
Abstract
Background:
Surveillance is important in addressing the significant public health concerns posed by tick-borne diseases. However, the southwestern U.S. presents particular challenges due to diverse tick fauna and varied ecologies.
Methods:
From 2021 to 2022, we conducted a partner-based tick surveillance program in Arizona and California to assess the presence of Rickettsia spp. pathogens and species composition of tick vectors.
Results:
A total of 913 ticks was collected, comprising in descending abundance Rhipicephalus sanguineus, Dermacentor similis, Ixodes pacificus (I. pacificus), Argas sp., Otobius megnini, and Haemaphysalis leporispalustris. Arizona submitted predominantly brown dog ticks (90.05% of all ticks from Arizona), while California showed greater tick species richness with five species identified. No Rickettsia rickettsii was detected, but a variety of other Rickettsia spp. was found in ticks from both Arizona and California and included Rickettsia rhipicephali (R. rhipicephali), Rickettsia massiliae, and Rickettsia monacensis-like rickettsial agents of I. pacificus, and two rickettsial organisms that were not identified to species: one Rickettsia montanensis or Rickettsia raoultii-like, and the other most similar to Candidatus Rickettsia tarasevichiae (R. tarasevichiae).
Conclusion:
This research contributes to our understanding of tickborne diseases in the southwestern U.S., and emphasizes the need for targeted surveillance and intervention initiatives in a region with complex relationships among ticks, hosts, and Rickettsia species. In particular, the finding of an apparently novel pairing of an unknown Argas sp. tick and R. tarasevichiae-like organism suggests that argasid species are an important target for future research. In addition, the results—both tick species submitted and resulting Rickettsia spp. identified—highlights the strengths and potential biases associated with a partner-based sampling method for tick surveillance.
Introduction
With increasing numbers of cases of tickborne diseases (Bishop et al., 2022; Eisen and Paddock, 2021), there is a pressing need for surveillance of ticks and the pathogens they transmit. The southwestern U.S. presents an ideal yet challenging landscape for studying tickborne diseases due to abundant tick species and diversity in ecologies. Important ticks in the region include Ixodes pacificus (I. pacificus), Dermacentor similis (D. similis), Rhipicephalus sanguineus (R. sanguineus), Amblyomma maculatum (A. maculatum), and Dermacentor occidentalis (D. occidentalis) (Hecht et al., 2020; Furman and Loomis, 1984).
Each of these ticks can vector gram-negative bacteria in the family Rickettsiaceae. Rickettsiae were some of the first vector-borne diseases discovered (Parola et al., 2005). The genus Rickettsia (R) consists of the spotted fever group (SFG), typhus group (TG), ancestral, and transitional groups of species (Merhej and Raoult, 2011). At least six zoonotic Rickettsia species have been identified along the U.S.–Mexico border, including SFG Rickettsia 364D (“R. phillipii”) transmitted by D. occidentalis, Rickettsia parkeri in A. maculatum, Rickettsia massiliae (R. massiliae) and Rickettsia rickettsia (R. rickettsia) in R. sanguineus (López-Pérez et al., 2021), and transitional group Rickettsia felis (R. felis) disseminated by fleas (Reif and Macaluso, 2009). Rocky Mountain spotted Fever (RMSF) caused by Rickettsia rickettsii (R. rickettsii) is a particularly dangerous threat to human health and is found across North, Central, and South America (Foley et al., 2024a). Before antibiotics were available, the mortality rate for RMSF was 23%, but even now, heavily impacted areas with marginalized populations experience case fatalities as high as 50% (Foley et al., 2024a).
There is a lack of knowledge about Rickettsia-caused disease incidence, geographic distribution, and contributing factors in the southwestern U.S., prompting targeted research and surveillance. In response, the Border Tick Rickettsia Surveillance Program (BiTeRS) was established as a collaboration among the Universities of California and Arizona and stakeholders in California and Arizona. This partnership offered tick collection and identification, pathogen testing, and training and provision of educational resources. The primary focus of the study was to enhance the detection of R. rickettsii to improve RMSF surveillance, with a secondary goal to identify other pathogenic rickettsial species in the region. This study documents tick species obtained during BiTeRS surveillance during 2021–2022, the prevalence of rickettsiae, temporal, spatial, and host characteristics associated with the submissions.
Materials and Methods
Tick collection
From April 2021 to December 2022, we performed a survey working with BiTeRS’ partners to collect ticks and metadata associated with the specimens including location, species of host, whether the tick was engorged, and date of collection. The collection focused on U.S.–Mexico border counties including San Diego and Imperial counties in California and Coconino, Navajo, Pima, Pinal, Maricopa, and Yuma counties in Arizona. Navajo and Coconino, farther north of the international border, were included due to partner requests. Initially, we sought 10 partner organizations (POs) in each state to collaborate including tribes, federal and state agencies (such as Border Patrol, and National Parks), county health departments or vector control agencies, and animal control agencies. Each PO would identify a key liaison point of contact trained in tick identification and surveillance techniques, to help adapt the tick surveillance program to their needs. Candidate organizations within 120 km of the U.S.–Mexico border in California and Arizona were invited and then we used word-of-mouth and professional referral to enroll as many as possible. POs utilized various collection techniques to obtain samples opportunistically, including flagging, dragging, detection of clothing, people, or animals, and manual removal from the environment. Samples were preserved in vials containing 70% ethanol and shipped to the University of California, Davis.
