
Abstract
Here we examined whether polymeric IgA (pIgA) and monomeric IgA (mIgA) antibodies differ in their ability to neutralize drift viruses within the same subtype. We used an IgA monoclonal antibody (mAb; H1-21) against influenza virus strain A/Hiroshima/52/2005 (A/Hiroshima; H3N2). The mAb was obtained after immunizing mice mucosally with a split-virion (SV) vaccine. The mAb contained both mIgA and pIgA forms. It reacted with the homologous virus and cross-reacted with drift viruses A/New York/55/2004 (H3N2) and A/Wyoming/3/2003 (H3N2) in hemagglutinin-inhibition (HI) and neutralizing Ab assays. The mAb also cross-reacted with A/Panama/2007/99 (H3N2) in an ELISA. We separated the mAb into pIgA and mIgA fractions by gel filtration, and then tested them for neutralizing Ab activity. The neutralizing activity for the A/Hiroshima/52/2005, A/New York/55/2004, and A/Wyoming/3/2003 viruses was lower for the mIgA than the pIgA fraction. However, the neutralizing efficiency for drift variants relative to that for the homotype did not differ between pIgA and mIgA, and pIgA only neutralized variants that could also be neutralized by mIgA. These results suggest that the polymerization of IgA enhances its antiviral immune responses, but does not increase the number of influenza virus strains neutralized by the IgA.
Introduction
In animal models, intranasal immunization with an influenza virus split-virion (SV) vaccine mixed with a mucosal adjuvant induces cross-protection against drift variants within a subtype and against different subtypes (6,22,24,25). Intranasal immunization, but not subcutaneous or intraperitoneal immunization, induces cross-reactive IgA antibody production in the respiratory tract, suggesting that cross-protection may be associated with sufficient levels of secreted IgA antibodies (23). IgA, which is the major contributor to humoral mucosal immunity, occurs in secretions as a polymer-bearing secretory component (9,17). Secretory IgA (S-IgA) can effectively neutralize viruses at mucosal surfaces and displays antibacterial activity (8,17). Although polymer-bearing IgA without the secretory component is less stable than the S-IgA molecule, being more susceptible to chemical or enzymatic cleavage, adding the secretory component does not influence its anti-virus activity either positively or negatively. These findings led to the suggestion that S-IgA's biological activity is associated with its antigen-binding fragment (Fab) (17).
A polymeric IgA (pIgA) monoclonal antibody (mAb) against the influenza A virus strain A/PR/8/34 (H1N1) neutralizes the homotype virus more effectively than the monomeric IgA (mIgA) form (17). However, there is no evidence as to whether a pIgA or an mIgA is more effective for cross-reactive neutralizing antibody activity. In the present study, we separated an anti-influenza IgA mAb into pIgA and mIgA on a gel filtration column, and assayed their individual activities to address this question.
Materials and Methods
Generation of hybridomas that secrete IgA mAbs against influenza
To obtain hybridomas, mice were immunized as described by Taylor et al. (26). Briefly, 8-week-old female BALB/C mice (n=5; Japan SLC, Hamamatsu, Japan) were immunized intraorally with a mixture of 500 μg SV vaccine prepared from A/Hiroshima/52/2005 (H3N2) and 40 μg of pertussis toxin (PT), on days 0, 16, and 34. On day 55, the immunized mice were given the SV vaccine intraorally (500 μg) and intravenously (40 μg). Spleen lymphocytes harvested from the mice on day 58 were fused with SP2/O mouse myeloma cells. Hybridomas secreting neutralizing mAb against A/Hiroshima/52/2005 were selected by an anti-IgA ELISA. ELISA screening for mAb cross-reactivity to other strains identified H1-21 as a cross-reactive IgA mAb. The hybridoma-producing H1-21 was cloned twice by limiting dilution. The cloned H1-21 hybridoma was injected intraperitoneally into BALB/c nu/nu mice (Japan SLC) to generate mAbs in the ascitic fluid, as previously described (4). The H1-21 IgA mAb was purified from the ascitic fluid by affinity chromatography using the AKTA purifier system (GE Healthcare, Little Chalfont, U.K.) with a KAPTIV-AE column (Tecnogen, Pontenure, Italy). The separation conditions used were as indicated by the manufacturer. The H1-21 polymeric and monomeric forms were then purified by size exclusion chromatography using the AKTA purifier system with a Superdex™ 200 10/300 GL column and fractional collector Frac-950 (GE Healthcare). The separation conditions were as follows: the mobile phase was PBS with a flow rate of 0.5 mL/min at room temperature, and the fractions were 0.3 mL each.
