
Abstract
Poxviruses have evolved numerous mechanisms to avoid the immune response of the infected host, and many of these mechanisms have not been fully described. Here, we studied the transcriptional response of innate immune genes in BALB/c and C57BL/6 peritoneal macrophages following infection with the Moscow strain of ectromelia virus (ECTV-Mos) with the aim of delineating innate immune genes that contribute to the difference between susceptibility and resistance to lethal infection. We show a generalized downregulation of many genes in four categories (toll-like receptor signaling, NOD-like receptor signaling, RIG-I-like receptor signaling, and type I interferon signaling) of antiviral innate immune receptors, downstream signaling pathways, and responsive components. Two important observations were made. First, 14 innate antiviral genes were differentially expressed with fold change upregulation of two and above occurring in C57BL/6 mice, known to be resistant to ECTV-Mos infection, whereas the same genes were downregulated in BALB/c mice with fold change of two and below. Second, the cathepsin group of genes was downregulated in both strains of mice but with profound fold changes of 17, 38, and 62 downregulation for CtsL, CtsB, and CtsS, respectively, in C57BL/6 mice. We show that a poxvirus profoundly downregulates both the mRNA and protein expression of these three cathepsins and this change appears to support virus replication. Based on these data we propose that the variations in gene expression observed may contribute to the difference in resistance/susceptibility between BALB/c and C57BL/6 mice to lethal infection by ECTV-Mos.
Introduction
I
The murine poxvirus, ectromelia virus (ECTV), is a valuable model to study the influence of viruses on host innate responses because of the availability of mouse strains that are susceptible or not to lethal infection with ECTV. Infection of B10.A(2R) or B10.G mice with ECTV leads to two different outcomes. In B10.G mice, ECTV infection usually produces a fatal disease, whereas B10.A(2R) mice are said to be resistant to lethal infection (28). In the latter case, the authors explained that the H-2 genes controlled resistance to ECTV by influencing the effectiveness of the cellular immune response (28). The explanation, however, does not accommodate the contribution of innate immune mechanisms, which are usually the first to respond to infection by a virus. Furthermore, the hypothesis that an inappropriate cytokine response may lead to increased susceptibility to poxvirus infection has been postulated in many studies and has been supported by some experimental data (8).
At the molecular level, several mechanisms have explained how vaccinia virus (VACV) or MPXV (2) has been successful in causing disease. In general, the immune response to infection with ECTV is fairly well known (5); however, only a few studies (6,25,46) describing the primary gene transcription signatures immediately following infection are available, let alone comparisons of these signatures between distinct mouse strains with different levels of susceptibility to infection. Therefore, a global transcriptional profile of innate immunity genes following infection with a poxvirus is not currently available. As a result, few signature innate genes that orchestrate the outcome of infection by poxviruses have been described during the responses of various target cells. Such knowledge would help to precisely define the molecular pathogenesis of poxviruses. Preferably, comparative studies between susceptible and nonsusceptible mouse strains should address this knowledge gap and focus on the influence of poxvirus infection on the transcriptional innate gene profile in frontline cells, such as dendritic cells (DCs) or macrophages (MΦ). The molecular pathogenesis pathways will show innate immunity initiating factors and the overall interaction of the innate immune components that are affected by poxvirus infection. A comprehensive understanding of global gene transcription during poxvirus infection will provide the basis to design better vectors, therapeutics, and/or vaccines against poxviruses.
It is widely appreciated that macrophages are involved in the crucial early events of infection with viruses (18). Therefore, they constitute an appropriate target to examine early innate molecular immunity during viral infection. We hypothesized that early changes in innate gene transcription and protein expression determine the direction of an immune response. Therefore, we investigated the global expression of mRNAs for genes involved in the innate antiviral responses to lethal infection of a susceptible or nonsusceptible host by a poxvirus. Consequently, a comprehensive overview of the transcriptional profiles of innate immune genes was determined in peritoneal macrophages of BALB/c and C57BL/6 mice infected with ECTV-Mos. The results show differential mRNA expression in at least 14 genes and primarily a downregulation of many genes, indicating that infection with poxvirus induces an initial arrest of both sensing and downstream signaling pathways. The suppression of mRNA expression was much more evident in the BALB/c mice, which have widely been reported to be highly susceptible to ECTV lethal infection. For the first time, we show that ECTV profoundly affects both the mRNA and protein expression of at least three cathepsins: CtsB, L, and S. The implication of this finding on the replication of ECTV-Mos is currently under investigation. In addition, we did not observe an effect on Ifnα1 and Ifnβ1 mRNA expression. Moreover, we could not see a change in protein expression that coincided with the mRNA expression pattern. With the inhibition of a significant number of important innate immunity genes, as shown in this report, the susceptible mouse strain (BALB/c) is unlikely to mount a protective innate immune response that can support a protective cellular immune response. Consequently, the infection is fatal.
