
Abstract
HIV/HCV coinfected patients tend to develop hepatitis C (HCV)-associated liver disorders. Because the chemokine receptor CXCR3 participates in lymphocyte trafficking during hepatic inflammation, it may participate in the escalated liver disorders of coinfected patients. However, to date, the relative frequency and density of receptor on lymphocytes has not been established. This study compared the CXCR3+ phenotype under various in vitro conditions between lymphocytes from healthy and coinfected individuals. Peripheral lymphocytes were stimulated with phytohemagluttinin for 0–7 d and phenotypes were determined by flow cytometry. Secreted cytokines were measured in culture supernatants by ELISA. Phenotypic differences were observed between groups. CD4+CXCR3+ frequency between groups was equivalent before and during early activation, but increased only among non-infected individuals during late activation (p<0.001). In contrast, CD8+CXCR3+ frequency was consistently greater (p<0.05) among HIV/HCV patients throughout activation. Among those who were non-infected, CD8+CXCR3+ frequency increased (p=0.002) during late activation. However, CD8+CXCR3+ frequency among HIV/HCV patients increased within 24 h of activation (p=0.008), and was nearly universal by late activation (p<0.001). Both groups elaborated Th-1 cytokine profiles; however, coinfected patients released more inflammatory cytokines (p<0.01) than non-infected individuals. In summary, we demonstrated that CD8+ lymphocytes from HIV/HCV-infected patients expressed more CXCR3 and showed greater upregulatory ability upon activation. The atypical CXCR3 expression and enhanced Th-1 cytokine elaboration among coinfected patients could potentially stimulate increased lymphocyte migration during hepatic inflammation.
Introduction
HCV induces inflammatory changes within the liver characterized by extensive infiltration of T lymphocytes. The extent of hepatic damage appears to be proportional to the number of infiltrating CD8+ lymphocytes (9,11,12,17). While the process of lymphocyte recruitment to the liver is complex, it begins with systemic release of chemokines from the liver. Chemokines then bind to unique receptors on specific inflammatory cells, which facilitates their migration to the liver. Numerous lines of evidence indicate that the chemokine interferon γ-inducible protein-10 (IP10), and its receptor CXCR3 are particularly relevant to lymphocyte trafficking in HCV disease (3,7,9,11,15,21,23,27,31). Intrahepatic mRNA levels of IP10 and CXCR3 are increased with HCV. Elevations in circulating IP10 levels indicate the degree of liver injury among both HCV- and HIV/HCV-infected patients. And histologically, CXCR3 is expressed by virtually all infiltrating lymphocytes during hepatic inflammation (1,3,4,6,9,10,11,12,17 –21,23,24,26,28 –30).
The magnitude of lymphocyte recruitment to the liver and the subsequent extent of liver damage during HCV inflammation are regulated, in part, by the number of receptor-positive lymphocytes (6,7,11,12,15,21,23). However, few data are available regarding the frequency or expression of CXCR3 on lymphocytes from either healthy or virally-infected individuals. This study compared in vitro expression of CXCR3 on peripheral lymphocytes from HIV/HCV-coinfected and non-infected healthy individuals before and after activation. Our results showed that coinfected patients have a (1) higher frequency and (2) more robust expansion of CD8+CXCR3+ lymphocytes than non-infected individuals. This atypical phenotype may contribute to the escalated hepatic inflammation experienced by coinfected patients.
Materials and Methods
Patients
The study included patients with HIV/HCV coinfection prospectively evaluated for chronic HCV. All patients were clinically stable and receiving HAART regimens. Patients were age >18 y and positive for anti-HIV antibody and HCV RNA using commercial assay. Quantitative HCV RNA testing was performed (Amplicor; Roche Molecular Systems, Branchberg, NJ), and HCV genotyped by Inno-Lipa (InnoGenetics, Tarrytown, NY). Patients were excluded if they had a prothrombin time >2 sec than controls, the presence of ascites, thrombocytopenia, active or recent (within 3 months) opportunistic infection related to HIV, advanced HIV disease with life expectancy less than 1 y, serum creatinine >2.5 mg/dL, were HBV surface antigen-positive, or had any other form of chronic liver disease. Blood was also collected from healthy volunteers (controls). All practices conformed to IRB guidelines for the protection of human subjects.