SDS-PAGE and Western blotting
Non-reducing gel electrophoresis was performed in an SDS-acrylamide gel consisting of 4% acrylamide-N,N′-diallyltartardiamide (29/1), 0.1 M phosphate buffer (pH 7.0), 0.1% SDS, and 15% glycerol. The sample buffer contained 0.02 M phosphate buffer (pH 7.0), 30% glycerol, 1% SDS, and 0.1% bromophenol blue. The electrophoresis buffer contained 0.1 M phosphate buffer and 0.1% SDS. IgA samples in sample buffer (100 ng/10 μL) were heated to 100°C and loaded onto the gels. The gels were then run at 20 mA for 3 h.
The purified IgA mAbs (10 ng) were subjected to SDS-PAGE as described above. The samples in the gels were then transferred to a polyvinylidene difluoride membrane and analyzed by Western blotting using horseradish peroxidase (HRP)-conjugated goat anti-mouse IgA (KPL, Gaithersburg, MD), and enhanced chemiluminescence (GE Healthcare), followed by x-ray film exposure.
Influenza virus strains and SV vaccine
The following influenza A virus strains were used for cross-reactive hemagglutinin inhibition (HI) and neutralization tests: A/Hiroshima/52/2005 (H3N2), A/New York/55/2004 (H3N2), A/Wyoming/3/2003 (H3N2), A/Panama/2007/99 (H3N2), A/Uruguay/716/2007 (H3N2), A/New Caledonia/20/99 (H1N1), A/Solomon Islands/3/2006 (H1N1), and A/Brisbane/59/2007 (H1N1), which were kindly provided by Dr. Takato Odagiri (National Institute of Infectious Diseases, Tokyo, Japan); A/PR/8/34 (H1N1) was kindly provided by Dr. Hideki Hasegawa (National Institute of Infectious Diseases, Tokyo, Japan). Influenza virus SV vaccines for immunization and viral surface antigens for ELISA were prepared from A/Hiroshima/52/2005, A/New York/55/2004, A/Wyoming/3/2003, A/Panama/2007/99, and A/Uruguay/716/2007, at Kanonji Institute, The Research Foundation for Microbial Diseases of Osaka University.
To prepare virus stocks, confluent MDCK cell monolayers were infected with viruses at a multiplicity of infection (MOI) of 0.001 using virus dilution medium (minimum essential medium [MEM] supplemented with 0.3 mg/mL
ELISA cross-reactive antibody screening
IgA mAbs were screened for cross-reactivity with several influenza HA antigens by IgA ELISA (14). Briefly, 96-well ELISA plates were coated with 0.5 μg of influenza SV vaccine in PBS and incubated overnight at 4°C. The wells were then blocked with PBS containing 10% FCS overnight at 4°C. Next, 100 μL of threefold serial dilutions of 100 μg/mL IgA mAb H1-21 were added to the wells. After incubating the plates for 1 h at room temperature, the wells were washed and incubated with HRP-conjugated goat anti-mouse IgA antibody (Southern Biotechnology Associates, Atlanta, GA) for 1 h at room temperature. The wells were then washed again and developed with TMB solution (Moss Inc., Pasadena, NJ). After 15 min, the enzyme reaction was stopped by adding 0.5 N HCl, and the A450 of the samples was read with a microplate reader. The IgA mAb H1-21 concentration capable of inducing positive ELISA activity that yielded an A450 more than 0.1 was recorded.