Materials and Methods
Animals
Male BALB/c and C57BL/6 mice were obtained from the Maria Skłodowska-Curie Memorial Cancer Centre and Institute of Oncology in Warsaw, Poland. The breeding colony is a certified breeder and is periodically tested for ECTV and is free from ECTV. The mice were used in the experiments at an age of 6–10 weeks and after a 10-day acclimatization following purchase. The 3rd Local Ethics Committee for Animal Research at the Warsaw University of Life Sciences-SGGW (Permit nr 41/2010) approved the experimental protocols. Unless otherwise stated, experiments described in this study were repeated three times with at least five mice per treatment group.
Cells and virus
The peritoneal macrophages were elicited by incomplete thioglycolate treatment, as described in Zhang et al. (49). On the second day after incomplete thioglycolate administration, the mice were intraperitoneally infected with ectromelia virus Moscow strain (ECTV-Mos, ATCC VR-1374, USA) at a dose of 1 × 103 pfu in 100 μL of phosphate-buffered saline (PBS). The control mice (uninfected) were similarly injected with PBS. On the fourth day after administration of incomplete thioglycolate, peritoneal macrophages (pMΦ) were collected as described in Zhang et al. (49) and washed at least three times in PBS. Following cell enumeration, the cells were sorted through magnetic labeling using the Miltenyi Biotech MACS System (Germany). Briefly, the cells were washed once in staining buffer (PBS, 3% fetal calf serum [FCS], and 2 mM EDTA) and suspended in 5 mL of the same buffer. The F4/80 macrophage marker was used to label macrophages for sorting. One microgram of rat anti-mouse F4/80: low endotoxin antibody (Clone Cl:A3-1, Bio-Rad, formerly Serotec) was added to 1 × 106 cells and incubated for 20 min on ice. Following three washes in staining buffer, anti-rat IgG microbeads (Miltenyi Biotec, Germany) were mixed with the F4/80-labeled macrophages, incubated for another 20 min on ice, washed three times, and finally separated on magnetic columns. At least 97% purity of the sorted cells was attained (Fig. 1). The L929 (ATCC-CCL1) and Vero cells were maintained in Dulbecco's minimum essential medium supplemented with 10% FCS and penicillin–streptomycin. Confluent monolayers were infected with ECTV-Mos at a multiplicity of infection (MOI) of 1 with adsorption for 1 h, and the cells were further incubated at 37°C in 5% CO2 for 48 h. For virus titration, Vero cells were inoculated with pMΦ lysates.