Cell cultivation
Blood was collected from 40 coinfected and 20 age- and gender-matched non-infected individuals (control). Peripheral mononuclear cells (PMNCs) were isolated under sterile conditions by sedimentation in Histopaque for 10 min at 3000 rpm. The cells were washed and red blood cells lysed with Tris-NH4Cl. The cells were resuspended in RPMI with 10% fetal bovine serum (complete media) and cryopreserved in 10% dimethyl sulfoxide under liquid nitrogen. Prior to assay, cells were thawed, resuspended in complete media, and incubated for 24 h at 37°C to allow cell and surface protein recovery. Initial experiments showed that allowing 24-h recovery yielded results equivalent to freshly-drawn cells. The cells were stimulated with phytohemaglutinnin (PHA; Sigma-Aldrich, St. Louis, MO) for up to 7 d. Briefly, the cells (2×106/mL) were mixed with an equal volume of 10 μg/mL PHA in complete media and incubated in Costar plates. Unstimulated cells were cultivated without PHA. In certain experiments, cells were treated with 100, 200, 500, and1000 ng/mL IP10 (Sigma-Aldrich) for up to 24 h.
PHA-induced proliferation was evaluated by plating cells into microculture wells and cultivating for 7 d (8). Proliferation was measured by tritiated thymidine incorporation, and data expressed as counts per minute (cpm). End-of-assay viability was assessed by trypan blue exclusion assay.
Flow cytometry
CXCR3+ lymphocytes were identified using 3-color immunofluorescence. Cells (4×106 cells/mL) were washed and resupended in PBS with 0.5% bovine serum albumin, 25 μL transferred to new tubes and non-specific binding was blocked by treatment with l μg IgG for 15 min at room temperature. Pelleted cells were incubated with 10 μL each of the following antibodies (CD4−PerCP, CD8-PE, and CXCR3-FITC; R&D Systems, Minneapolis, MN) for 45 min at 4°C. The cells were washed, resuspended in PBS, and enumerated on a BD FacScan device. Isotype-matched controls were used to monitor background fluorescence and set quadrant gates. Lymphocytes were initially gated using forward- and side-scatter parameters. Then, lymphocytes were further gated based upon CD4+ or CD8+ expression and dot plots created based upon CXCR3+. Ten thousand events of CD4+ and CD8+ cells were collected. Signal strength was quantitated by comparing mean fluorescence intensity to a standard curve of calibrated beads (Quantum Beads; Bangs Lab, Fisher, IN), and data converted to molecules of equivalent soluble fluorochrome (MESF). MESF values indirectly indicate receptor density on the cell surface (8). All experiments were performed a minimum of 5 times.
ELISA
Cells were cultured with and without PHA for 24 h. Supernatants were collected and assayed in duplicate for IL-2, IFN-γ, TNF-α, and IL-10 by commercial ELISA following the manufacturer's instructions (R&D Systems).
EliSpot
The frequency of IFN-γ-producing cells was determined using EliSpot assays (R&D Systems). Briefly, the cells (5×105/mL) were cultured in triplicate and stimulated with PHA for 24 h at 37°C in antibody-coated microtiter plates. The number of IFN-γ-producing cells was visualized following the manufacturer's protocol and spots were counted manually with a dissecting scope.
Statistical analysis
Results were expressed as means±standard deviation. Group parameters were compared using two-way non-parametric t-tests with 95% confidence intervals using Graphpad Prism software (Graphpad Software, San Diego, CA). Significance was defined as p≤0.05.
Results
Patients
Patient demographics are shown in Table 1. Patients were primarily HCV genotype 1 with evidence of liver dysfunction and low CD4 percentages. HIV RNA was non-detectable in the majority of patients.
HCV, hepatitis C virus.
Phenotypic expression of CXCR3
CXCR3 expression on resting lymphocytes was first determined (Table 2). A low frequency of CD4+CXCR3+ cells was seen among controls and co-infected patients. Frequency was unaffected by exogenous IP10. The frequency of CXCR3+ cells was greater on CD8 than CD4 cells among control (p<0.001) and HIV/HCV patients (p<0.001). However, the frequency of CD8+CXCR3+ cells was greater (p=0.03) among coinfected patients than controls. Inclusion of exogenous IP10 did not affect receptor expression.