Measurement of HI activity
The HI assay, a standard method for the serologic detection of influenza virus infections, was performed. One part H1-21 (1 mg/mL) was diluted with three parts receptor-destroying enzyme (RDE), and the mixture was incubated for 18 h in a 37°C water bath, then for 30 min at 56°C to inactivate the enzyme. Next, 6 parts of 0.85% physiological saline was added for a final dilution of 1:10 (100 μg/mL). HI assays using 50 μL of threefold serial dilutions of 100 μg/mL-treated H1-21 were performed in U-bottom 96-well microtiter plates with 0.6% guinea pig erythrocytes by a standard method (15,19). The IgA mAb H1-21 concentration capable of inducing HI activity was recorded.
Anti-influenza neutralizing antibody titer determination
The IgA mAb neutralizing antibody titers were determined by a micro-CPE neutralizing test. H1-21 IgA mAbs (1 mg/mL) were treated with RDE as described above. One hundred microliters of threefold serial dilutions of 100 μg/mL H1-21 IgA mAb was mixed 1:1 (v/v) with a suspension containing 100 TCID50 of various influenza virus strains, and the mixture was incubated at 37°C for 1 h. The mixtures were transferred onto MDCK cell monolayers in 96-well plates and incubated at 37°C for 1 h. The cells were then washed with MEM supplemented with 0.01 M HEPES, 0.2% bovine albumin, and 1 μg/mL trypsin (MEM maintenance medium), after which 100 μl of MEM maintenance medium was added to each well, and the cells were incubated at 37°C in a 5% CO2 incubator. The CPE status was observed 96 h after infection. The culture medium was removed by absorption, and adherent MDCK cells were fixed with 4% formaldehyde-PBS for more than 1 h. The PBS was removed by absorption, and the fixed cells were stained with 0.1% naphthol blue black for 30 min. The plates were washed, 50 μL of 0.1 M NaOH was added to each well, and the A595 was read using a microplate reader. The neutralizing antibody titer was expressed as the concentration of IgA mAb H1-21 that neutralized 50% of 100 TCID50 of virus.
Sequence analysis of the HA gene
Viral RNA was extracted from the influenza virus stock as described above using TRIzol reagent (Life Technologies, Carlsbad, CA), and reverse-transcribed with the Uni 12 primer (5), using the SS III RT kit (Life Technologies). Viral genes were amplified by polymerase chain reaction (PCR) with KOD-Plus-DNA polymerase (Toyobo, Osaka, Japan). Primers for the H3 HA gene were as follows: forward primer 5′-TATTCGTCTCAGGGAGCAAAAGCAGGGGATAATTC-3′ and reverse primer 5′- ATATCGTCTCGTATTAGTAGAAACAAGGGTGTTTTTAATTAATGC-3′. The HA gene was directly sequenced using an ABI 3500 genetic analyzer (Life Technologies), with the above primers and HA 780-F-SQ 5′-GCAGAATAAGCATCTATTGG-3′.
Statistical evaluation
Nonparametric Student's t-tests were performed using Statcel2 software (OMS, Tokyo, Japan) to evaluate differences between groups in the experiments. A p value<0.05 was considered significant.
Results
Characterization of the anti-influenza IgA mAb produced by hybridoma H1-21
The H1-21 IgA mAb against A/Hiroshima/52/2005 (H3N2) was harvested from mice that had been intraorally immunized with an influenza SV vaccine plus PT. The cloned H1-21 hybridoma was injected intraperitoneally into BALB/c nu/nu mice to generate mAbs in the ascitic fluid, and the IgA mAb was purified from the ascitic fluid with a KAPTIV-AE column. Non-reducing SDS-PAGE showed that the IgA mAb from H1-21 formed both monomers and polymers, including the H3L3 form (21) (Fig. 1A, left panel), and these bands were hybridized by an anti-mouse IgA mAb in Western blots (Fig. 1A, right panel).
