
Abstract
The current DNA vaccine formulations are not optimal for stimulation of CD8+ T cells, which are required for clearing virally-infected cells. Here we show that CD8+ T cell-stimulating activity can be effectively augmented by combining DNA vaccination with protein transfer. C57BL/6 mice were injected intramuscularly with an anti-SARS-CoV DNA vaccine admixed with a lipid-derived conjugate of 4-1BBL, a potential CD8+ T-cell co-stimulator. The inclusion of the lipidated co-stimulator greatly enhanced cellular immune responses, especially the CTL response, induced by the DNA vaccine. The adjuvant effect of 4-1BBL was lipidation-dependent, indicating that it functions as a cell membrane–anchored co-stimulator. Results of our study suggest, for the first time, that muscle cells may be modified in situ, at the DNA injection site, into APC-like cells to allow direct priming of CD8+ T cells and thereby improve the efficacy of DNA vaccines.
Introduction
Despite the fact that DNA vaccines can stimulate CTL responses, these responses have been consistently inferior to those induced by live attenuated vaccines, both quantitatively and qualitatively. Unlike a live virus that can infect immune cells, such as dendritic cells (DC), that are specially equipped to activate T cells, a DNA vaccine administered into the body (mostly by intramuscular injection) enters mainly non-immunogenic cells, such as muscle cells. These cells, although capable of translating the DNA sequence into a protein antigen, inherently lack the ability to express adjuvant protein molecules (called co-stimulators) necessary for sensitizing T cells, and are therefore unable to directly activate T cells. Instead these antigen-producing cells must release the antigen to DC, which in turn present it to T cells, along with the expression of necessary co-stimulators. The requirement for antigen transfer, or so-called “cross-priming,” is a rate-limiting event that fundamentally limits the efficacy of DNA vaccines (16).
The field of DNA vaccine research is currently focused on improving the efficiency of cross-priming and/or the means to deliver DNA vaccine directly to DC (8,24). Here we are exploring a cell modification method known as protein transfer to enhance the immune response of DNA vaccine. In this method, protein A (PA, a commercially available, soluble staphylococcal protein), after being chemically derivatized with palmitate, is combined with a T-cell co-stimulator fused with the Fc domain of human IgGγ1. The resulting conjugate can be directly incorporated into the cell membrane. We previously showed that co-stimulators displayed in this manner on the cell surface retain their native capacity to co-stimulate T cells (14,27,28).
In the present study we used protein transfer to convert non-immunogenic cells in situ into immunogenic antigen-presenting cells capable of directly activating CD8+ T cells. Our study demonstrates for the first time that in-situ protein transfer can improve the efficacy of DNA vaccines.
Materials and Methods
Mice
C57BL/6 and BALB/c mice (6–8 weeks of age) were purchased from the Jackson Laboratory (Bar Harbor, ME) and used in accordance with institutional guidelines for animal care.
Vaccines and antigens
DNA plasmid encoding the N protein of SARS-CoV (pcD3d/N) was constructed as previously described (9), and used as a vaccine. The recombinant plasmid pET28a-N was constructed by subcloning the N gene in the prokaryotic expression plasmid and transformed into E. coli DH5a for expression of the N protein. Then the N protein of SARS-CoV was identified with SDS and Western blot using pooled sera from patients in the convalescence phase of SARS. The expressed N protein was purified using Ni-NTA Fast Start (Qiagen Inc., Valencia, CA) according to the manufacturer's directions. The purified N protein was further treated with Detoxi-Gel Endotoxin Removing Gel (Pierce, Rockford, IL) and tested by the Limulus amebocyte lysate assay (Sigma-Aldrich, St. Louis, MO), to ensure that it was free of endotoxin contamination (below the detection level of 0.1 EU/mL). The production of murine 4-1BBL Fc fusion protein (4-1BBL-Fc), and palmitated-protein A (PPA) used for Fc protein transfer have been described previously (27).
