated.
Ethical approval
Ethical approval for the removal of ticks from live animals was provided by the CQ University Animal Ethics Committee (approval no. A15/12-342).
Results
Several potential tick-borne zoonoses were identified in ticks included in this study (Table 3). Of the CQ tick midgut pool samples, three tick pooled midgut samples (2%) were positive for both the com1 and htpAB targets. A further three samples (2%) were positive and one equivocal for the com1 gene only. Twenty samples (13%) were positive by real-time PCR targeting Rickettsia gltA. A further seven yielded results in the equivocal range. Due to lower test sensitivity, subsequent conventional hemi-nested PCR revealed 12 high-quality gltA sequences and 16 17 kDa sequences. Phylogenetic analysis of these sequence products is summarized in two phylogenetic trees (Fig. 2). All yielded products in the tick 18S rRNA PCR extraction and inhibition control assay.
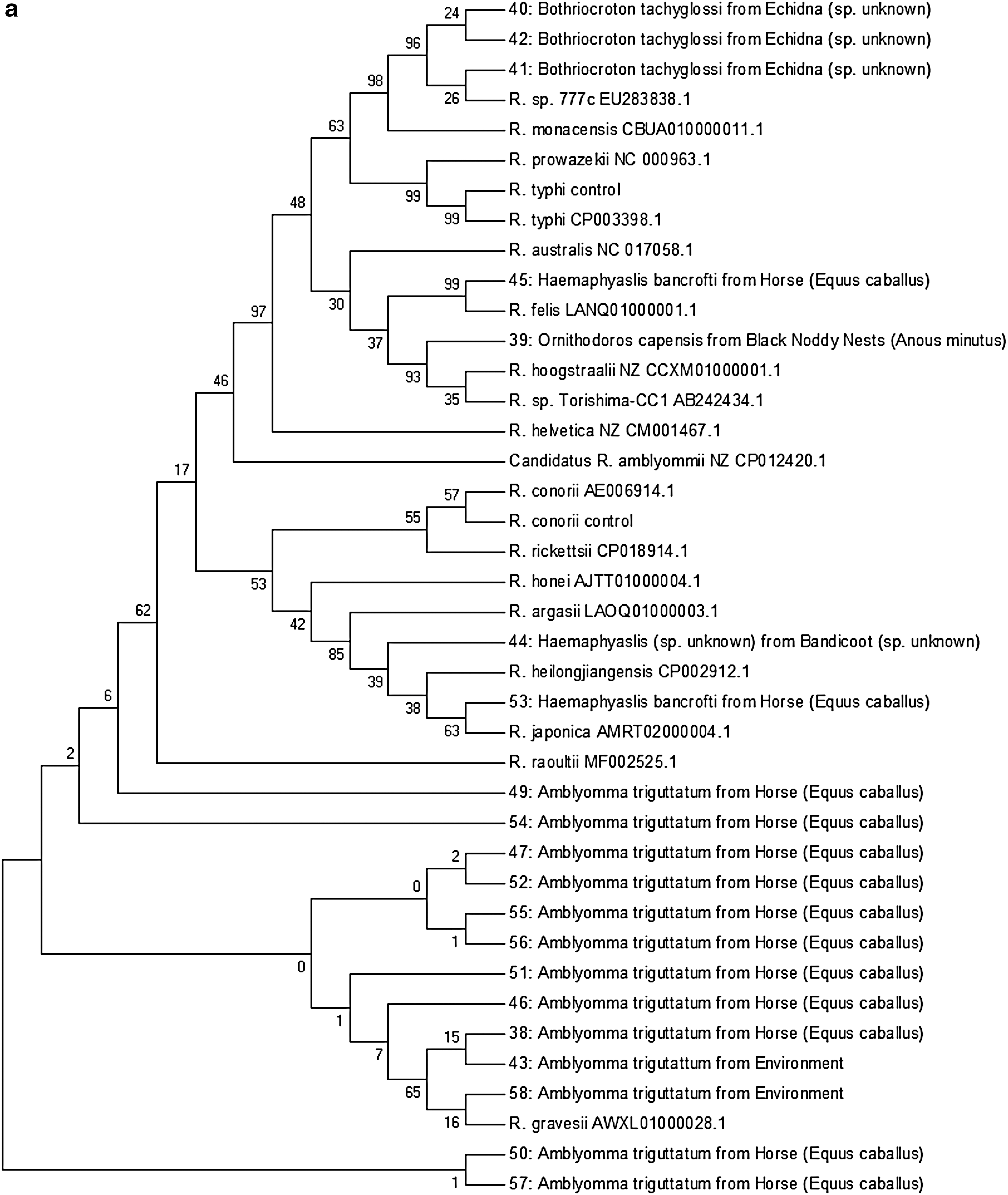
Bootstrapped neighbor-joining depiction of the evolutionary relationships of (

R. 17 kDa PCR and, where necessary, gltA, products were sequenced to allow definitive identification of species detected.