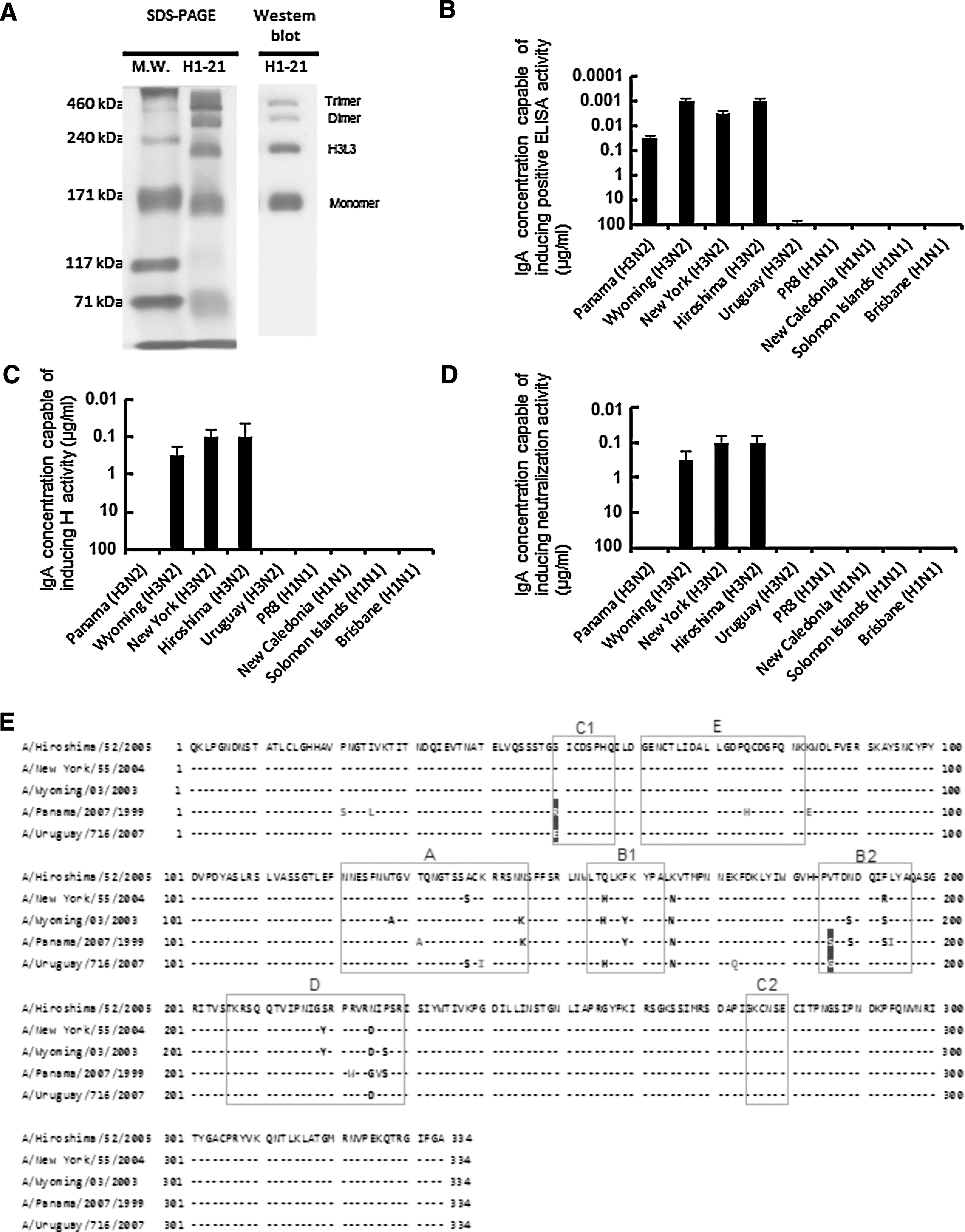
Structure and cross-reactivity of the anti-influenza IgA mAb H1-21. (
ELISAs for influenza virus antigen cross-reactivity showed that H1-21 bound to virus surface antigens of A/Panama/2007/99, A/Wyoming/3/2003, and A/New York/55/2004, which are drift variants of A/Hiroshima/52/2005 (Fig. 1B). However, H1-21 did not bind to A/Uruguay/716/2007 (H3N2) or H1N1-strain antigens (Fig. 1B). Cross-reactive HI tests showed that H1-21 inhibited the HA activity in A/Hiroshima/52/2005, A/New York/55/2004, and A/Wyoming/3/2003 (Fig. 1C); however, it did not inhibit the activity in the A/Panama/2007/99 and A/Uruguay/716/2007 variants or in H1N1 strains. The three strains in which HA activity was inhibited by the H1-21 IgA mAb were neutralized by the mAb (Fig. 1D).
We further compared the deduced amino acid sequence of HA1 among the strains. As shown in Fig. 1E, A/Hiroshima/52/2005, A/New York/55/2004, and A/Wyoming/3/2003, which were neutralized by H1-21, had the same amino acid at positions 50 and 186 (G and V, respectively). However, A/Panama/2007/99 and A/Uruguay/716/2007, which were not neutralized by H1-21, had different amino acids at positions 50 and 186 (R/E and S/G, respectively, in the dark-colored