Immunization
The conjugates of PPA and 4-1BBL-Fc fusion or human IgG1 (14) proteins were first generated by combining the components at a 1:1 ratio (wt/wt) in DMEM and incubated on ice for 30 min before injection as described previously (1). The conjugates of PPA (2 μg PPA/mouse) and 4-1BBL-Fc fusion (2 μg PPA/mouse) or hIgG (2 μg PPA/mouse) proteins were mixed with pcD3d/N (10, 30, or 100 μg/mouse) and injected intramuscularly (in 50 μL of DMEM) into mice (26). The animals were boosted with the same formulation on day 14 after the first immunization. Mice were injected with saline, pcD3d/N alone, pcD3d/N plus PPA, pcD3d/N plus 4-1BBL, pcD3d/N plus protein A and 4-1BBL, or pcD3d/N plus PPA and hIgG as controls. Immunized animals were bled on day 21 after the second immunization and serum were collected.
Antigen-specific DTH
On day 14 after the second immunization, all mice were challenged with the N protein of SARS-CoV in the right footpad (10 μg N protein in 20 μL saline/mouse). The same animals were also contralaterally injected with saline solution (20 μL saline/mouse) in the left footpad as control. The thickness of the footpad was measured 24, 48, and 72 h later with a digital micrometer. The thickness of the right footpad minus that of the left footpad was used to quantify delayed-type hypersensitivity (DTH).
T-cell proliferation
Splenocytes were prepared on day 14 after the second immunization and CD4+ T cells were purified as previously described (29). CD4+ T cells (0.5–1 × 105) were activated by NP protein-pulsed (5 μg/mL) and mitomycin C-inactivated splenic accessory cells (2.5–5 × 104). The cells were plated in triplicate into the wells of a U-bottom 96-well plate. Proliferation was quantitated by 3H-thymidine incorporation.
CTL assay in vitro