Frequency of receptor-positive cells was greater on CD8 than CD4 cells; p<0.001.
Peripheral mononuclear cells (PMNCs) were collected from non-infected healthy volunteers (control) and HIV/HCV co-infected patients and cryopreserved in liquid nitrogen prior to the study. Thawed cells were incubated 24 h in complete media at 37°C to allow cell recovery and stained as described in the text. The percent of unstimulated CD4+CXCR3+ and CD8+CXCR3+ cells were analyzed using a BD FACScan device. Data are also shown for cells treated with 100 ng/mL IP10 for 4 h.
HCV, hepatitis C virus.
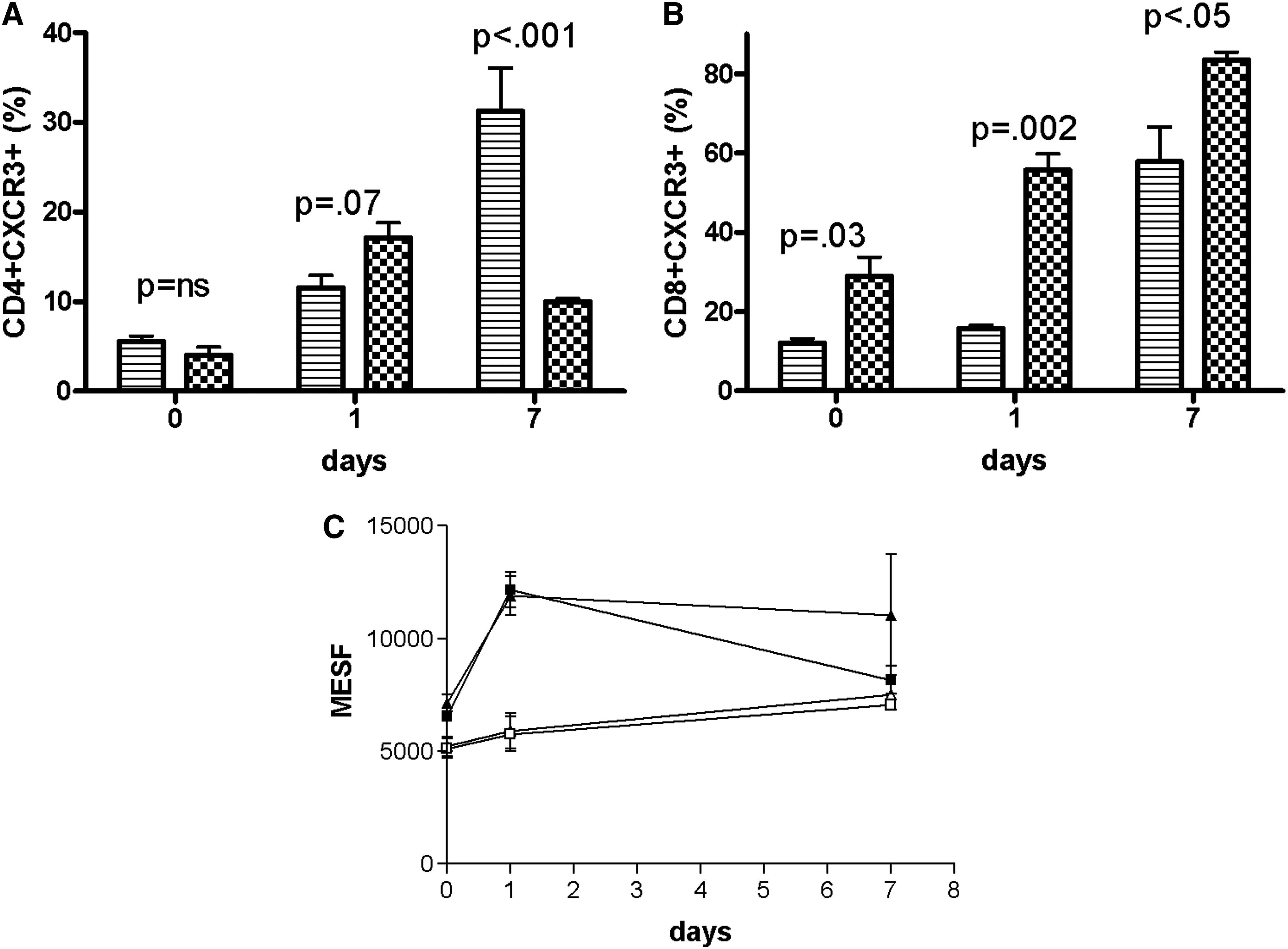
Cell activation with PHA elicited different responses on CD4 cells from control and HIV/HCV-infected individuals. A representative flow cytometric dot plot from an individual control and coinfected patient are shown in Fig. 1A. Among controls, the percent of CD4+CXCR3+ cells increased from 5% to 10% (p=0.001) after 24 h of PHA stimulation, and to 30% (p<0.001) after 7 d of stimulation. In contrast, the percent of CD4+CXCR3+ cells from coinfected patients (Fig. 1B) increased from 4% to 17% (p=0.001) after 24 hrs of PHA stimulation, but failed to increase with continued cultivation. Between the non-infected and coinfected groups, the frequency of CD4+CXCR3+ cells was equivalent before (p=ns) and after (p=0.07) 24 h of PHA stimulation (Fig. 2A). However, the frequency of CD4+CXCR3+ cells was greater (p<0.001) among controls than HIV/HCV individuals after 7 d of cultivation.