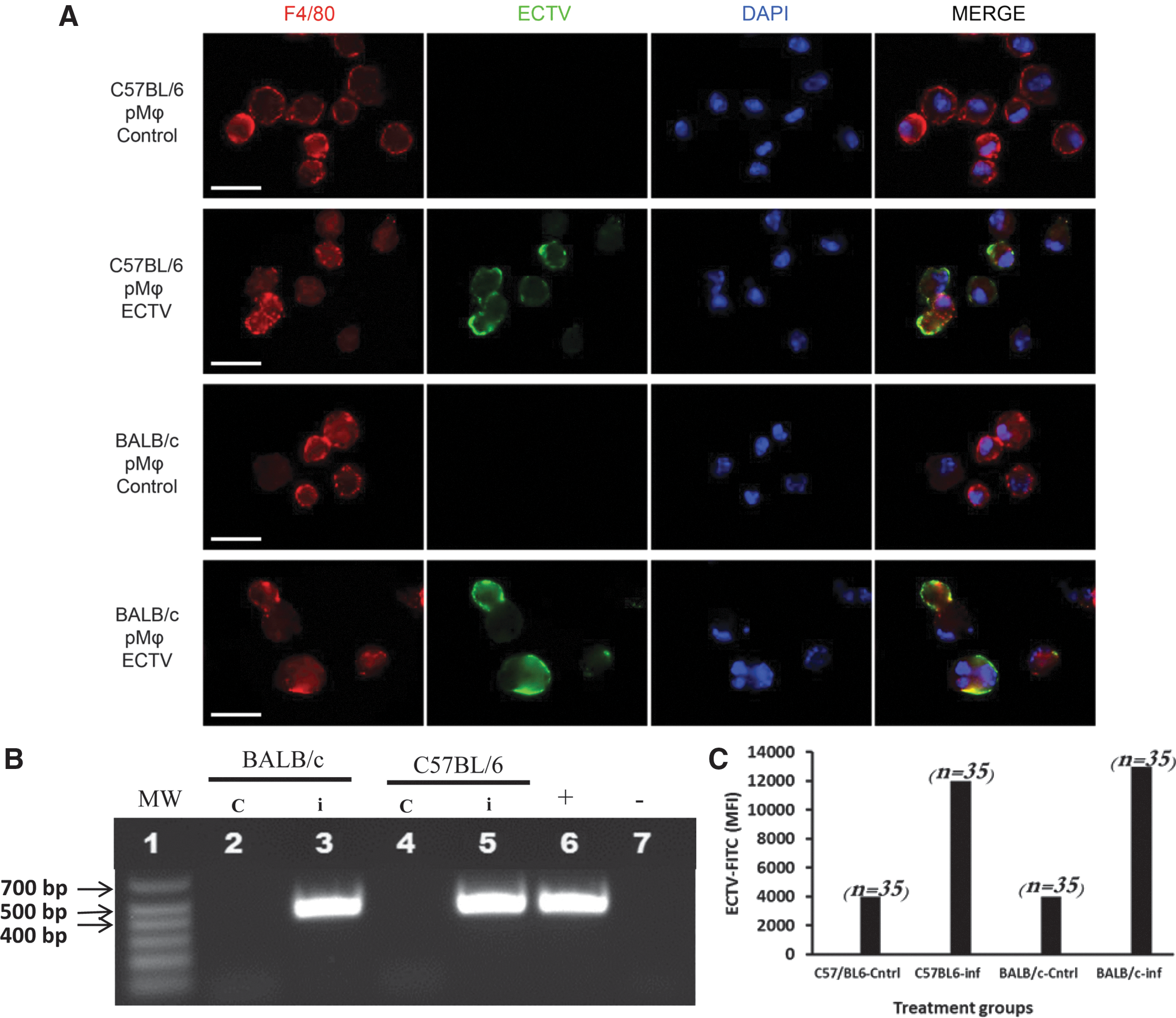
Peritoneal macrophages infected with ECTV-Mos in vivo.
RNA isolation and PCR
RNA was isolated from 1 × 107 sorted pMΦ using the Qiagen RNAeasy Mini Kit (Qiagen, USA), as recommended by the manufacturer. In addition, an on-column DNase I reaction step was included. Briefly, after cell lysis and absorption of the RNA on columns, 1,500 Kunitz units of DNase I were added, and the columns were incubated at room temperature for 15 min followed by two washes. Purified RNA was then eluted into RNase-free tubes with RNase-free H2O. The RNA concentration was measured using the Take-3 system on an Epoch BioTek spectrophotometer and quantified using Gen5 software (BioTek Instruments, Inc.). Messenger RNA transcript profiling was performed using RT2 Profiler PCR array plates purchased from Qiagen. First, cDNA was prepared from 500 ng of RNA using the RT2 First-Strand Kit, as recommended by the manufacturer (Qiagen). Complementary DNA was then added to the RT2 SYBR Green Master Mix and aliquoted into the RT2 Profiler PCR array plates at 25 μL/well. Amplification was conducted in an ABI 7500 thermocycler (Life Technologies) at 95°C for 10 min, 40 cycles of 95°C for 15 sec, and 60°C for 1 min. The fluorescence data were collected at the 60°C/1-min cycle. Each PCR array plate contained lyophilized primers for 84 genes associated with the innate antiviral immune response in mice grouped into four categories: toll-like receptor signaling, NOD-like receptor signaling, RIG-I-like receptor signaling and type-I interferon signaling. Five housekeeping genes (Actb, B2m, Gapdh, Gusb, and Hsp90ab1) were run concurrently to allow selection of the most stable mRNA transcript for normalizing mRNA expression.