Splenocytes of immunized C57BL/6 mice were collected on day 14 after the second immunization, stimulated with N protein (5 μg/mL) for 3 d, and used as effectors. Splenocytes from naïve mice were depleted of T cells and pulsed with the N protein (10 μg/mL) overnight, labeled with CSFE, and used as specific targets. Killing was initiated by mixing the effectors with the targets at different ratios. After 12 h of incubation, the cell mixture was analyzed by flow cytometry as previously described (29). Lysed target cells were identified as the CFSE+/7-AAD+ population. Raw data were converted into specific lysis using the formula: specific lysis = (x − c)/(100 − c), where x is the percentage of 7-AAD+ target cells in the presence of effectors, and c is that in the absence of effectors (spontaneous lysis).
Antibody measurement
Serum anti-N antibody levels were assayed by ELISA as previously described (9).
Statistical analysis
Data were analyzed for statistical significance using Student's t-test. Difference was deemed statistically significant if p < 0.05.
Results
4-1BBL protein transfer enhances DNA immunization-induced anti-N DTH reactions
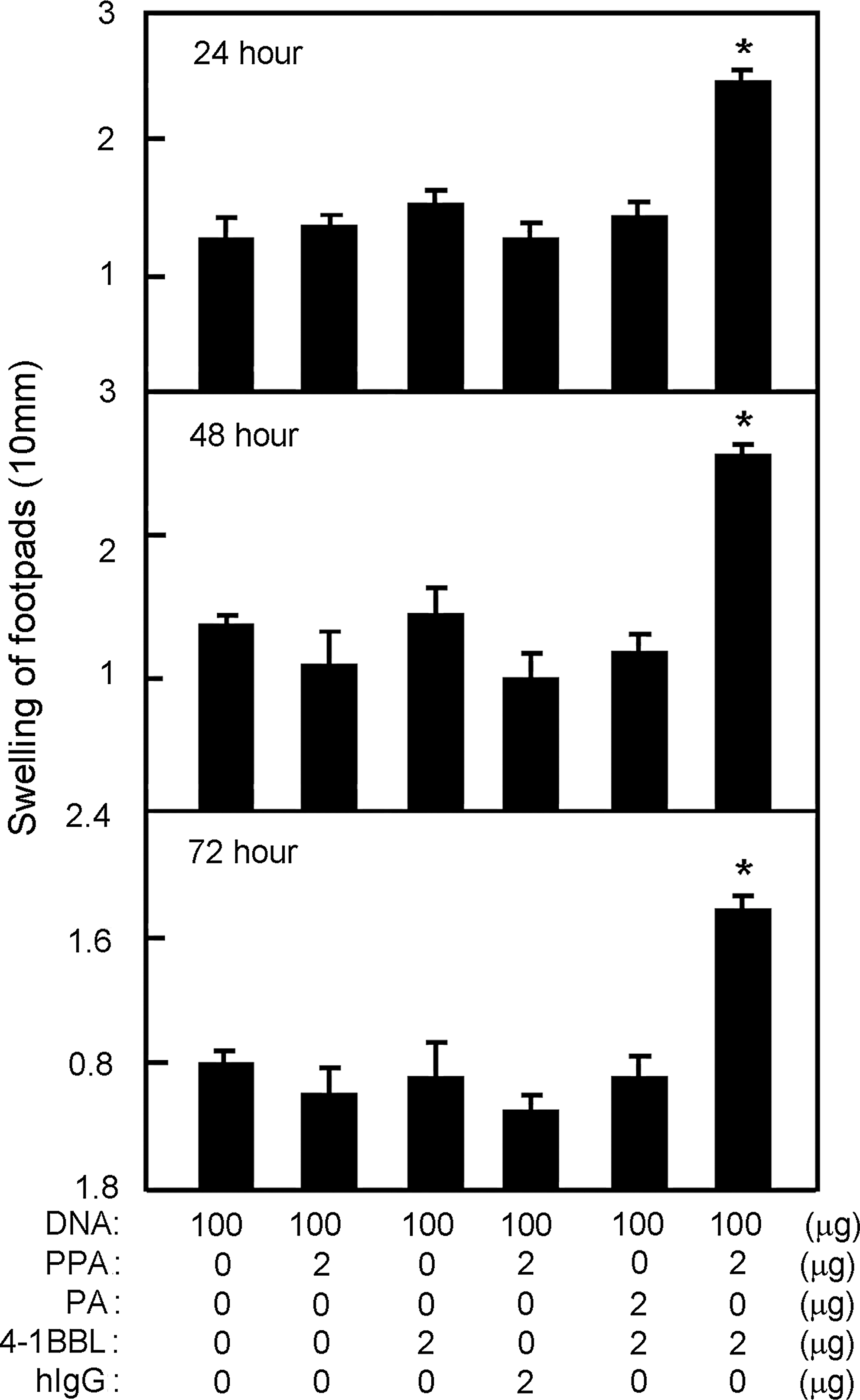
To first determine whether 4-1BBL protein transfer can enhance the immune response to DNA vaccination, we titrated the dose of PPA-4-1BBL conjugates needed for effective induction of DTH. We immunized C57BL/6 mice with a constant amount of pcD3d/N together with graded amounts of PPA-4-1BBL conjugates on days 0 and 14. Seven days after the second immunization, the mice were rechallenged with the N protein at a footpad. The swelling of the footpad was recorded at 24, 48, and 72 h. The strongest DTH reaction was from mice immunized with 100 μg of pcD3d/N and 2 μg 4-1BBL conjugated to 2 μg of PPA (Fig. 1A). Moreover, the adjuvant effect of 4-1BBL was lipid-dependent, because 4-1BBL alone or PA-4-1BBL showed no adjuvant effect. This confirms that only membrane-anchored 4-1BBL can serve as an effective adjuvant (Fig. 1B).