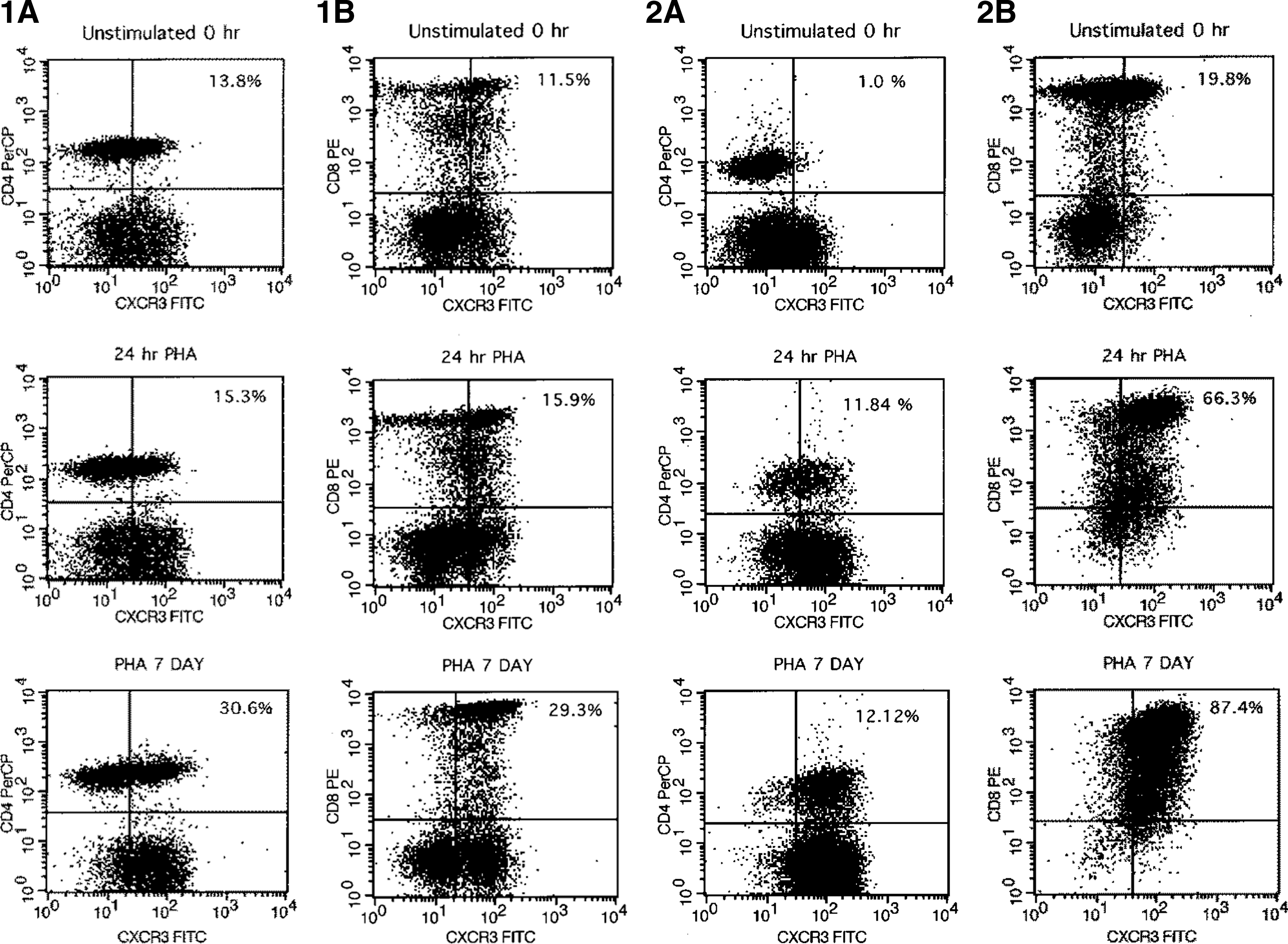
Representative dot plots from a control and coinfected individual. Representative dot plots of the three time points are shown. Cells were cultivated, stained, and CXCR3+ T-lymphocytes enumerated as described in the text. The relevant population and the percent dually-stained positive cells are displayed in the upper right quadrant of each dot plot. Panels
(
Activation-induced changes in receptor expression differed among CD8 cells (Fig. 2B). The frequency of CD8+CXCR3+ cells from controls was equivalent before and after 24 h of PHA stimulation (12±3.6% versus 15±2.4%, p=ns) but increased with 7 d of cultivation (57%, p=0.002). In contrast, the frequency of CD8+CXCR3+ cells from coinfected patients increased within 24 h of PHA stimulation (29% versus 55%, p=0.008), and escalated to 83% after 7 d of cultivation (p<0.001). Comparison between the groups showed that coinfected patients consistently exhibited a greater proportion of CD8+CXCR3+ cells than controls before (p=0.03), at 24 h (p=0.002), and at 7 d (p<0.05) after PHA stimulation.
Comparison of receptor density between patient groups was also determined (Fig. 2C). MESF values were equivalent (p=ns) between CD4 and CD8 cells at all time points for both control and HIV/HCV individuals. Prior to activation, MESF values on resting CD4 and CD8 cells from coinfected patients were slightly lower (p=0.01) than controls. PHA stimulation for 24 h caused MESF values to increase on CD4 