−14 bp ins: a 14 bp insert was removed from sequence before comparative sequence analysis.
Except where stated, the taxonomic species of where host animal ticks were drawn from was not determined.
Coxiella-like symbiont is only speculation based on only one of the two Coxiella burnetii gene targets (htpAB and 17 kDa) testing positive in PCR. This suggests that these organisms were not C. burnetii, but are something closely related in phylogeny.
These ticks were collected from separate horses at the same stable in Rockhampton.
Unable to determine subspecies of nymph stages.
nd, not determined.
The 17 kDa PCR product of a pool of three midguts of O. capensis shared 409/409 bp (100%) with the 17 kDa gene sequence of Rickettsia sp. Torishima CC1, described by Kawabata et al. (2006) and that of Rickettsia hoogstraalii, described by Andoh et al. (2015). A gltA sequence was not obtained in this instance. A pool of six midguts from H. bancrofti feeding on a horse in Rockhampton yielded an identical sequence to Rickettsia felis 17 kDa; a gltA sequence could not be obtained. The midgut pool from six H. bancrofti ticks removed from a horse at a different Rockhampton stable and a H. humerosa removed from a bandicoot in Port Douglas yielded gltA and 17 kDa sequences very similar to Rickettsia japonica, Rickettsia raoulti, and Rickettsia argasii. These two unknown rickettsial species shared 329/332 bp (99.1%) of their gltA sequences and 407/409 bp (99.5%) of their 17 kDa sequences. Full identification could not be achieved.
Two midgut samples from A. triguttatum ticks, recovered by flagging, in Rockhampton and Chinchilla had 17 kDa and gltA sequences identical to the published sequences of Rickettsia gravesii. An A. triguttatum from a horse in Alpha also had 100% 17 kDa sequence homology to R. gravesii (a gltA sequence was not obtained for this sample). Ten midgut pools of A. triguttatum ticks parasitizing 10 horses at the same stable in Rockhampton yielded sequences very similar to R. gravesii. All samples had 99.8% 17 kDa homology to R. gravesii. Eight showed 99.7% gltA homology, a ninth sample showed 99.4% homology, and a 10th sample yielded a gltA sequence of too poor quality for analysis.
Sequences 40, 41, and 42 generated from 17 kDa PCR of the midgut pools of three B. tachyglossi ticks taken from one echidna in Rockhampton were identical to each other and most closely related to Rickettsia spp. 777b, 774c, and 774e, previously described in Amblyomma fimbriatum ticks removed from a yellow spotted monitor lizard (Varanus panoptes) in the Northern Territory of Australia (Vilcins et al. 2009). The 17 kDa products each contained an extra 14 bp insertion toward the center of the sequence that no other Rickettsia species possesses. Attempts to match this small 14 bp insert sequence with other Rickettsia species sequences were unsuccessful. When the extra 14 bp insertion was excluded, the 17 kDa sequence was otherwise identical to Rickettsia sp. 777c and 99.3% homologous with Rickettsia monacensis. A gltA sequence was yielded by only one sample (42), which shared 100% homology with both Rickettsia sp. 774e and Rickettsia sp. 774b.
PCRs for Borrelia genus (16s rRNA and ospC) and Babesia genus (18s rRNA) were all negative. All controls reacted as expected. The PCR for the tick 18s rRNA gene was positive for all samples, demonstrating effective DNA extraction and the absence of inhibitors.
Discussion
This project aimed to investigate the prevalence of tick-borne pathogens in CQ, an area of Australia with a dry tropics climate and a sparse, regional or rural population. No Borrelia or Babesia spp. was identified in the ticks tested. C. burnetii and several known or potential zoonotic rickettsial pathogens were identified.