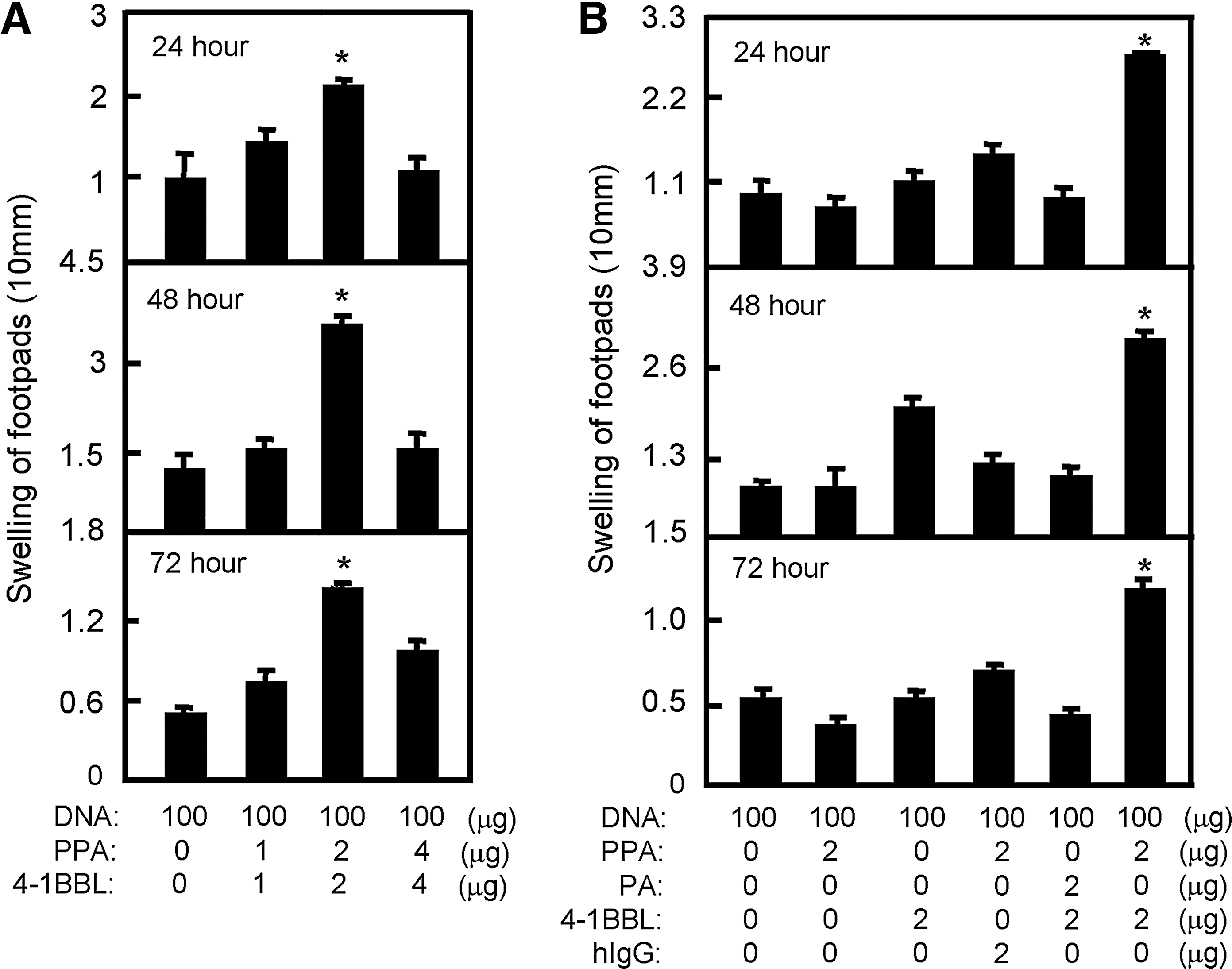
4-1BBL protein transfer enhances DNA immunization-induced anti-N DTH reactions. Immunized mice were rechallenged at a footpad with the N protein. (
4-1BBL protein transfer enhances antigen-specific T-cell proliferation
DTH responses are T-cell-dependent. To determine whether 4-1BBL enhances the induction of T-cell responses, splenocytes were isolated from mice 7 days after the boost injection, and CD4+ T-cell proliferation in response to ex-vivo antigen re-stimulation was assessed by [3H]-thymidine incorporation. The results showed that the strongest T-cell proliferation was induced in mice immunized in the presence of PPA-4-1BBL, whereas 4-1BBL without PPA was only moderately effective (Fig. 2A), confirming that 4-1BBL protein transfer enhances antigen-specific T-cell proliferation.