and CD8 cells from controls (p<0.001), but not coinfected patients (p=ns). However, continued cultivation resulted in increased MESF levels among HIV/HCV patients (p=0.008). MESF levels from control cells declined after 7 d of cultivation (p<0.01), and were equivalent to levels from coinfected patients (p=ns). Among both patient groups, MESF values were greater after 7 of PHA stimulation than at resting levels.
Impact of exogenous IP10 on CXCR3 expression
Coinfected patients have higher levels of circulating IP10 than normal individuals. In addition, inflammation leading to fibrosis is greater among coinfected patients with the highest IP10 levels (2,11,15,29 –31). The possibility that elevated IP10 levels might cause receptor upregulation and enhanced inflammation was tested by measuring the impact of various IP10 concentrations upon receptor expression. Results from a representative experiment using unstimulated control cells treated with IP10 for 4 h are shown (Fig. 3). CD8+CXCR3+ cells were more prevalent than CD4+CXCR3+ cells (p<0.001). Addition of IP10 in concentrations up to 1000 ng/mL did not affect CXCR3+ expression on CD4 or CD8 cells (p=ns). MESF levels were equivalent between CD4 and CD8 cells (p=ns), and were unaffected by exogenous addition of 100–1000 ng/mL IP10. Various concentrations of IP10 were included in all the experiments described in the previous section. Since IP10 failed to alter any parameter for either patient group, results are not shown.