boxes in Fig. 1E). Furthermore, in A/Panama/2007/99 the amino acids at positions 21, 25, 75, 83, 131, 194, 222, and 226 were different from those of the other four strains (Fig. 1E), and in A/Uruguay/716/2007, the amino acids at positions 140 and 173 were different from those of the other four strains (Fig. 1E).
Effect of IgA polymerization on cross-reactive antibody responses
Renegar et al. (17) demonstrated that a pIgA mAb against A/PR/8/34 was more effective than the mIgA in neutralizing the homotype virus. However, it has not been clarified whether pIgA is also more effective than mIgA in cross-reactive neutralization. We addressed this question by separating the H1-21 IgA mAb monomer and polymer (H3L3, dimer, and trimer) forms by gel chromatography (Fig. 2A), and comparing their activities. Assays showed that pIgA's cross-reactive neutralizing activity was significantly more effective than that of mIgA (Fig. 2B). However, neither pIgA nor mIgA cross-reacted with the variants A/Panama/2007/99 and A/Uruguay/716/2007.
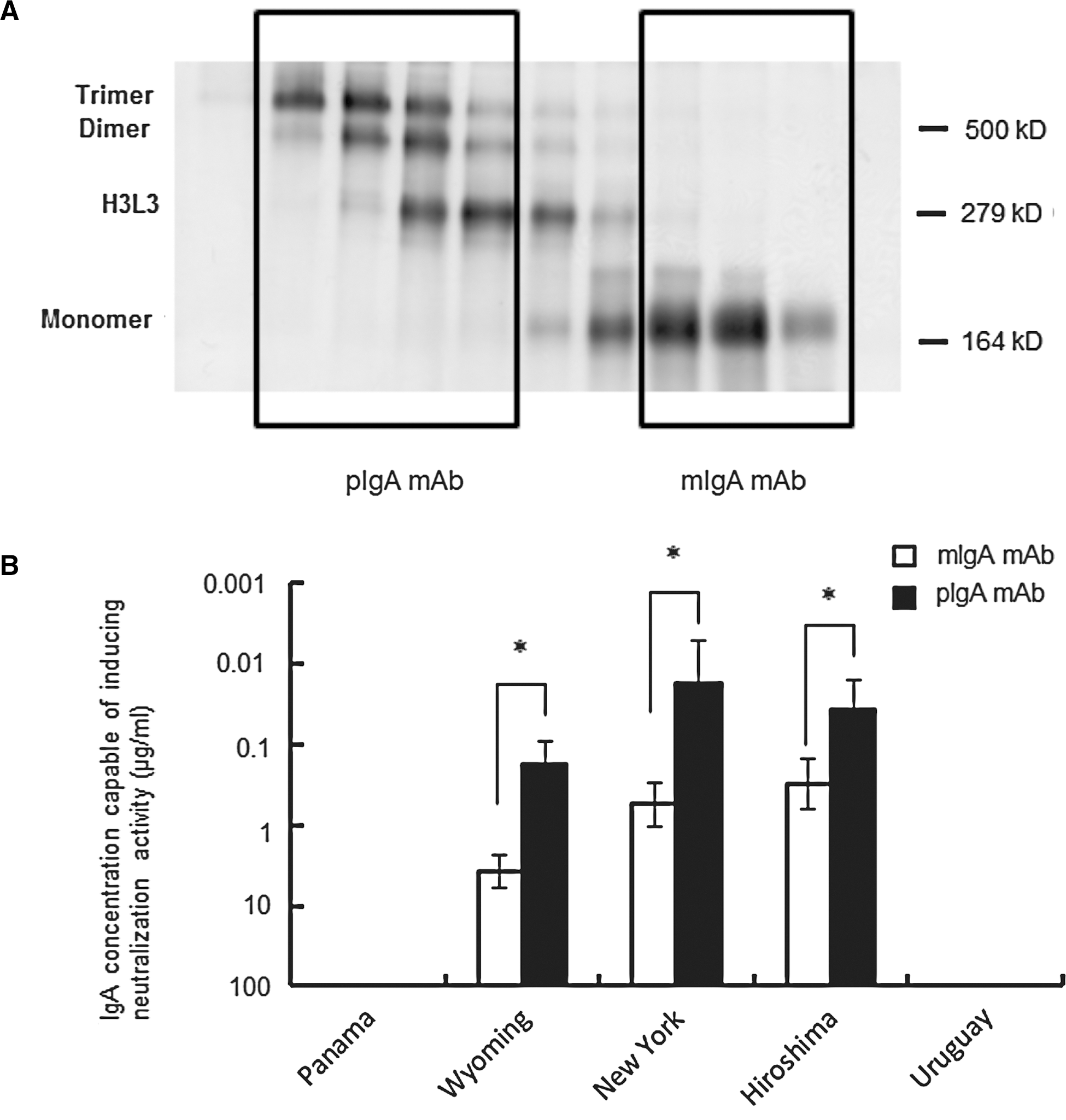
Separation of pIgA and mIgA and comparison of their neutralizing antibody activity. (
Discussion
In the present study, we examined whether pIgA and mIgA differ in their ability to cross-neutralize variant influenza viruses, using an IgA mAb H1-21 against the A/Hiroshima/52/2005 (H3N2) strain. The H1-21 IgA mAb, composed of both mIgA and pIgA forms, was one of several mAbs obtained after immunizing mice mucosally with various influenza split-virion vaccines. This mAb had neutralizing activity resulting from its recognition of the viral HA. The H1-21 IgA mAb reacted with the homologous virus and cross-reacted with the drift viruses A/New York/55/2004 and A/Wyoming/3/2003 in HI and neutralizing antibody assays.
There are several reports in which mucosal immunization with an influenza vaccine has induced cross-reactive immune responses against the homologous virus, drift variants within the subtype, and different subtypes of influenza viruses (6,22,24,25). In these reports, the cross-reactive antibody responses could be detected by ELISA, but not by HI or neutralizing antibody assays. Thus cross-reactive antibodies against variant viruses can be detected more broadly by ELISA than by HI or neutralizing antibody assays. While the roles of the epitope detected by HI or neutralizing antibody assays in protecting the host against virus infection are known, that of the epitope detected by ELISA remains to be discovered.