A tick midgut sample was only considered containing the species C. burnetii if both C. burnetii PCR targets (com1 and htpAB) were positive. Three tick midgut pools from I. holocyclus, H. bancrofti, and R. sanguineus tested positive for C. burnetii. Although I. holocyclus presents as a vector, which may readily transmit this pathogen to humans, Rhipicephalus sanguineus, only rarely feed on humans, thus exposure of humans to C. burnetii by these latter two vectors is unlikely. The finding of C. burnetii in an I. holocyclus removed from a laughing kookaburra chick (species undetermined) was unexpected. C. burnetii DNA has been detected in the blood of a number of other native animals in Queensland (Cooper et al. 2013) and previous cases of C. burnetii in multiple species of birds have been reported (Stein and Roult 1999, Psaroulaki et al. 2014), but none from any kookaburra species. This finding may represent true acquisition of C. burnetii from the kookaburra host, though it remains possible that the tick was infected while feeding on a previous host of a different species.
A limitation in the methodology of this study was that it was not recorded if ticks were engorged or flat when their midguts were removed. The absence of this information does not allow for determination whether the pathogen has survived transstadially, better evidencing that the tick species concerned is a true vector for the pathogen recovered and not just present in the blood meal from a reservoir animal. Furthermore, individual tick salivary glands were not tested separately to midguts; Rickettsia, Coxiella and Borrelia have been found to move freely between the midgut and salivary glands (Jasinskas et al. 2007, Coumou 2016, Martins et al. 2017) and thus, this limitation in sampling technique should not have significantly lowered recovery of those pathogens. However, in cases where a pathogen was recovered from a tick that is not already a confirmed vector, the true relationship between the pathogen and vector cannot be established from the data presented in this study.
Midguts of H. bancrofti removed from a horse in Rockhampton also tested positive for C. burnetii. C. burnetii has not previously been associated with H. bancrofti ticks and this may represent a C. burnetii-like symbiont. It has been found previously that H. bancrofti harbors Coxiella-like symbionts related closely to those found in Haemaphysalis lagrangei and Haemaphysalis longicornis (Gofton et al. 2015a). Two tick midgut pools from I. holocyclus and A. triguttatum were positive and one equivocal for the com1, but negative for htpAb PCR. Only pooled midgut sample was positive for htpAB PCR alone, from O. capensis in black noddy (Anous minutus) nests on Heron Island. Since these were only positive for one of the two gene targets, it is therefore hypothesized that these represent Coxiella symbiont organisms that naturally occur in a variety of tick genera; indeed, it is thought that C. burnetii originated from a tick Coxiella symbiont ancestor (Duron et al. 2014). Such symbionts may be falsely identified as C. burnetii (Duron et al. 2014, 2015). No R. sanguineus was found to harbor C. burnetii in previous work by Oksam et al. {2017), which reported the absence of this pathogen in R. sanguineus from dogs in Australia, despite high C. burnetii antibody seroprevalence in Australian dogs.
Thirteen samples in this study contained Rickettsia identified as R. gravesii. These were obtained from the midguts of A. triguttatum tick pools from 10 different horses in the sample Rockhampton stable, 1 A. triguttatum from a horse in Alpha, 1 A. triguttatum from the vegetation in Chinchilla, and 1 A. triguttatum from vegetation in Rockhampton. R. gravesii is a spotted fever group Rickettsia (Owen et al. 2006) closely related to Rickettsia massiliae, a human pathogen in Europe and Africa (Sentausa 2013). R. gravesii has been found at very high prevalence in A. triguttatum ticks from Western Australia and this tick species and is thought to represent the main reservoir of the organism (Li et al. 2010). Previous reports of R. gravesii in the scientific literature only describe its presence in Western Australia (Owen et al. 2006, Li et al. 2010, Sentausa et al. 2013). This is the first time that R. gravesii has been reported in Queensland.
R. felis was recovered from the pooled midguts of six H. bancrofti ticks taken from a horse in Rockhampton. Rickettsia 17 kDa sequencing performed on this isolate showed 100% identity to R. felis, a cause of spotted fever worldwide (Pérez-Osorio et al. 2008), which has been detected previously in Australia, including Queensland (Barrs et al. 2010, Hii et al. 2013). The primary vector of R. felis is the cat flea, Ctenocephalides felis (Pérez-Osorio et al. 2008). R. felis has never been detected in Australian ticks to date, but was detected in Haemaphysalis flava, Haemaphysalis kitaokai, and Ixodes ovatus in Japan (Ishikura et al. 2003) and other tick genera in Brazil (Cardoso et al. 2006) and South Carolina (Durden et al. 1999). Therefore, its presence in an Australian Amblyomma tick is not unexpected, although the equine host from which it was obtained is most unusual. The paddock in which the horse was stabled was close to residential areas where many pet and feral cats roam. It is speculated that the A. triguttatum obtained from the horse had previously fed from a cat host and the source of R. felis was this prior host.