Tick identification and rickettsia spp. detection
Using a dissecting microscope, we identified ticks to life stages and species with a morphological key (Furman and Loomis, 1984). Unengorged ticks were individually minced using a sterile scalpel and DNA from these specimens was extracted with an ammonium hydroxide protocol, which is a very inexpensive protocol that has yielded excellent results in numerous studies as long as there is no blood contamination (Humair et al., 2007). Engorged ticks were halved sagittally to ensure penetration of lysis buffer and, if more than 25 mg due to engorgement, halved again in cross-section with only the anterior portions retained. Both halves were then immersed in ATL buffer (Qiagen, Valencia, CA, USA) and proteinase K solution and incubated at 56°C overnight. This minimized the large blood load in DNA extraction which can contribute to inhibition during PCR. The remaining extraction steps were completed per the DNeasy Blood and Tissue Kit (Qiagen) manufacturer instructions.
Each tick was individually subject to pan-Rickettsia real-time PCR to detect a conserved region within the 133-base pair citrate synthase (gltA) gene (Stenos et al., 2005) of Rickettsia species. However, this method does not yield a positive result for Rickettsia near neighbors, such as Bartonella, Coxiella, Ehrlichia, and Orientia species nor the ancestral group Rickettsia bellii. A positive control (R. felis) and a negative control (saline water) were included in each run. Samples with a cycle threshold (Ct) score <40 and with a characteristic amplification curve were deemed positive.
DNA samples that gave a positive result in the pan-Rickettsia test then underwent a real-time PCR protocol specific to R. rickettsii (Kato et al., 2013). Pan-Rickettsia PCR-positive but R. rickettsii-negative samples were further characterized by conventional PCR and DNA sequencing of portions of the 17 kDa gene (200 base pairs) and the gltA gene used for a subset of those (800 base pairs) (Roux et al., 1996; Fournier et al., 2003; Shapiro et al., 2010). Additional PCR and sequencing were performed if the initial 17 dDa sequence was inadequate to identify the rickettsial agent or, at minimum, to classify it as likely nonpathogenic. In the case of more than 10 ticks positive via real-time pan-Rickettsia PCR originating from a single individual host, 10% were selected for sequencing. When ticks could not be identified definitively by species morphologically (i.e., for immature argasids), analysis of a 460-bp region of the 16S rDNA gene was performed (Black and Piesman, 1994). All sequencing was performed at the University of California Davis DNA Sequencing Facility using the ABI Prism 3730 Genetic Analyzer (Thermo Fisher Scientific, Waltham, MA, USA). Using the Basic Local Alignment Search Tool, sequences were contrasted with those in GenBank. Each gene’s sequences were aligned using Muscle in Mega 11 (Tamura et al., 2021). Trees documenting phylogenetic relationships were built in Mega 11 through maximum likelihood and the Tamura–Nei model.
Data analysis
Data were managed in Excel (Microsoft, Redmond, WA, USA) and analyzed in R version 4.2.2. We adopted a p-value threshold of 0.05 for inferring statistical significance. Univariate logistic regression was performed to look for significant associations between the presence of Rickettsia spp. and season (spring: March–May, summer: June–August, autumn: September–November, winter: December–February), host animal species, whether or not the tick was attached to a host, and tick life stage. Separate models were constructed for each tick species except for rabbit (Haemaphysalis leporispalustris) and spinose ear (Otobius megnini) ticks which were not analyzed due to low sample size. This study was not subject to institutional review board R oversight.
Results
From 2021 to 2022, we assessed 913 ticks from Arizona and California from kennels, parks, people, and other animals provided by 17 POs. Figure 1 employs graduated symbols to represent tick samples’ geographic distribution and count. We observed notable differences between the two states’ annual tick collection numbers. In 2021, Arizona contributed 309 (53.00% of the total ticks collected in Arizona) ticks, while California accounted for 182 (55.15% of California’s total). The following year saw a minor shift in these numbers, with Arizona’s contribution decreasing to 274 ticks (47.00% of Arizona’s total) and California’s decreasing to 148 ticks (44.85% of California’s total).