In the present study, we compared the deduced amino acid sequences of HA1, and found that the amino acids at positions 50 and 186 were different in A/Panama/2007/99 and A/Uruguay/716/2007, which were not neutralized by H1-21 (Fig. 1E). Position 50 is in the C1 neutralizing epitope for the IgG antibody, and position 186 is in the B2 neutralizing epitope for the IgG antibody (dark-colored boxes in Fig. 1E) (13,28). These results led us to speculate that these two positions are associated with the IgA mAb H1-21's recognition of HA for HI and virus neutralization. Furthermore, since positions 140 and 173 in A/Uruguay/716/2007 were different from those of the other four H3N2 viruses, an amino acid change at position 140 or 173 might be involved in the reduced ELISA antibody activity of H1-21. Further study to determine whether the C1 and B2 epitopes represent the IgA mAb H1-21 binding site on the respective virus HA molecules will help clarify the mechanism behind the cross-reactivities between the homologous virus and variant viruses.
IgA mAbs form both monomers and polymers, including the H3L3 form (21), and it has been speculated that not only increased antigen-binding sites on IgA molecules, but also the polymerization of mIgA, is important for enhancing antiviral protection (1,17,18). Polymeric anti-A/PR/8/34 (H1N1) HA IgA mAbs were reported to be 5–10 times more effective than the monomeric form for neutralizing the A/PR/8/34 virus (17). However, there is no direct evidence of whether pIgA or mIgA Ab is more effective in cross-reactive viral neutralization. In the present study, we found that the pIgA mAb was more effective than the mIgA mAb for eliciting cross-reactive virus neutralization. However, the relative neutralizing activity for the drift variants versus the homotype virus did not differ between the polymer and monomer forms. Furthermore, the polymer form only neutralized drift variants that could also be neutralized by the monomer form. These results suggest that IgA polymerization enhances the antibody reactivity against viruses, but does not make the antibody effective against a greater number of strains. The mechanism by which IgA polymerization enhances antibody responses remains elusive.
In animal models, cytotoxic T lymphocytes, IgG, and secretory IgA (S-IgA) antibodies contribute to the cross-protective immunity against influenza; cytotoxic T lymphocytes eliminate infected cells, while the S-IgA and IgG antibodies help eliminate viruses from the mucosae of the nose, trachea, and lungs (5,6,16,30). S-IgA is composed of pIgA and a secretory component, part of the polymeric Ig receptor (pIgR). Polymeric IgA antibodies are produced by IgA antibody-producing cells in the lamina propria, transported by the pIgR from the basolateral to the apical surface of mucosal epithelial cells, and secreted as S-IgA (7,9,18). However, polymorphic IgA without a secretory component is reported to have the same virus-clearance activity as S-IgA (18).
In the present study, we demonstrated that the pIgA mAb (H1-21) had cross-reactive neutralizing antibody activity against drift viruses. Thus the ability of pIgA to neutralize drift viruses could be similar to that displayed by S-IgA for cross-protection against a drift virus infection. Current parenteral influenza split-virus vaccines only stimulate systemic IgG production, which is less cross-protective than pIgA (3,10,29). Since cross-reactive pIgA could elicit cross-protection against influenza strains, further studies on mucosal vaccines that produce IgA Abs will be important for developing influenza vaccines that are more effective than the current parenteral ones.
Footnotes
Acknowledgments
We thank Drs. Takato Odagiri and Hideki Hasegawa for providing influenza virus strains, and Dr. Sumiko Matsuoka for technical assistance. This study was partly supported by grants from the Japanese Ministry of Health, Welfare, and Labor, and the Japanese Ministry of Education, Culture, Sports, Science, and Technology.
Author Disclosure Statement
No competing financial interests exist.