
Abstract
Vaccination is the most effective way to control swine influenza virus (SIV) in the field. Classical vaccines are based on inactivated antigens formulated with an oil emulsion or a polymeric adjuvant. Standard adjuvants enhance the humoral response and orient the immune response toward a Th2 response. An important issue is that current vaccines do not protect against new strains. One approach to improve cross-protection is to enhance Th1 and cytotoxic responses. The development of adjuvants orienting the immune response of inactivated vaccines toward Th1/Cytotoxic responses would be highly beneficial. This study shows that the water in oil in water emulsion adjuvant Montanide™ ISA 201 VG allows the induction of anti-influenza CD8 T cell in mice and induces homologous protection against an H1N1 challenge in swine. Such adjuvants that induce both humoral and cell-mediated immunity could improve the protection conferred by SIV vaccines in the field.
Introduction
V
The development of new oil adjuvants that would enhance both the humoral and cell-mediated immune response could improve the efficacy of swine and cattle vaccines further. For example, they could help to extend the vaccinal cross-protection against different viral variants or serotypes of a given pathogen. The new W/O/W adjuvant Montanide™ ISA 201 VG (ISA 201) has been developed recently, and is based on a specific light mineral oil and a mannide oleate–based surfactant system. This new formulation has been selected in order to improve the cell-mediated immune response of W/O/W vaccines while preserving a constant safety profile and humoral immune response compared with the reference adjuvant ISA 206 (Seppic, unpublished data). It has been shown recently that ISA 201 increases protection against homologous FMD challenge compared to ISA 206 and another classical W/O/W adjuvant (8).
The present study was performed to assess the humoral and cell-mediated immunity conferred by ISA 201 in a H1N1 influenza vaccine in a mouse model and in the target species, swine, using the naturally occurring swine H1N1 virus (A/swine/California/07/09), which caused the last human pandemic (3,12). Swine influenza is a major disease of swine that causes production losses in the pig industry and is also still a concern for human health. Swine influenza virus (SIV) is currently controlled in pig farming through vaccination with adjuvanted vaccines containing inactivated whole virus, which induce a mostly humoral response against antigenic influenza proteins such as the hemagglutinin protein. Vaccinal strains should therefore be matched closely to the field viral strains to ensure protection, even though it is not legally required of SIV vaccine manufacturers. In spite of the fact that antigenic drift and reassortment is not as frequent in SIV as in human influenza strains, some extent of antigenic and genetic drift has been observed (9,22), and thus vaccines could become nonprotective against field strains (17). In order to broaden the protection against SIV variants, one option is to induce a cytotoxic T cell response against conserved internal influenza proteins, such as nucleoprotein (NP) (4,26). Using new adjuvants that enhance the cytotoxic response of the vaccine may therefore be of interest for SIV vaccination.
In this study, the humoral and cell-mediated immunity conferred by ISA 201 in a H1N1 influenza vaccine was assessed in mice, and enhancement of the protection against pandemic H1N1 SIV was confirmed in swine.
Materials and Methods
Ethics statement
Mice experiments were carried out in accordance with the Institut National de la Recherche Agronomique (INRA) guidelines in compliance with European animal welfare regulation. The protocol was approved by the Animal Care and Use Committee at Centre de Recherche de Jouy-en-Josas under relevant institutional (DSV, permit number: 78–112) and INRA “Santé Animale” department guidelines. All pig experiments were approved by the Federal Centre for Animal Health (FGBI ARRIAH Bioethical committee; reference 169, 5/9/2006) in accordance with Russian Federation law and the norms of the Institute of Laboratory Animal Resources, National Research Council (NRC), United States. All experimental procedures were performed in Biosafety level 2 facilities.
Mice study: vaccines
Influenza A virus strain A/Puerto Rico/8/1934 H1N1 virus (H1N1) was amplified on eggs according to standard procedures. The virus was inactivated by incubation with formalin for 4 days at 4°C (25), and then purified on sucrose gradient. Inactivation of the virus was tested using plaque assay. Serial 10-fold dilutions of the virus were allowed to adsorb onto confluent monolayers of a Madin–Darby canine kidney (MDCK) cell line on a 24-well plate for 1 h. The supernatant was then removed and replaced with 1% agarose (Invitrogen) supplemented with serum-free Dulbecco's modified Eagle's medium (DMEM; Lonza) and 2 mg/ml L-1-tosylamido-2-phenylethyl chloromethyl ketone-treated trypsin. Plates were incubated for 3 days at 37°C with 5% CO2. Agarose overlay was then removed, and the plaques were visualized by addition of crystal violet (Sigma-Aldrich). Adjuvants Montanide™ ISA 206 VG (ISA 206) and Montanide™ ISA 201 VG (ISA 201; Seppic) were formulated following the manufacturer's recommendations with inactivated virus in NaCl (aqueous phase) at a 50/50 (weight/weight) ratio. ISA 201 and ISA 206 are two different W/O/W emulsion adjuvants composed of a light mineral oil and a refined emulsifier system obtained from mannitol and purified oleic acid of vegetable origin. ISA 201 has been developed recently and is based on a specific enriched light mineral oil and an extremely refined mannide oleate–based surfactant system.