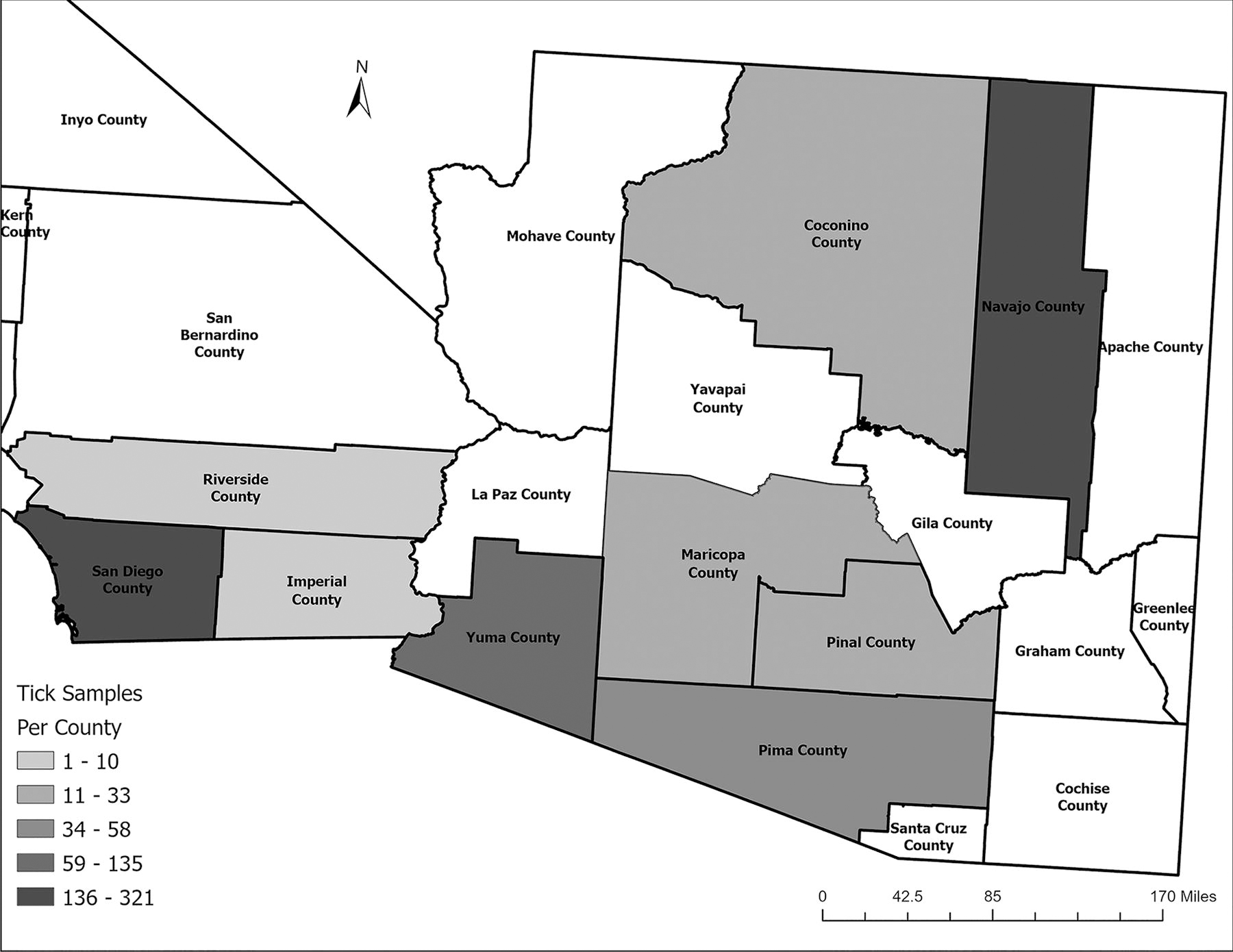
Map of counties in the southwestern U.S. participating in the 2021–2022 BiTeRS program, highlighting sampling sites and frequency of tick collections.
Ticks from Arizona included R. sanguineus (90.05%, 525 of 583) and Argas sp. (9.95%, 58 of 583), compared with California with D. similis (47.40%, 155 of 330), brown dog ticks (43.70%, 143 of 330), I. pacificus (7.88%, 26 of 330), O. megnini (0.61%, 2 of 330), and H. leporispalustris (0.30%, 1 of 330) (Table 1). Due to missing or damaged body parts, three ticks were not identified as species. Figure 2 is a barplot of the tick species found during each season.
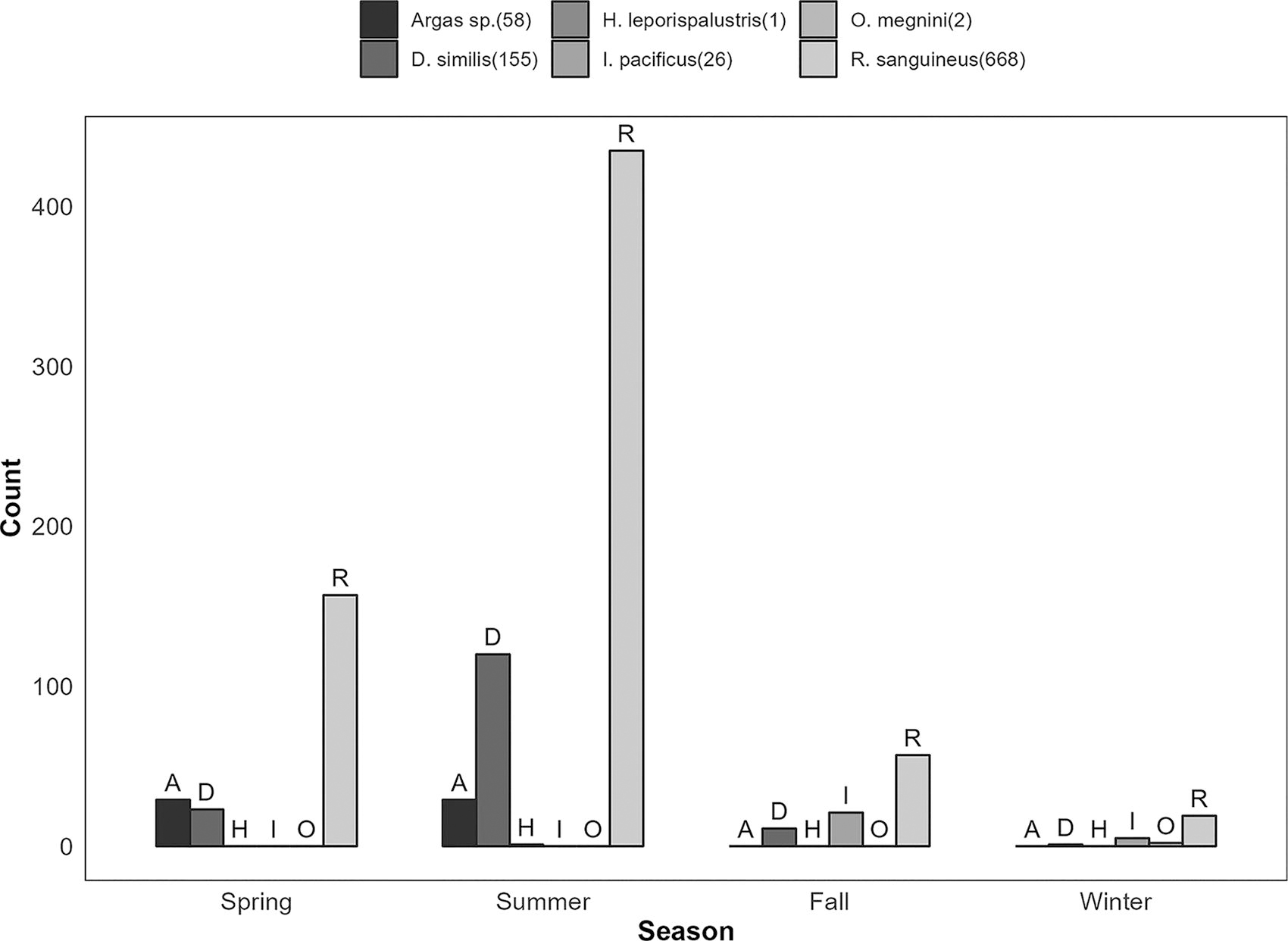
Counts of R. sanguineus, D. similis, Argas sp., H. leporispalustris, I. pacificus, and O. megnini ticks collected for the BiTeRS project in Arizona and California, 2021–2022. BiTeRS, border tick rickettsia surveillance; D. similis, Dermacentor similis; H. leporispalustris, Haemaphysalis leporispalustris; I. pacificus, Ixodes pacificus; O. megnini, Otobius megnini; R. sanguineus, Rickettsia sanguineus.
Descriptive Statistics for Rickettsia Spp. Infection in R. sanguineus, D. similis, and I. pacificus Ticks Collected in California and Arizona, 2021–2022
D. similis, Dermacentor similis; I. pacificus, Ixodes pacificus; R. sanguineus, Rickettsia sanguineus.
Forty (5.99%) of the R. sanguineus were PCR-positive for Rickettsia spp. (Table 1). The prevalence in 2021 was 8.66% compared with 4.09% in 2022. In Arizona, the prevalence was 5.71% similar to the 6.99% from California. More R. sanguineus (48.05%, 321 of 668) were collected from Navajo County than any other county, and these ticks had a 9.03% Rickettsia PCR-prevalence. R. sanguineus collections were seasonal, with none collected in winter and the highest number of ticks (65.12%, 435 of 668) with a PCR-prevalence of 6.67% in summer. Dogs were the host species for 98.35% (657 of 668) of R. sanguineus, with 5.94% PCR-positive. Ticks found feeding and attached constituted 98.80% (660 of 668) of the samples, with a Rickettsia spp. prevalence of 5.91%. Adult male ticks (48.65%, 325 of 668) and adult females (38.02%, 254 of 668) had a 4.92% and 8.66% prevalence, respectively, though the slightly elevated prevalence in females was not statistically significant ([odds ratio] OR: 1.84, 95% [confidence interval] CI: 0.95–3.64, p-value = 0.073).