Midgut pools of O. capensis were also found to contain Rickettsia spp. The 17 kDa sequence of Rickettsia spp. Torishima CC1 (GenBank: AB242434.1) was first detected in O. capensis ticks of Japan (Kawabata et al. 2006) and has been isolated from O. capensis ticks of Georgia (Mattila et al. 2007), and in Amblyomma transversale removed from a snake that was imported to Japan from Ghana (Andoh et al. 2015). Rickettsia spp. Torishima CC1 shares high identity in the 17 kDa gene with R. hoogstraalii (GenBank: Z_CCXM01000001.1) with base pairs 409/409 (100%) matching in both the work presented in this study and in the research of Andoh et al. (2015), but requires confirmation by further sequencing. R. hoogstraalii has also been isolated from Haemaphysalis sulcata from Croatia (Duh et al. 2010). The pathogenicity of R. hoogstraalii is not yet apparent in the scientific literature, but certainly appears to have a worldwide distribution with a number of possible vector tick species. These findings are consistent with the worldwide distribution of the tick O. capensis and its host birds (Dietrich et al. 2011).
A Rickettsia thought to be closely related to R. japonica and that clustered with R. japonica, R. raoulti, R. argasii, and Rickettsia heilongjiangensis was found in a pool of six H. bancrofti taken from a horse. A similar, but not identical Rickettsia was found in the midgut of a Haemaphysalis tick removed from a Bandicoot (species undetermined) in Port Douglas. R. japonica is a cause of oriental spotted fever in Japan (Uchida 1993), but exists globally in a variety of Haemaphysalis species, including H. longicornis, H. flava, Haemaphysalis formosensis, and Haemaphysalis hystricis in Japan (Uchida et al. 1995, Fournier et al. 2002), H. longicornis in Korea (Lee 2003), and H. hystricis in Thailand (Takada et al. 2009). H. longicornis is also present in Australia, thought to have originated from Japan (Barker et al. 2014). This tick shares similar hosts to H. bancrofti and therefore it is not impossible that R. japonica may have made its way to Australia and its fauna. Further gene amplification and sequencing are required to confirm whether this Rickettsia is actually R. japonica or only a very closely related organism.
The distribution ticks recovered in this study were unremarkable except for one I. holocyclus taken from a dog in Emerald, 310 km inland. The geographic range of I. holocyclus is considered to be primarily coastal, in areas with at least 1000 mm of rainfall per year (Barker et al. 2014), well above the average received in the Emerald region. However, CQ dogs will often travel with their owners when on recreation or shopping in the larger population centers on the coast, and it should be considered that this tick might have been acquired there and returned to inland Emerald on the host dog. No further information on this dog or its travels could be obtained.
This study did not test for the presence of A. platys in R. sanguineus from CQ and further investigation of the potential presence of this pathogen in that region is warranted. No Borrelia spp. or Babesia spp. was detected in this study. This is despite the known presence of Babesia spp. in Australian dogs and cattle, as well as one suspected locally acquired case of B. microti in a human. In contrast, regions endemic for B. burgdorferi s.l. show up to 30% of Ixodes ticks infected (Cao et al. 2003, Gofton et al. 2015b). It is highly probable that these two genera of pathogens are not present in CQ.
Conclusions
This study surveyed several species of CQ ticks for tick-borne pathogens. Several species of Rickettsia were identified, many of which are known (R. felis and R. gravesii), or potential (R. japonica like, Rickettsia sp. Torishima-CC1, and Rickettsia sp. 777c), zoonoses and all of which had not been previously identified in the CQ region. The detection of C. burnetii in several species of ticks further reinforces the risk of Q fever in this region. No Babesia spp. of medical, veterinary, or agricultural importance were identified, despite the continued presence of B. bigemina and B. bovis in isolated pockets in the remote north of Queensland. No Borrelia spp. was detected, reinforcing the likely absence of Lyme-causing Borrelia spp. in Australia.
Footnotes
Acknowledgments
The authors would like to acknowledge the assistance of Mr. Saba Sinai for his assistance in the collection of ticks from inland CQ and Prof. Stephen Barker of University of Queensland for his assistance with confirmation of tick identifications. The authors thank the laboratory of Prof. Peter Irwin at Murdoch University for the provision of control DNA and technical advice. The authors would also like to thank Mr. Wayne Pederick and his employer, QML Pathology, for their assistance in transport of ticks. This work was funded by a grant from the Graduate Research Office of Central Queensland University.
Author Disclosure Statement
No competing financial interests exist. Richard Bradbury is co-authoring this article in his personal capacity and in his capacity as an adjunct academic at Central Queensland University.