Mice study: animals and vaccination groups
Thirty-six C57BL6 female mice were purchased from the Centre d'Elevage R. Janvier, and were used around 8 weeks of age. Mice were anesthetized with a mixture of ketamine and xylazine (1 and 0.2 mg per mouse, respectively). Mice were vaccinated subcutaneously at day 0 with 1 μg inactivated virus in 50 μL NaCl solution or V/V NaCl/adjuvant. The infected group was infected intranasally with 50 μL of 0.9% endotoxin-free NaCl containing 500 plaque forming units (PFU) of A/PR8/34 in BSL2 confinement. Vaccinated groups were boosted at day 21 post-vaccination under the same conditions as for day 0. Sera were obtained from blood collected via cheek puncture 14 days after the prime and 14 days after the boost immunizations. Mice were killed by gentle cervical dislocation.
The following vaccination groups were included in the study (six mice per group): not vaccinated control group (0/0), mice vaccinated with nonadjuvanted inactivated H1N1 vaccine (inactH1N1/0), mice vaccinated with ISA206 or ISA 201 adjuvanted inactivated H1N1 vaccine (inactH1N1/ISA206 and inactH1N1/ISA201, respectively), mice injected with adjuvant only (0/ISA 206), and mice infected intranasally with live H1N1 virus (0/liveH1N1).
Mice study: IgG1 and IgG2 antibody titration
Individual mouse sera were assayed for NP or whole inactivated influenza particle-specific antibodies IgG2a or IgG1 by enzyme-linked immunosorbent assay (ELISA) using NP recombinant protein produced, as previously described (18), as coating (200 ng per well) or the same whole inactivated influenza particle used for vaccination as coating (200 ng per well). Antigen-bound antibodies were detected using horseradish peroxidase (HRP)-conjugated rat antimouse IgG1 or IgG2a (BD-Biosciences), as previously described (23). Endpoint antibody titers were calculated by regression analysis, plotting serum dilution versus the absorbance at 450 nm using Microcal Origin (OriginLab Corp.; regression curve y = (b + cx)/(1 + ax)). Endpoint titers were defined as the highest dilution resulting in an absorbance value twice that of blank points (points without immune serum).
Mice study: in vitro restimulation of spleen T lymphocytes
Freshly isolated, red blood cell depleted splenocytes were incubated with 2 μM CFSE (Life Technologies) in RPMI (Lonza) without fetal calf serum (FCS) for 10 min at 37°C and then co-cultured for 5 days with either live PR8/34 virus (1 PFU/cell) or inactivated PR8/34 virus (10 μg/mL final concentration), or with medium alone as negative control, all diluted in RPMI 1640 supplemented with 10% FCS, PS, 2 mM L-glutamine, and 50 μM β2-mercaptoethanol (Sigma). After 20 min incubation with anti-CD32/CD16 (FcBlock, BD-Biosciences), CFSE-labeled splenocytes were stained with an anti-CD3-Alexa647, anti-CD8-PE, and anti-CD4-PerCP (BD-Biosciences; 5 μg/mL) for 30 min on ice, washed, and then fixed in 10% CellFIX (BD-Biosciences) and analyzed on a FACScalibur (BD-Biosciences) collecting 100,000 events. Proliferating cells were characterized as CFSElow. As expected CFSElow lymphocytes displayed higher FSC and SSC parameters than their nondividing CFSEhigh counterpart.
Pig trial: vaccines
Formulation of the adjuvanted vaccine was performed by combining swine influenza inactivated virus (strain A/swine/California/07/09 (H1N1; FGBI ARRIAH) at 8 log EID50/mL and the adjuvant Montanide™ ISA 201 VG (Seppic) at a weight ratio of 50% of adjuvant and 50% of antigenic phase, following the manufacturers' recommendations. The nonadjuvanted vaccine was formulated by dilution of the antigen in saline buffer. Both vaccines contained the same antigenic load.
Pig trial: animals and vaccination schedule
Thirty pigs (2 months old, SIV seronegative at day 0) were separated randomly into three groups of 10 pigs. All animals were kept in quarantine for 4 days before the beginning of the trial.
Ten pigs were left untouched (group 0/0). For the two other groups, at day 0 and day 21, each pig was injected intramuscularly with either 2 mL of inactivated H1N1 with ISA201 adjuvant (group H1N1/ISA201) or 2 mL of inactivated H1N1 without adjuvant (inactH1N1/0).
Pig trial: safety assessment
After each injection, the body temperature of all animals was recorded for 5 days (at 4 h, 24 h, 48 h, 72 h, 96 h, and 120 h post-vaccination). Occurrence of systemic reactions and local reactions at the injection site was also observed for 5 days post-injection. After slaughter at day 56, injection sites were dissected to assess the presence of local reactions.