Of the 155 adult D. similis collected, 18 (11.61%) were PCR-positive (Table 1). The majority (83.23%, 129 of 155) of these ticks were collected in 2021, with a Rickettsia spp. prevalence of 7.75%, compared with 16.77% (26 of 155) collected in 2022, with a higher prevalence of 30.77%. Almost the entire sample (99.35%, 154 of 155) was from San Diego County, with a PCR-prevalence of 11.69%. Most D. similis (77.42%, 120 of 155) were collected in summer. Seventy-one percent (70.97%, 110 of 155) were sourced from natural areas and had a pan-Rickettsia prevalence of 9.09% which was significantly lower compared to those ticks that were collected from dogs (OR: 0.31, 95% CI: 0.11–0.95, p-value = 0.034). Ticks collected from dogs accounted for 18.71% (29 of 155) of the samples with a 24.14% Rickettsia prevalence. About a quarter of the ticks, 24.52% (38 of 155), were attached and feeding, with a prevalence of 18.42% compared with 72.26% (112 of 155) not feeding, with a prevalence of 9.82%. These were not significantly different (OR: 0.48, 95% CI: 0.17–1.41, p-value = 0.165). Adult female ticks made up 87.10% (135 of 155) of the samples, with a prevalence of 10.37%, and had a lower pan-Rickettsia prevalence than adult males (prevalence of 20.00%), though this difference was not statistically significant (OR: 2.62, 95% CI: 0.67–8.63, p-value = 0.132). All D. similis that were submitted were adults.
A total of 26 western black-legged ticks were evaluated, 20 of which (76.92%) were positive for Rickettsia spp. Most (24/26, 92.31%) were collected in 2021, and only two (7.69%) in 2022. All were from San Diego County, California. Most (88.77%, 21 of 26) were collected during fall from a single individual dog host. Most I. pacificus (84.62%, 22 of 26) were adult females and the remaining four were adult males.