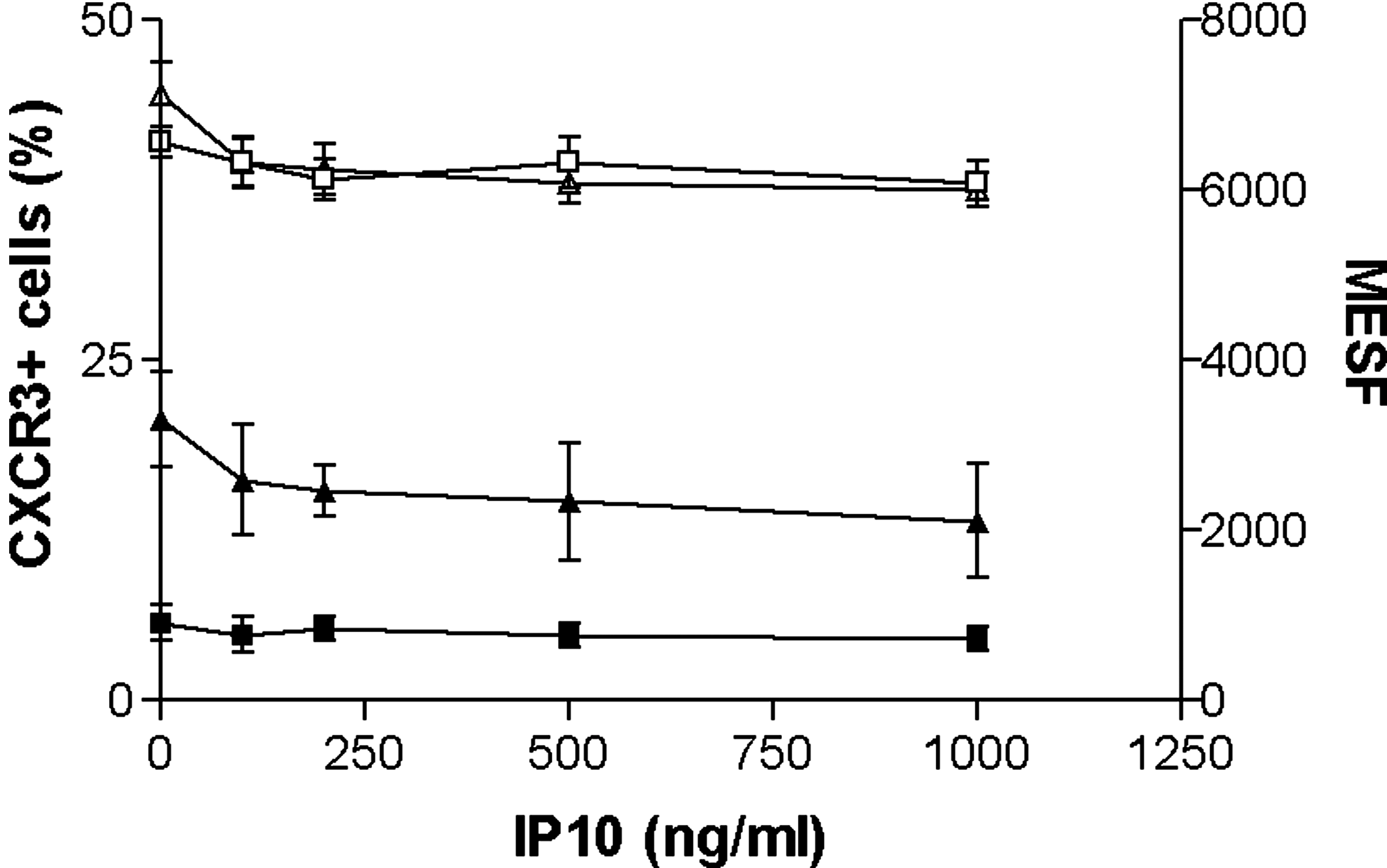
Impact of exogenous IP10 upon CXCR3 expression. (
Functional assays
PMNC functional responses were evaluated by soluble cytokine secretion and EliSpot assay. PMNCs from both controls and coinfected patients showed a Th-1-cytokine profile; however, the secretion levels differed markedly (Table 3). HIV/HCV patients secreted greater amounts of IL-2 (p=0.01), IFN-γ (p=0.001), and TNF-α (p=0.006) than controls. In contrast, IL-10 production was equivalent between groups. Inclusion of exogenous IP10 did not impact cytokine elaboration (data not shown).
Cryopreserved peripheral mononuclear cells (PMNCs) were incubated for 24 h in complete media at 37°C to allow cell recovery. Cells were then stimulated with phytohemaglutinnin (PHA) for 24 h as described in the text. Supernatants were collected and soluble cytokine levels were determined by commercial ELISA. In separate experiments, exogenous IP10 (100–1000 ng/mL) was added during PHA stimulation. However, since this did not change cytokine elaboration (p=ns), data are not shown.