Pig trial: antibody titration
Blood samples were taken from each pig at day 0 (before vaccination), day 21 (before revaccination), day 42 (before virulent challenge), and day 56 (at slaughter). Blood sera were prepared by centrifugation of blood samples (2,000 g, 4°C) for 30 min and stored at −50°C before analysis. Specific antibody titration in blood sera was performed by hemagglutination inhibition (HI) titration kit (FGBI ARRIAH) and ELISA analysis (“Swine influenza virus antibody test kit”; IDEXX).
Pig trial: infectious challenge
At day 21, five pigs in each group were challenged intranasally with SIV strain A/swine/California/07/09 (H1N1) at 8.3 log EID50 per challenge dose (2 mL). After challenge, the body temperature of each animal was recorded for 14 days to measure hyperthermia post-infection. Nasal swabs were collected at 2, 4, 6, 8, 10, and 14 days post-challenge, and inoculated to the MDCK cell monolayer for isolation of the SIV. In case of cytopathic effect on cell monolayer, the material was tested by ELISA analysis.
Pigs were slaughtered 14 days post-challenge (at day 56 post-vaccination). After slaughter, lung lesions were scored in each animal according to Halbur et al.'s method (13). Briefly, lesions were scored in each part of the lungs, with the upper left lobe accounting for 5%, the upper right lobe 10%, the middle left lobe 10%, the middle right lobe 10%, the major left lobe 30%, the major right lobe 35%, and the cardiac lobe 5% of the total lung tissues. A total lesion score expressed in percent was assessed for each animal.
Statistics
Results are expressed as the mean ± standard deviation (SD) of six animals per group for mice, and of 10 animals per group for swine. Nonparametric Mann–Whitney U-tests were used to compare the different group means, and Wilcoxon signed rank tests were used when comparing ratios to 1 (GraphPad software). Levels of significance are indicated on the graphs by asterisks or circles (*p < 0.05; **p < 0.01; ***p < 0.001).
Results
Assessment of adjuvant immunogenic profile in mice
The immune response conferred by the vaccine based on the adjuvant ISA 201 combined with inactivated influenza virus (PR8/34) was first assessed in a mouse vaccination study. This new adjuvant was compared with reference adjuvant ISA 206. Both adjuvants are W/O/W emulsion adjuvants, and are used at the same concentration in the vaccine. Vaccines based on ISA 206 or ISA 201 were injected into mice at day 0 and day 21. The response conferred by vaccination with adjuvanted vaccines was also compared with an infection at day 0 with the live influenza virus. Infection is known to induce a Th1 biased response and a strong cytotoxic T CD8 cellular response, and was included in the study as a positive control for cellular immune response.
In all groups, IgG1 and IgG2a antibody titers to influenza N protein were determined at day 0, day 21, and day 42 post-vaccination. Both ISA 201 and ISA 206–based inactivated vaccines induced strong IgG1 and IgG2a titers at day 21 and day 42, whereas the nonadjuvanted antigen induced only low nonsignificant IgG1 titers and no IgG2a, and the adjuvant with no antigen did not induce any response (Fig. 1A and B). As expected, infection with the live virus induced the strongest IgG1 and IgG2a responses. In each group, the IgG2a/IgG1 ratio was calculated. The IgG2a/IgG1 ratio of the infected mice was significantly higher than 1 (4.2 ± 2.2, p < 0.05), indicating a Th1 biased immune response. Mice vaccinated with nonadjuvanted antigen had a ratio lower than 1 (0.3 ± 0.4, p = 0.06), which indicated a tendency toward a Th2 response. The vaccination with antigen + ISA 206 or antigen + ISA 201 induced a ratio indicative of mixed Th1/Th2 responses, 0.9 ± 0.8 and 0.9 ± 0.5 respectively (Fig. 1C). Similar results were obtained when IgG1 and IgG2a antibody titers against the whole inactivated virus were measured (data not shown).