Among 58 Argas sp. ticks, all of which were collected from birds in Pima County, Arizona in spring and summer 2021, 24 (41.38%) were PCR-positive. All collected ticks were morphologically consistent larvae and were attached and fed on six avian host species. Comparison of a portion of the 16S rDNA gene to existing Genbank sequences showed the highest similarity to Argas giganteus (A. giganteus) also collected from raptors in Arizona (Genbank accession MK078102), but with only 86.6% identity and 100% coverage, the species could not be determined definitively. The majority of the Argas larvae were collected from western screech owls (27.59% of ticks collected, 16 of 58), Harris hawks (27.59%, 16 of 58), and mourning doves (17.24%, 10 of 48). PCR-prevalence was highest in the ticks collected from Cooper’s hawks (83.33%) and was 12.50% in ticks from the western screech owl and 31.25% in ticks from the Harris hawk.
We found only two spinose ears and one rabbit tick larvae, all from San Diego, California in 2021. Spinose ear ticks were found in winter from dogs, and rabbit ticks in summer from a natural area. All were PCR-negative for Rickettsia sp.
None of the 102 pan-Rickettsia PCR-positive samples yielded a positive result on R. rickettsii-specific PCR. Of the positive samples, DNA sequencing of the 17 kDa gene was attempted for 38 samples and successful for 30, while four were also sequenced successfully for the gltA gene. Phylogenetic trees can be seen for the 17 kDa gene in Figure 3 and gltA in Figure 4. The most commonly identified Rickettsia species in our collection was Rickettsia rhipicephali (R. rhipicephali): it was found in both R. sanguineus and D. similis from San Diego County (Table 2). Two additional D. similis from San Diego were found to be infected with a Rickettsial species that