HCV, hepatitis C virus.
To determine if the high concentration of Th-1 cytokines from HIV/HCV PMNCs reflected a higher proportion of cytokine-secreting cells, the number of cells elaborating IFN-γ was measured using EliSpot assay (Table 4). The frequency of IFN-γ-producing cells among unstimulated PMNCs was equivalent between patient groups (p=ns). PHA stimulation increased the number of IFN-γ-producing cells in both groups (p=0.004). However, the frequency remained equivalent between controls and HIV/HCV patients (p=ns). Inclusion of IP10 for 24 h did not change the frequency of cytokine-producing cells in controls or HIV/HCV patients (p=ns).
Cryopreserved peripheral mononuclear cells (PMNCs) were allowed to recover for 24 h in complete media at 37°C prior to the experiments. Cells (5×°105) were cultivated in triplicate for 24 h in EliSpot wells both with and without phytohemaglutinnin (PHA), stained, and enumerated following the manufacturer's instructions. Data are included for stimulated cells treated with 500 ng/mL IP10 for 24 h.
HCV, hepatitis C virus.
Proliferation
Considerable intra-individual variation was seen in both the HIV/HCV and control groups. However, the average isotopic incorporation was similar between controls and HIV/HCV patients (47,674±31,827 cpm versus 41,063±33,326 cpm, p=ns). End-of-assay viability was >85% in both groups (p=ns). Thus, differences in proliferative ability or viability do not appear to be responsible for the atypical CXCR3 expression and cytokine elaboration seen among coinfected individuals.
Discussion
The interaction between circulating chemokines and unique cell-bound receptors culminates in the migration of specific immune cells to the liver during hepatic inflammation. The chemokine IP10 and its receptor CXCR3 appear particularly responsible for lymphocyte trafficking to the liver during HCV infection (3,4,7,9,11,15,17,18,22). The magnitude of lymphocyte recruitment to the liver depends upon (1) the level of circulating IP10, and (2) the number of receptor-positive cells. Evidence abounds that systemic increases in IP10 levels are associated with greater lymphocytic infiltration and liver damage during HCV infection among both HCV- and HIV/HCV-coinfected patients (2,3,6,9 –12,18,20,23,24,26,30). However, little is known regarding the availability of receptor-positive lymphocytes among healthy or virally-infected individuals. We speculated that coinfected patients might have an atypical presentation of CXCR3 that could contribute to their propensity for hepatic inflammation. This premise was explored by comparing CXCR3 content on resting and activated CD4+ and CD8+ cells from HIV/HCV-coinfected and healthy non-infected individuals.
It has been suggested that CXCR3 expression is high on effector/memory cells and low/absent on naive cells. However, a review of the literature shows that limited data on receptor expression are actually available. The sparse data available varies considerably due to technical differences between studies. In addition, few studies have examined phenotype on both CD4+ and CD8+ populations, or under various activational conditions (6,16,19). Thus, systematic assessment of CXCR3 expression on CD4+ and CD8+ lymphocytes during activation was needed in order to identify potential differences between healthy and virally-infected subjects. We initially tested the frequency and density of CXCR3 on resting, unstimulated lymphocytes from healthy and coinfected individuals. Certain features were common to both groups. First, the majority of resting lymphocytes were receptor-free in both groups, with less than 10% of CD4+ and 30% of CD8+ cells expressing CXCR3. This observation supports the idea that receptor frequency is relatively low on resting naive cells. Second, the frequency of receptor-positive CD8+ cells was more than twofold greater than CD4+ cells among both groups. Several studies, but not all, similarly observed that CXCR3 phenotype was more prevalent among CD8 than CD4 cells (6,19). The differential receptor content may summon more CD8+ than CD4+ cells to the liver during inflammation among coinfected individuals. Although outside the scope of this study, that speculation would explain the histologic prevalence of intrahepatic CD8+ cells during HCV infection, and may be a fundamental mechanism of host attempts at viral eradication (6,9,11,12,17). In contrast to the similar frequency of resting CD4+CXCR3+ cells between patient groups, the frequency of CD8+CXCR3+ cells differed significantly. Coinfected patients showed approximately twofold greater frequency of CD8+CXCR3+ cells than their non-infected counterparts, which to the best of our knowledge is a novel observation. This elevated frequency may produce a more robust response to IP10 recruitment among coinfected individuals. Unfortunately, since HCV monoinfected patients were not included in this study, we cannot predict whether variation in receptor phenotype exists between mono- and coinfected patients. Despite variations in the frequency of receptor-expressing cells seen between patient groups, receptor density was equivalent between patient groups and lymphocyte subpopulations. Perney et al. (16) similarly observed that CXCR3 density on CD4+ cells was equivalent between healthy and HCV-infected individuals. These results suggest that receptor density may be relatively constant and non-contributory to lymphocyte recruitment by elevated IP10 levels.