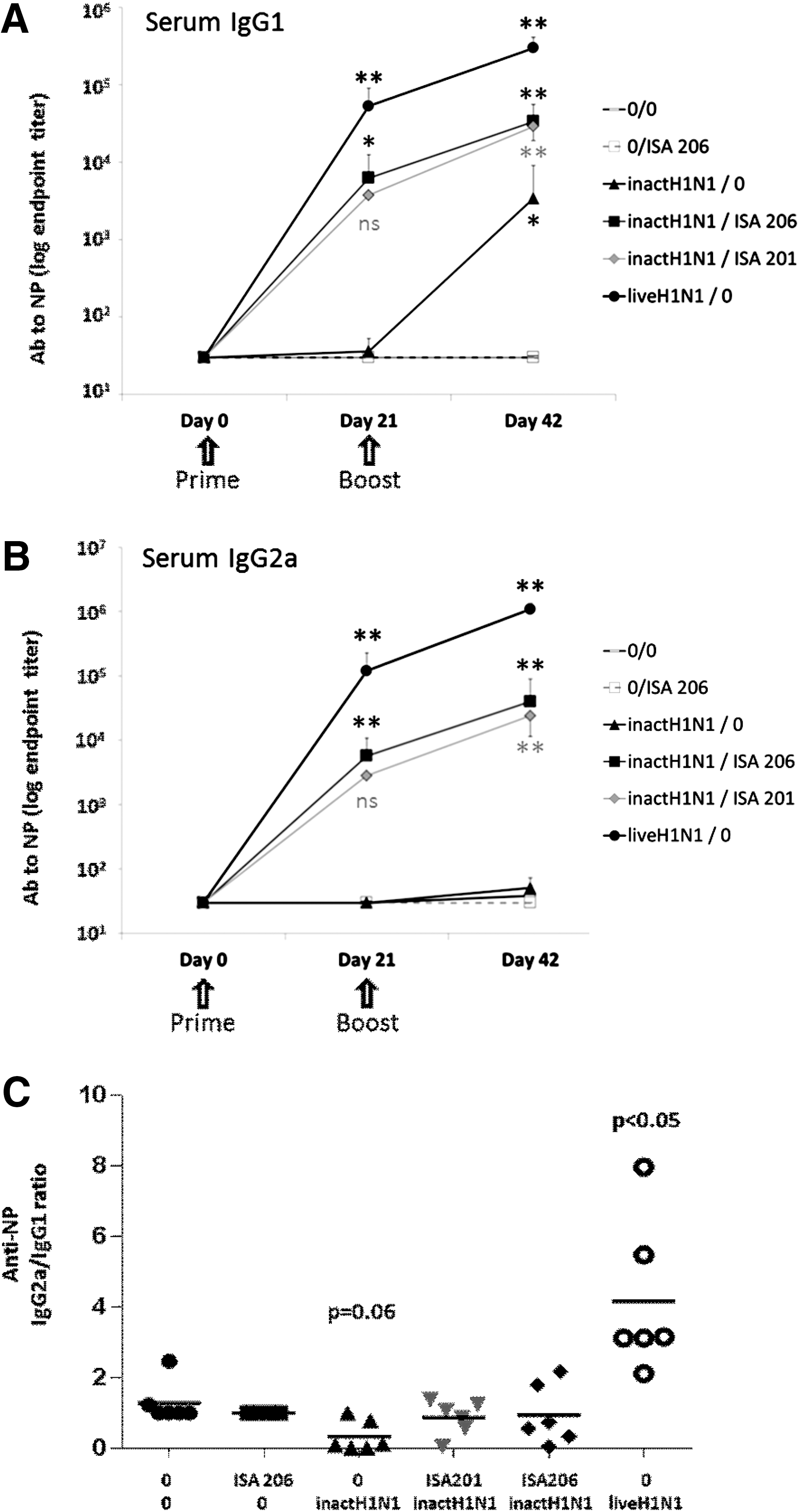
ISA 206 and ISA 201 adjuvants induce similar anti-influenza serum IgG1 and IgG2a levels. (
To assess the T cellular response induced by vaccination with adjuvanted vaccines, splenocytes were isolated at day 42 and CFSE stained. They were then restimulated in vitro with inactivated or live influenza virus, and the proliferation (CFSElow cells) of CD4 and CD8 T lymphocytes after restimulation was measured. In the three control groups (0/0, 0/ISA 206, InactH1N1/0), no specific proliferation was observed, although a high background was detected in the group vaccinated with adjuvant alone (Fig. 2A–C). In the group InactH1N1/ISA206, a low specific proliferation of CD4 and CD8 T cells was detected, whereas a significantly higher proliferation of both CD4 and CD8 T cells was observed using ISA 201 instead of ISA 206 adjuvant. The H1N1 infected group presents the highest CD4 and CD8 T cells proliferations. Interestingly, the in vitro restimulation using live H1N1 revealed more specific CD8 T cells, probably because of the induction, in infected mice, of CD8 T cells directed against determinants only present during live influenza infection but not in the inactivated viral particles, such as nonstructural proteins. These results show that the use of the ISA 201 adjuvant in influenza vaccine can induce specific CD8 and CD4 responses against influenza virus.