could not be differentiated from Rickettsia raoultii and Rickettsia montanensis via 17 kDa sequencing (Fig. 3). The Rickettsia species derived from I. pacificus matched closely with 17 kDa sequences from both R. monacensis (Rickettsia monacensis) and Rickettsia tamurae (R. tamurae) (99.02% similarity to both, multiple GenBank submissions). In Navajo County, R. massiliae was found in R. sanguineus. The gltA gene sequencing indicated that Argas sp. ticks from Pima County were infected with a rickettsial species most closely related to Candidatus Rickettsia tarasevichiae (R. tarasevichiae) (GenBank: MN450397, 97.79% identity) and had only 90% identity (70% coverage) to the Rickettsia hoogstraalii (R. hoogstraalii) previously identified in Argas spp. ticks collected from raptors in the same region (Genbank: MH717094).
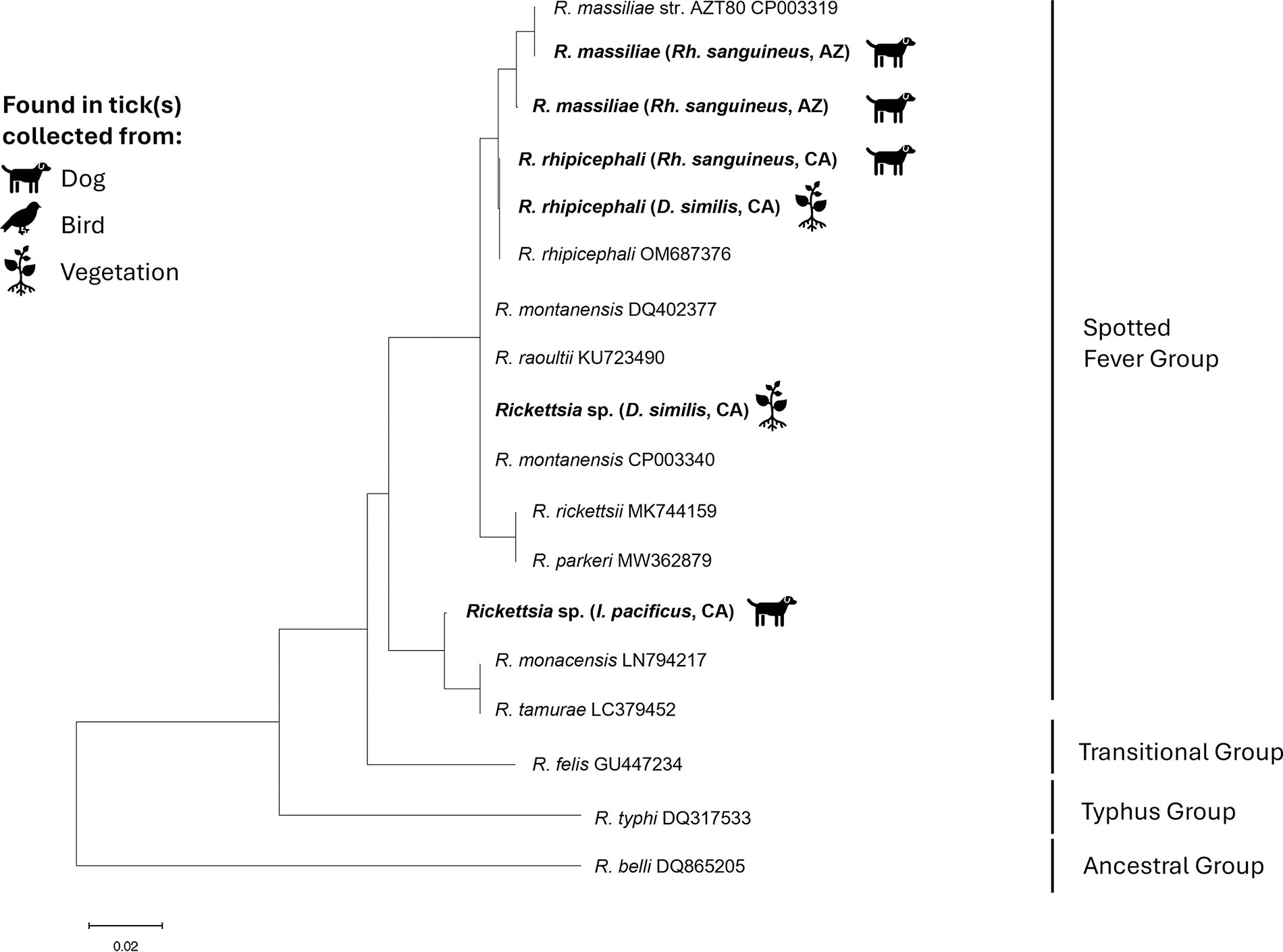
Phylogenetic tree based on comparison of 200 base pair portion of the 17 kDa gene of Rickettsia species showing relationship between samples collected as part of the BiTeRS program from 2021 to 2022 in Arizona and California (in bold) and reference samples collected from GenBank (included with accession numbers). Tree constructed using the maximum likelihood method and Tamura–Nei model in Mega 11. BiTeRS, border tick rickettsia surveillance.
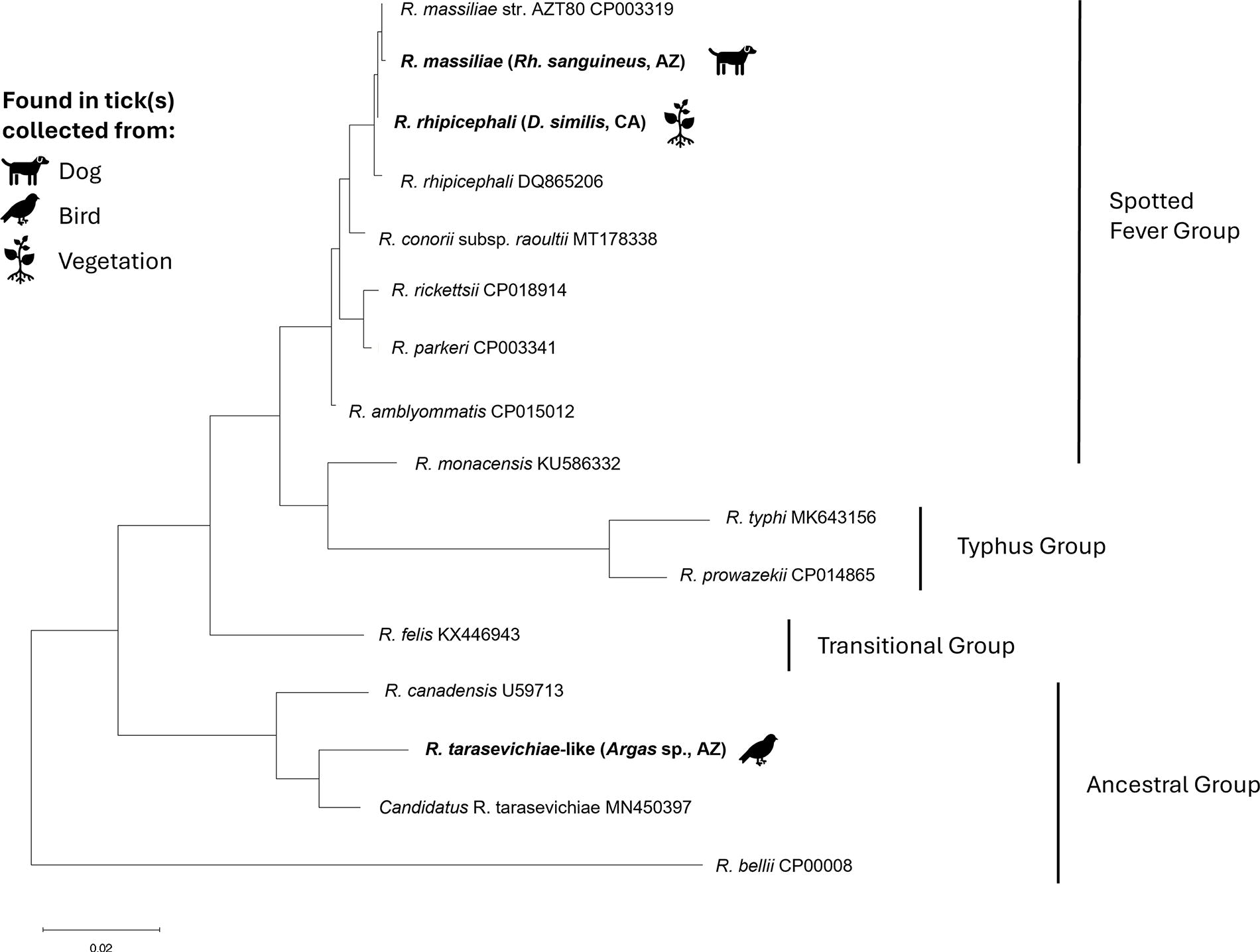
Phylogenetic tree based on comparison of an approximately 800 base pair portion of the gltA gene of Rickettsia species showing the relationship between samples collected as part of the BiTeRS program from 2021 to 2022 in Arizona and California (in bold) and reference samples collected from GenBank (included with accession numbers). Tree constructed using the maximum likelihood method and Tamura–Nei model in Mega 11. BiTeRS, border tick rickettsia surveillance program.