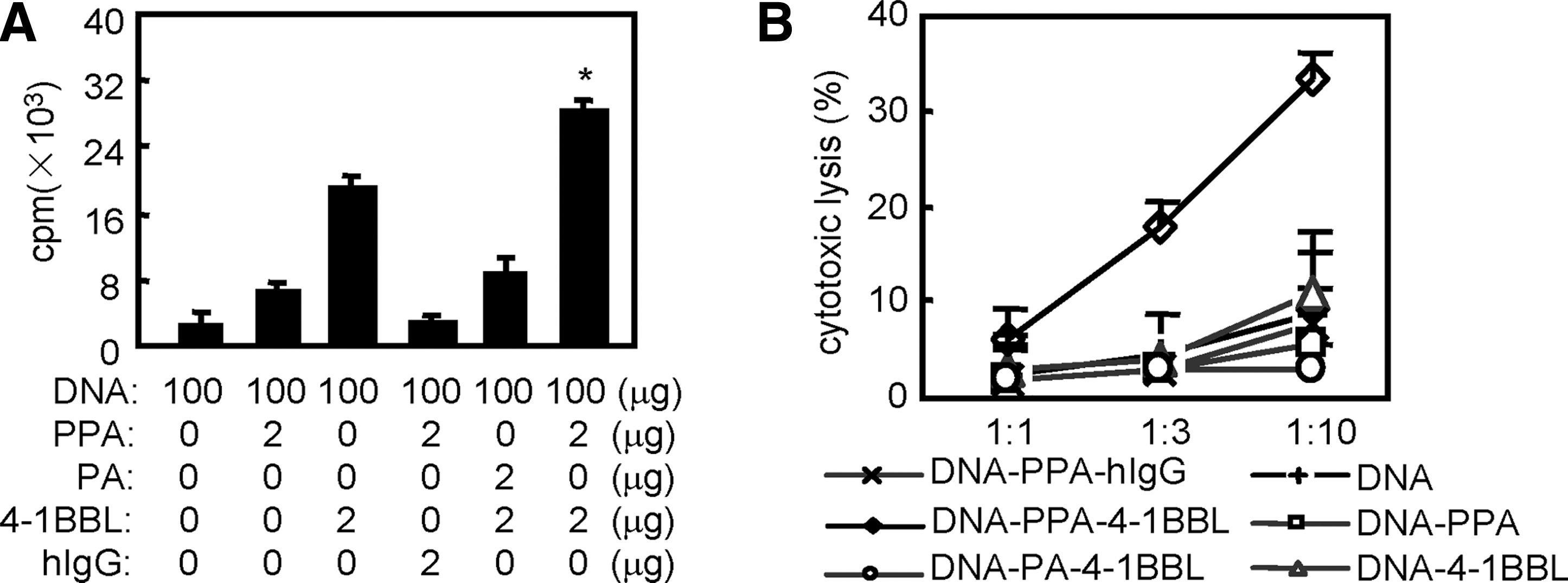
4-1BBL protein transfer enhances antigen-specific T-cell immune responses. (
4-1BBL protein transfer enhances antigen-specific CTL activity
The antigen-specific CTL response plays a key role in the protection against or eradication of viruses or other intracellular pathogens. To examine whether 4-1BBL protein transfer enhances CTL activity, we performed an in-vitro CTL assay on day 7 after the second immunization. The results showed that DNA vaccination in the presence of the PPA-4-1BBL conjugates induced much stronger CTL activity than in the absence of the conjugates (Fig. 2B), consistent with the idea that CD8+ T cells might be activated at the injection site by 4-1BBL-modified muscle cells.
4-1BBL protein transfer enhances the humoral response
To determine the adjuvant effect of the PPA-4-1BBL conjugates on the humoral response stimulated by DNA vaccination, we collected serum from immunized mice 14 days after the second immunization and analyzed serum IgG levels by ELISA. The results showed that the serum from mice treated with pcD3d/N and the PPA-4-1BBL conjugates contained significantly higher anti-N antibody titer than pcD3d/N or other controls (Fig. 3), suggesting that 4-1BBL protein transfer also enhances the humoral response.
4-1BBL protein transfer enhances the humoral response to N protein. C57BL/6 mice were immunized with the indicated combinations and anti-sera were tested for N-specific IgG titer by ELISA. Shown is one of three independent experiments using three mice per group (bar indicates the mean and SD from triplicate wells; *p < 0.05 compared with the other groups).