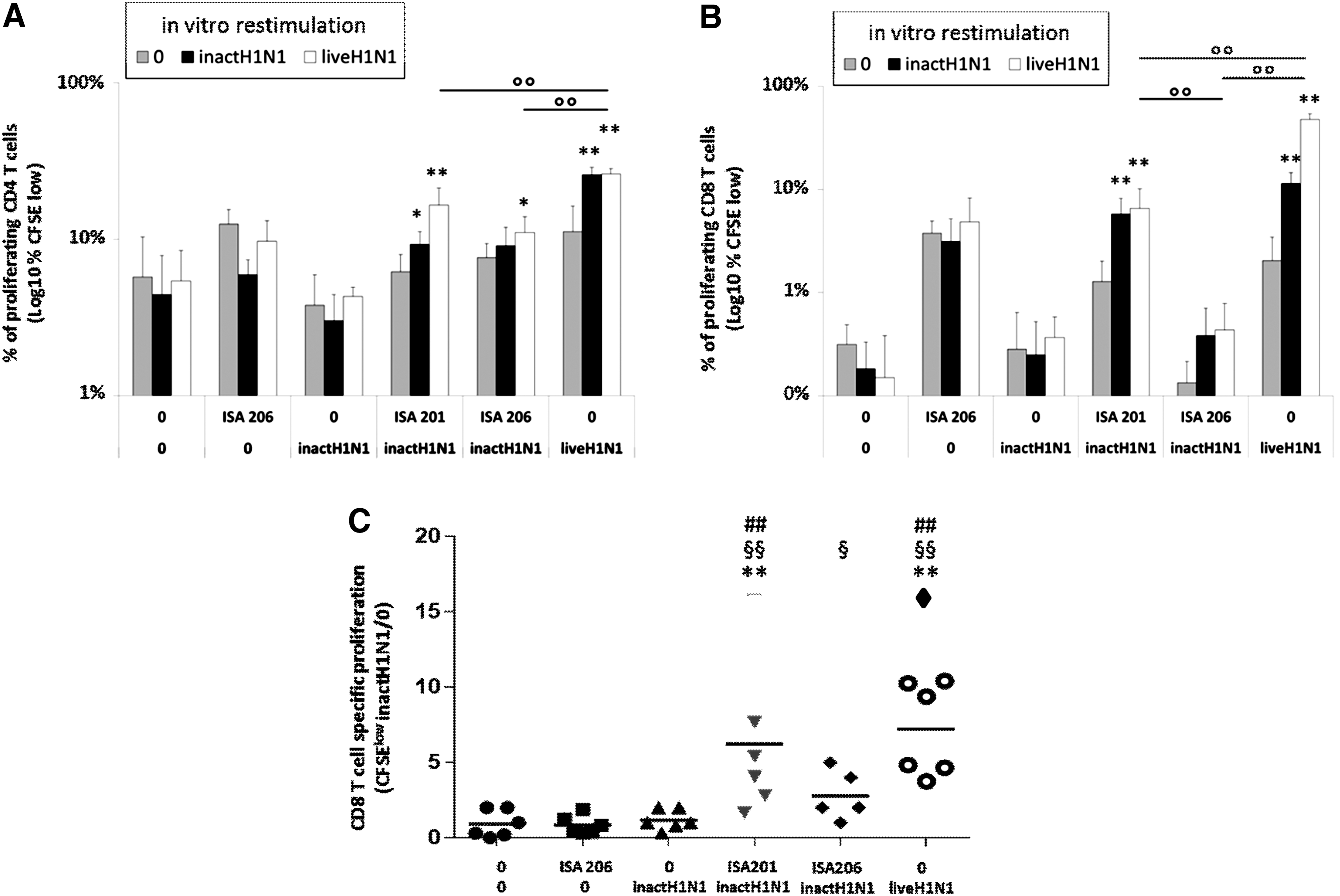
Adjuvant ISA 201 induces strong CD4 and CD8 T cell responses. (
Altogether, these results show that the ISA 201 adjuvant induces a strong humoral response with a mixed Th1/Th2 response, and an antigen-specific T CD4 and T CD8 cellular response. Compared with ISA 201, the reference adjuvant ISA 206 induces a similar humoral response but low CD4 and CD8T cell responses. It should be noted that vaccination with the nonadjuvanted antigen induces a very weak Th2 biased humoral response and no cellular response.
Swine vaccine trial: safety
Based on the previous results obtained in mice, the efficacy and safety profiles of a vaccine against swine influenza based on ISA 201 were assessed in swine. Two-month-old pigs were vaccinated at day 0 and day 21 with ISA 201 adjuvanted or nonadjuvanted inactivated swine H1N1 vaccines. A group of unvaccinated animals was included in the trial as a negative control.
The safety profile of the adjuvanted vaccine was assessed by body temperature measurement after first vaccination and boost vaccination, and observation of local reactions during the trial and after dissection of the injection site at slaughter (day 56). No body temperature increase >1°C was observed after the first or second injection in any group. No local reactions at the injection site were observed during the trial. At slaughter, no reactions were observed in animals vaccinated with the nonadjuvanted vaccine. In the group vaccinated with antigen plus ISA 201, small granulomas (with a diameter from 1 mm to 1 cm) were observed in 50% of the pigs (5/10). This type of reaction is acceptable, and these results show that the safety profile of ISA 201 in this model is compatible with swine vaccination.