Results of BLAST Search of DNA Sequences from gltA or 17 kDa Genes of 31 pan-Rickettsia PCR-Positive Samples from 2021 to 2022 BiTeRS Project from Arizona and California
D. similis, Dermacentor similis; I. pacificus, Ixodes pacificus; R. monacensis, Rickettsia monacensis; R. raoultii, Rickettsia raoultii; R. sanguineus, Rickettsia sanguineus; R. tamurae, Rickettsia tamurae.
Discussion
Tickborne diseases pose an urgent public health concern, demanding rigorous tick surveillance to tackle this critical issue, particularly in the southwestern U.S. with its diverse tick population. In this study, through an opportunistic, partner-based approach, we identified six species of ticks carrying five species of Rickettsia, none of which were R. rickettsii, the species of greatest public health concern. Our study utilized a hybrid opportunistic and passive surveillance system, allowing us to tap into the benefits of working with partners and large-scale data collection via passive monitoring while incorporating active surveillance elements when partners had the motivation and resources to do so. Partners were offered resources to develop an active surveillance plan, the time, but differed in their ability to devote resources to this work. The approach, therefore, was to encourage tick collection as a corollary to activities in which the partner was already engaged, resulting in largely opportunistic sampling. Passive, opportunistic data gathering has shown significant success in large-scale tick and pathogen surveillance (Stromdahl et al., 2001; Barrett et al., 2015; Xu et al., 2016; Nieto et al., 2018) because it can help create large networks of stakeholders and provide more extensive geographic coverage with less requirement for data collection person-power. For example, New York state has a program of community-engaged tick surveillance which has fostered the development of an interactive mapping tool, tickMAP, to monitor tick emergence and tickborne pathogens (Hart et al., 2022). Programs like BiTeRS also serve to educate and remind stakeholders and the public about the risks of tick-borne diseases. However, passive, partner-engaged surveillance can be hampered by variables such as population density and the extent of community engagement and education (Cortinas and Spomer, 2014; Nelder et al., 2014) and relies heavily on the willingness and capacity of local communities to contribute. It also is difficult to manage the variability of the data. It would be very difficult with data such as ours to infer underlying tick density or other key attributes of the population’s risk. There were inherently self-selection biases for partners who decided to participate, and while data was not collected on partners’ decisions to participate, it is likely that entities with pre-existing concerns about ticks and tick-borne diseases were more likely to participate and submit ticks. While active surveillance arguably provides more precise estimates of pathogen prevalence (Nsubuga et al., 2011), its high cost and labor intensity often prevent large-scale application (Ogden et al., 2014; Centers for Disease Control and Prevention, 2008).
The reason R. rickettsii was not identified in any tick sample submissions is unlikely to represent total absence in the region and is very likely a product of where samples were submitted from and very low prevalence. Even in RMSF epidemic regions, the prevalence of R. sanguineus is typically less than 1% (Eremeeva et al., 2011)(Foley et al., 2019; Foley et al., 2024b) and while R. rickettsii may occur in Dermacentor ticks in California, it is found infrequently (Osborne et al., 2020). It may also represent an additional form of bias, where due to social or economic factors, detection and reporting of ticks is less likely to occur in the circumstances most likely to drive R. rickettsii transmission (Nicholson et al., 2006), and presents an additional limitation to the surveillance methods used here.
Even in the absence of R. rickettsii detection, however, several other important findings arose from this work. The study supports that there is no specific “tick season” in southern California, with ticks collected and submitted in every season. We received R. sanguineus and D. similis primarily during the fall and summer, and I. pacificus mainly in winter and fall. This pattern aligns with known seasonality for these ticks in the state. Dermacentor species are known to tolerate some aridity and elevated temperatures, remaining active into summer months in California, in contrast with I. pacificus which requires a tight, relatively cool, and high humidity environmental window (MacDonald, 2018). However, an important caveat to these inferences is that systematic prospective sampling was not done, limiting possible positive data points during seasons when partners were not undertaking surveillance. Moreover, there are too few data on juvenile ticks to allow for analysis of the seasonality of the different life stages. We also found the single H. leporispalustris, which rarely bites humans but is a competent vector for R. rickettsii (Freitas et al., 2009) and has been implicated in human RMSF in Costa Rica (Hun et al., 2008). O. megnini feeds primarily on livestock with many records of it feeding on other hosts as well (Keirans and Pound, 2003; Nava et al., 2009). The agents of Q-fever and RMSF have been associated with O. megnini in older literature (Bustamante et al., 1946; Jellison et al., 1948).