4-1BBL protein transfer enhances DTH reactions in BALB/c mice
To exclude the possibility that the adjuvant effect of the PPA-4-1BBL conjugates was due to the particular genetic background of the C57BL/6 strain, we tested the effect of the conjugates by immunizing a different strain, BALB/c, with the same pcD3d/N (100 μg/mouse) and the PPA-4-1BBL combination. On day 7 after the second immunization, the DTH reaction was tested as described earlier. The strongest DTH reaction was induced in mice immunized with pcD3d/N and the PPA-4-1BBL conjugates (Fig. 4), as noted before in the C57BL/6 mice, indicating the general applicability of the PPA-4-1BBL conjugates as an adjuvant.
4-1BBL protein transfer enhances DTH reactions in BALB mice. BALB/c mice were immunized with the indicated combinations and DTH reactions were measured (bars indicate mean and SD from 2–4 independent experiments, each using at least three mice per group; *p < 0.05 between the right footpads and the left footpads of the same group).
Discussion
DNA vaccination can induce both humoral and cellular immune responses, which lead to protection against various types of infection, including SARS (26). Several DNA vaccines encoding different genes of SARS-CoV can elicit different levels of humoral and cellular immune responses (6,7,30). Some researchers report that N protein is a highly immunogenic protein of the SARS-CoV (2). Our previous study demonstrated that DNA vaccine encoding N protein of the SARS-CoV induces high level humoral and CTL responses in comparison with other proteins (M and E) (9). So in this study, a plasmid pcD3d/N, encoding N protein of the SARS-CoV, was used as the DNA vaccine.
To date, the strength of the immune response induced by DNA vaccines has been relatively weak compared with conventional vaccines. Unlike a live virus that can infect immune cells, a DNA vaccine administered into the body enters mainly non-immunogenic cells, such as muscle cells. These cells lack the ability to express co-stimulators that are necessary for sensitizing T cells. Co-stimulators play a key role in the induction of the T-cell response (10). Effective T-cell activation by APC requires the first signal from antigens presented by the MHC molecules, and the second signal from co-stimulators. Although a significant level of DNA is expressed in muscle cells following intramuscular inoculation of plasmid DNA, and the resulting antigen can be presented by the MHC I molecules expressed by the cells, these cells do not normally express co-stimulators (4,5). Previously we developed a protein transfer–based strategy for co-expressing co-stimulators, including B7.1, 4-1BBL, and TRANCE on the surface of cells to generate anti-tumor immune responses (27 –29). We showed that multiple co-stimulator-Fc fusion proteins, after being pre-conjugated with PPA ex vivo, can be directly injected into a tumor and generate anti-tumor immune responses (29). Thus co-stimulator protein transfer offers a useful means to provide the necessary co-stimulatory signals for vaccination. Here we sought to use protein transfer to modify muscle cells with a co-stimulator, 4-1BBL, in order to enable direct priming of CD8+ T cells by the cells.
4-1BB is a co-stimulator known to promote proliferation and survival of T cells, especially CD8+ T cells (3,12). When provided in conjunction with a strong antigenic signal through the TCR, 4-1BBL can allow the expansion, cytokine production, and survival of T cells (3,20). Consistent with these previous observations, the present study shows that the PPA-4-1BBL conjugates induced strong immune responses, especially the proliferative and CTL responses, against a major SARS-derived antigen. Whether PPA-4-1BBL actually converted muscle cells at the vaccination site into APC-like cells requires further investigation.
With the simplicity in delivering defined amounts of immunostimulatory protein in combination with a DNA vaccine, protein transfer may offer a useful means to enhance the efficacy of DNA vaccination. In support of this notion, the present study shows that, compared to animals immunized with the pcD3d/N DNA alone, the inclusion of the PPA-4-1BBL conjugates significantly enhanced the cellular and humoral responses to the vaccination. Importantly, the adjuvant effect of the conjugates is both PPA- and 4-1BBL-Fc-dependent, suggesting that the mode of action involves cell membrane anchoring. Thus this study suggests, for the first time, that protein transfer can be combined with DNA vaccination more efficiently to stimulate immune responses.