Swine vaccine trial: antibody response
Specific antibody titers against SIV were measured for each animal at day 0, day 21, day 42 (before challenge), and day 56 (for infected and uninfected animals). The ISA 201 adjuvanted vaccine induced positive titers from day 21 to day 56. At day 56, vaccinated uninfected animals reached high titers that were similar to serological results obtained in blood samples of infected animals (Fig. 3A). Importantly, similar results were obtained by HI experiments (Fig. 3B), in agreement with the presence of neutralizing antibody in the serum of vaccinated pigs.
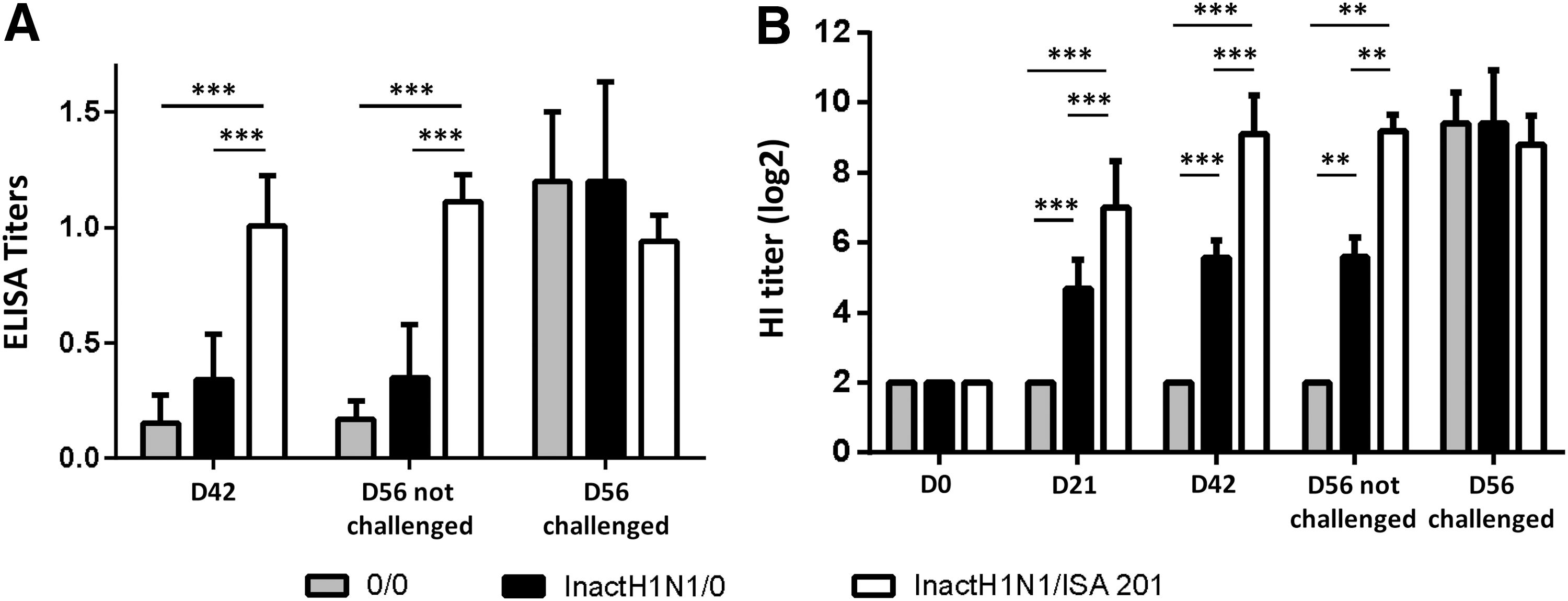
Adjuvant ISA 201 induces a strong humoral immune response in swine. (
Swine vaccine trial: protection to challenge
As a first proof of concept of ISA201 adjuvant efficacy, it was tested in a homologous field infection, using a very demanding virus dose (108 EID50), which would ascertain that the nonadjuvanted vaccine would not be totally protective and thus would allow any improvement of the vaccine given by ISA201 adjuvant to be monitored. In each group, five pigs were submitted at day 42 to an intranasal infectious challenge with SIV. Body temperature after infection was measured for 14 days post-challenge. In these demanding conditions, unvaccinated animals presented two peaks of hyperthermia at days 1/2 and days 5/6. Interestingly, unadjuvanted vaccine protected the animals from the first signs of infections (hyperthermia from day 0 to day 4 post-infection) but could not avoid some delayed hyperthermia (Fig. 4A). At slaughter at day 56, the lung lesions of infected pigs were scored using the method described by Halbur et al. (13). According to the presence of lung lesions 14 days post-challenge (Fig. 4B) and to polymerase chain reaction detection of circovirus type 2 and Haemophilus parasuis (data not shown) on randomly tested animals, the second peak of fever was most probably due to secondary opportunistic infections. Interestingly, despite the very high challenge dose, animals vaccinated with the adjuvanted vaccine presented reduced signs of secondary infections (Fig. 4A and B).