Of concern is the finding of R. sanguineus in the summer and fall, given suggestions that this species may demonstrate increased aggression towards humans at elevated temperatures (Parola et al., 2008; Backus et al., 2021). The data also show the widespread presence of R. sanguineus across multiple counties within the southwestern U.S., corroborating previous findings that have indicated a significant prevalence of this tick species along the U.S.–Mexico border, particularly in arid regions of the western border (Eremeeva and Dasch, 2015). However, none of the 945 R. sanguineus collected during this study tested positive for R. rickettsii, in contrast with other studies in northern Mexico (López-Pérez et al., 2021; Álvarez-Hernández et al., 2017; Zazueta et al., 2021). This may be due to the reduced presence of roaming dogs in the southwestern U.S. (Rowan and Kartal, 2018) compared to parts of northern Mexico (Tinoco-Gracia et al., 2009), which can amplify R. rickettsii and R. sanguineus (Demma et al., 2005; Nicholson et al., 2006; Drexler et al., 2014). It is possible that sampling closer to the U.S.–Mexico border could have resulted in a higher R. rickettsii infection prevalence in our samples (Eremeeva et al., 2011; Bustamente Moreno and Pon Méndez, 2010). This highlights the need for further research with larger sample sizes and uniform sampling methods across different geographical regions.
All of the Argas sp. ticks in this study came from a facility in Tucson that cares for wildlife, and elevated numbers of these ticks have been reported previously (Latas et al., 2020). However, both the tick species A. giganteus identified in that study differ markedly from what we identified here. Our research broadens our understanding of the relationship between bird species and the soft tick genus Argas sp. in Arizona, which has also been documented in California, New Mexico, Utah, and Sonora, Mexico (Cooley et al., 1944; Furman and Loomis, 1984). The ticks are often hosted by a number of neotropical bird species that migrate through or nest in the Sonoran Desert (eBird, 2024; Rappole et al., 2011). Some of the estimated 19 million neotropical ticks annually introduced to the U.S. via bird migration may contribute to expanding tick species’ geographical ranges, potentially exacerbated by climate change (Cohen et al., 2015; Sonenshine, 2018). Although our study identified ticks, particularly Cooper’s hawk, Harris’s hawk, and red-tailed hawk, the challenges of comprehensive tick sampling from migratory birds may mean that the actual diversity and distribution of Argas ticks in the area could be underrepresented. We also likely have only a biased sense of the tick’s geographical distribution given our limited sampling space.
The study found five species of rickettsiae in various tick species. Infections from SFG R. massiliae are milder compared to RMSF and typically not life-threatening (Biggs et al., 2016). R. massiliae is reported to be a pathogen of both humans and dogs (Beeler et al., 2011; Eremeeva et al., 2006). It is often reported from the Mediterranean (Beati and Raoult, 1993), and has been reported before in R. sanguineus from California (Beeler et al., 2011), Virginia (Fornadel et al., 2013), and Eastern Arizona (Eremeeva et al., 2006). In this dataset, only R. sanguineus from Arizona tested positive for R. massiliae. R. rhipicephali is not considered pathogenic but is closely related to R. massiliae, nesting within the same clade (Fig. 3) and was found in both Dermacentor and Rhipicephalus ticks and in both California and Arizona in this study.
A rickettsial species in I. pacificus in San Diego County most closely matched zoonotic R. tamurae and R. monacensis (Jado et al., 2007; Imaoka et al., 2011; Kim et al., 2017). R. monacensis has a broad range of vectors including Ixodes ricinus (I. ricinus) in Germany (Simser et al., 2002), Ixodes boliviensis in Costa Rica (Springer et al., 2018), and I. pacificus in northern California (Osborne et al., 2020). The rickettsiae in the Argus ticks are closely related to Candidatus R. tarasevichiae, which has been primarily detected in Ixodes persulcatus ticks from China and Russia (Shpynov et al., 2003; Li et al., 2016) and thought to be an emerging human pathogen. It falls within the Ancestral group in a clade with Rickettsia canadensis (Fig. 4). The species identified here differs markedly in the gltA sequence from the R. hoogstraalii found in A. giganteus on raptors in the region previously (Latas et al., 2020), and R. hoogstraalii has only 90% identity with Candidatus R. tarasevichiae, indicating a separate species found here. A closely related organism was found in I. pacificus ticks from a gray fox in Ensenada, Baja California (López-Pérez et al., 2021). Definitively identifying the Rickettsia species would require culture and further molecular assays.
It is clear that the full scope of tick-host associations in the American Southwest has not yet been revealed. Pressing concerns are tick and infection seasonality and that poorly studied Argas sp. ticks and their rickettsiae may be spread by neotropical birds. Regular monitoring of R. sanguineus is crucial. Further study of rickettsial and tick diversity could illuminate roles in disease dynamics and opportunities for management. Hybrid active, passive, sentinel, and community-based programs will enable us to detect early signs of disease spread, mitigate potential impacts, and protect public health more effectively.
Footnotes
Acknowledgment
The authors sincere gratitude extends to Pranav Pandit for assistance in statistical analysis, Christopher Barker for help with ArcGIS Pro, Paulo Salazar for assistance with MEGA 11, and the partner organizations that form the backbone of the BiTeRS program. The authors also extend their thanks to Mia Reed, Anaisia Brown, and Olivia Smith for their invaluable help with laboratory analysis, RT-PCR, and DNA extraction.
Author Disclosure Statement
Authors declare that they have no financial or personal interests to disclose.
Funding Information
This research was funded by the Pacific Southwest Regional Center of Excellence for Vector-Borne Diseases, which is funded by the U.S. Centers for Disease Control and Prevention (Cooperative Agreement 1U01CK000516).