Our study next evaluated CXCR3 frequency and density among lymphocytes during early and late activation, and found significant variations between healthy and coinfected individuals. Among healthy individuals, the frequency of CD4+CXCR3+ cells doubled during early activation and continued to expand during late activation. This result is similar to a study showing that CXCR3 expression on CD4 cells from various healthy lymphoid tissues rapidly expanded during early and late activation (18). In contrast, CD8+CXCR3+ frequency among healthy individuals did not expand until late activation. Lymphocytes from coinfected individuals displayed a different CXCR3 pattern. Although CD4+CXCR3+ frequency among coinfected individuals increased during early activation, levels plateaued and failed to expand during late activation. This underexpression is likely a consequence of HIV infection and suggests that T-helper responses, already compromised due to HIV, are additionally marginalized during IP10 recruitment, and may fail to appropriately traffic or offer T-cell help at the inflammatory site. In contrast, the frequency of CD8+CXCR3+ cells showed explosive increases with activation. Within 24 h of activation, the frequency of CD8+CXCR3+ cells doubled, and by late activation virtually all CD8+ cells expressed receptor. These novel observations suggest that CD8 cells from HIV/HCV coinfected patients are primed to respond more vigorously to IP10 than non-infected individuals. These intrinsic differences may contribute to the intensity of response to hepatic inflammation and clinical outcome among coinfected patients. Because it has been speculated that CXCR3 expression is higher on memory cells than naïve cells, it is tempting to interpret the different profiles between healthy and coinfected individuals as showing that peripheral lymphocytes are primarily naïve in healthy individuals, and primarily are memory/effector cells among the coinfected. However, Rabin et al. (18) showed that the relation between CD4+ memory cells and CXCR3 expression is complex and regulated by whether cells are activated by Th-1 or Th-2 pathways. Studies to determine the relationship between receptor content and naive/memory cell status among healthy and coinfected individuals are currently in progress.
To determine if IP10 exerted an autoregulatory effect upon receptor expression, the impact of physiologic concentrations of exogenous IP10 was evaluated in all experiments. However, various concentrations of IP10 did not affect any of the parameters tested in CD4+ or CD8+ cells from healthy or coinfected individuals. This implies that in vivo elevations of IP10 do not induce receptor upregulation. However, it is important to consider that inflammation causes a cytokine/chemokine “storm,” of which IP10 is only one component. Apolinario et al. (1) showed that IP10 gene expression in hepatocytes could be positively or negatively modulated by exogenous cytokines. Thus it is possible that CXCR3 expression in vivo may respond to changing IP10 levels when coupled with inflammatory cytokines.
Lastly, we compared lymphocyte function between healthy and coinfected individuals by evaluating cytokine release following activation. Both groups elaborated a Th-1 cytokine profile in response to mitogenic challenge; however, cytokine levels from coinfected patients were significantly higher than those in non-infected individuals. This is consistent with previous observations that coinfected patients produce elevated levels of Th-1 cytokines (17). Using the EliSpot assay, we determined that elevated IFN-γ resulted from greater production of IFN-γ per cell instead of a greater number of cells producing the cytokine. The biological consequences of this may be significant. Elevated production of Th-1 cytokines among infiltrating CD8+ cells could promote hepatic damage during liver inflammation. Alternatively, this may be a manifestation of functional disequilibrium in which cytokine functions are abnormal and promote liver fibrosis.
In summary, we have shown that CXCR3 expression and upregulation on CD8+ lymphocytes from HIV/HCV-coinfected individuals differs significantly from healthy individuals. Whether atypical CXCR3 presentation among coinfected individuals impacts clinical outcome remains to be elucidated.
Footnotes
Author Disclosure Statement
No competing financial interests exist.