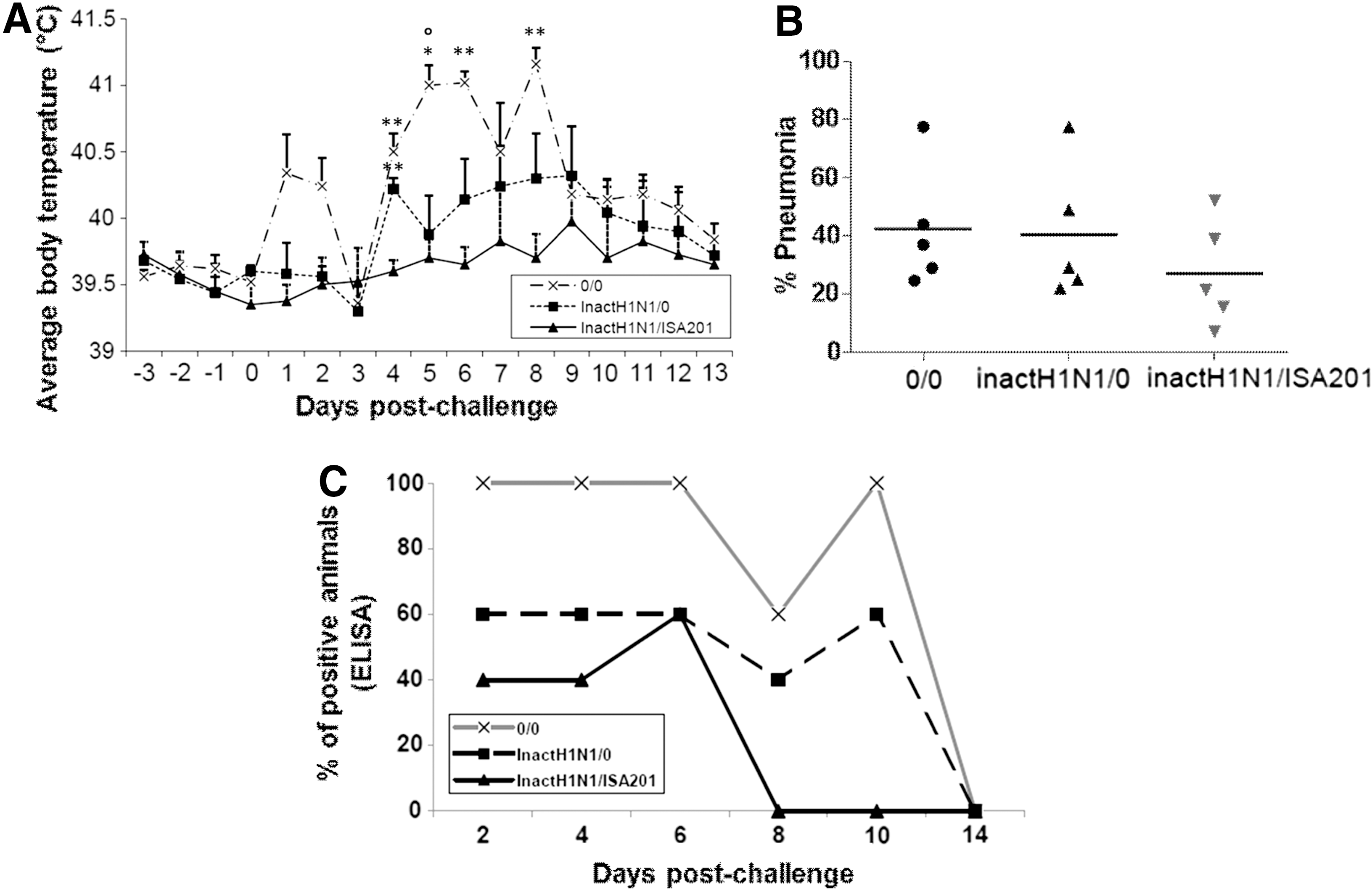
InactH1N1 + adjuvant ISA 201 decreases influenza symptoms after challenge in swine. Five pigs per group were challenged with A/swine/California/07/09 (H1N1) live virus at day 42 after vaccination with inactH1N1 ± ISA 201. Clinical symptoms were recorded for 14 days after challenge (until day 56). (
Viral shedding in nasal swabs was assessed at day 2, day 4, day 6, day 8, day 10, and day 14 post-infection. Surprisingly, probably because of the very high virus dose used for challenge, all the unvaccinated pigs excreted virus for a very long period of time, until day 10 post-challenge. Similarly, 60% of the vaccinated animals with nonadjuvanted vaccine excreted virus as late as at 10 days post challenge, whereas the animals vaccinated with adjuvanted vaccine did not excrete virus after day 6 post-infection (Fig. 4C). Thus, ISA 201–based vaccines allowed the reduction of the number of virus excreting pigs and the duration of viral shedding compared to unvaccinated pigs and pigs vaccinated with the nonadjuvanted vaccine. These results show that in very challenging conditions, ISA 201 increases the protection conferred to animals by SIV inactivated vaccines.
Discussion
Swine influenza vaccination is the most effective way to control SIV in the field. However, standard inactivated vaccines against SIV mostly induce humoral immunity and do not protect against new field strains that have been submitted to antigenic drift or shift. Diverse approaches have been developed to enhance cross-protection conferred by influenza vaccines, such as the development of DNA, subunit or vectored vaccines, or the production of autogenous vaccines that are adapted to the strain circulating into a specific farming unit (5). Another option would be to improve inactivated whole virus vaccines that are still mainly used in the field, in order to enhance their efficacy, by optimization of the virus inactivation process (4) or by improvement of the vaccine adjuvant properties.
Classical veterinary adjuvants mostly orient the vaccinal immune response toward a Th2 humoral immune response. However, induction of cell-mediated immunity and in particular cytotoxic CD8 T lymphocytes response (CTL) against conserved influenza epitopes could enhance heterologous protection against influenza strains (24). The development of adjuvants able to orient the immune response of inactivated vaccines toward a mixed Th1/Th2 immune response and the production of CTL response directed against internal conserved antigens would be highly beneficial to improve existing SIV vaccines (10,15). This study shows that the W/O/W adjuvant Montanide™ ISA 201 VG allows the induction in mice of a higher CD8 T cell response when compared with adjuvants that are classically used in the field.
As expected, experiments in mice have shown that vaccination with nonadjuvanted inactivated whole virus induces a weak response that is strongly biased toward a Th2 humoral response. On the other side, infection with the live pathogenic virus induces a strong humoral and cellular response with a strong bias toward a Th1 response. Vaccination with adjuvanted vaccines induces a mixed Th1/Th2 response, including a strong antigen-specific humoral response and CD4 T cell response. However, mice vaccinated with ISA 201 adjuvanted vaccine developed an antigen-specific CD8 T cell response significantly stronger than mice vaccinated with ISA 206 adjuvant.
Furthermore, this study shows that this adjuvant is well tolerated in swine when combined in SIV vaccines. The use of safe adjuvants is crucial for pig farming, as pigs are sensitive animals in which local reaction and hyperthermia after vaccination are frequent and cause losses to swine production. ISA 201 is therefore an interesting candidate to improve CTL response against inactivated vaccines in swine.
Using field conditions, with a very demanding virus dose (8.3 log10TCID50), this study shows that the ISA 201 adjuvanted SIV vaccine induces homologous protection against H1N1 influenza challenge in swine, and that it allows a better protection against secondary infections than the nonadjuvanted vaccine. Indeed, when the ISA 201 adjuvanted vaccine is compared with nonadjuvanted SIV vaccine or to unvaccinated animals, hyperthermia intensity and duration are reduced in animals submitted to a viral influenza challenge. Accordingly, lung lesions and viral shedding in nasal swabs after challenge are also reduced, without being totally abrogated, in agreement with others using similar infection doses (7.5 log10TCID50) of a virus strain (A/Netherlands/602/2009/H1N1) closely related to the H1N1 virus A/swine/California/07/09 used in this study (19). It is interesting to note that the two opportunistic infections emerging in absence of the adjuvanted vaccine were two components of the porcine respiratory disease complex (PRDC) (6), namely circovirus type 2 and H. parasuis. These data highlight the possibility that the use of efficient vaccines directed against only two or three of the pathogens participating to PRDC would alleviate the economic burden of PRDC. Indeed, any management procedure that will reduce the chance of combined infections would be beneficial, since in pigs, >88% of cases of respiratory disease are caused by coinfection (6).
As it has been shown that ISA 201 enhances cytotoxic immune response against conserved antigens (NP) in mice, it is expected that ISA 201 adjuvant may have the potential to enhance cross-protection conferred by SIV inactivated vaccines in swine. Since antigenic drift and reassortment is not frequent in SIV strains, the goal of a true “universal vaccine” protecting against the few strains circulating in pig farms might be more easily achievable in swine than in humans, using, as suggested by others, whole inactivated viral particles associated to the right adjuvant (10). To test this possibility, a heterologous challenge study in swine vaccinated using inactivated influenza plus ISA 201 adjuvant should be performed with other SIV strains.
This study also indicates that adjuvants should be chosen with specific care for each new vaccine development project, as they are not only enhancers of the strength of the immune response to the vaccine, but can also influence the orientation of the immune response. New developments in the field of adjuvants should improve existing and new vaccines in the future.
Footnotes
Acknowledgments
We thank Nathalie Lejal and Any Slama-Schwok for the grateful gift of purified PR8/34 nucleoprotein (NP). This work was funded by the Institut National de la Recherche Agronomique (INRA) and by SEPPIC.
Author Disclosure Statement
Seppic commercializes the Montanide range of vaccine adjuvants. Elodie Goncalves, Sebastien Deville, and Juliette Ben Arous are employees of Seppic. No competing financial interests exist for the remaining authors.