Data quality control
The data were quality controlled in three ways: (1) PCR array reproducibility, (2) reverse transcriptase control, and (3) genomic DNA contamination (GDC) control. The criteria for passing the PCR array reproducibility step were an average positive PCR control (PPC) Ct value of 20 ± 2 and no two arrays having an average PPC Ct value >2 away from one another. The criterion for passing the reverse transcription control (RTC) step was a ΔCt (AVG RTC − AVG PPC) ≤5, which meant there was no inhibition of the RT reactions. The criterion for passing the GDC step was Ct(GDC) ≥35, which meant that GDC was absent or too low to affect the gene expression profiling results. To facilitate the elimination of GDC, DNA digestion with DNAse I was applied during RNA isolation. In addition, a genomic DNA elimination step was included before reverse transcription. The data reported here fulfilled all of the criteria specified above.
Data analysis, interpretation, and statistics
The amplification data were acquired through SDS software (Applied Biosystems). The data analysis was performed at the Qiagen Data Analysis Center (
Conventional PCR and immunofluorescence
To confirm infection of pMΦ, a fragment of EVM012 (zinc finger-like protein) of ECTV was amplified using forward primer 1: ACAATGGTGCAAGATATCCATTATCGAGCAAGCATTGCTAAAACTCG and reverse primer 2: AGCTGATAGTGGTACCTAGAGACGGCCAATTAGAAGTTCGT. Primers were designed using SnapGene software (GSL Biotech LLC). Samples from infected and noninfected cells, including positive and negative controls, were amplified with the CloneAmp HiFi PCR Premix (Clontech, Japan) using the following cycling parameters: denaturation at 94°C for 5 min, followed by 30 cycles of denaturation at 94°C for 1 min, annealing at 55°C for 1 min, and elongation at 72°C for 3 min; and a final elongation at 72°C for 5 min. The reactions were then maintained at 4°C pending further analysis. Standard procedures were followed for electrophoresis, and the gels were documented in VersaDoc using Quantity One software (Bio-Rad).
Immunofluorescence was performed to confirm the localization of viral antigen in pMΦ. Approximately 106 cells/mL were first stained with F4/80 antibody (BioLegend) for 20 min on ice followed by an anti-rat IgG-Texas Red antibody for another 20 min. The cells were then permeabilized with the BD Cytofix/Cytoperm solution, washed in BD Perm/Wash buffer (BD Biosciences), and finally labeled with a polyclonal FITC-conjugated anti-ECTV-Mos antibody for 1 h on ice. Then, the cells were fixed in 4% paraformaldehyde for 20 min at room temperature, washed twice, and resuspended in PBS. Fifty microliters of the stained cells were pipetted onto silanated glass slides and allowed to dry at 37°C. Later, the cells were hydrated in PBS and dried in air, followed by mounting in 4′6′-diamidino-2-phenylin-dole (DAPI; Vector Labs, Burlingame, CA) for nuclear counterstaining. Image documentation and analysis were performed on an Olympus BX60 fluorescence microscope attached to a Color View III cooled CCD camera and Cell^F software (Olympus, Hamburg, Germany). Mean fluorescence intensity (MFI) was measured on photomicrographs with ImageJ (32). The MFI = integrated density − (area of selected cell × mean fluorescence of background readings).
Protein expression
Depending on the availability of the appropriate antibody reagents, protein expression was monitored by Western blot or a cytometric bead array (CBA; BD Biosciences) assay, as follows.
Western blotting analysis
Peritoneal macrophages, L929, or Vero cells prepared as described in the “Cells and virus” section were first washed with ice-cold PBS and lysed in ice-cold radioimmunoprecipitation assay buffer (RIPA buffer; Thermo Scientific). The lysates were clarified by centrifugation for 15 min at 4°C. Quantitation of the protein content in lysates was performed by the bicinchoninic acid assay (Thermo Scientific) and spectrophotometry on an Epoch BioTek spectrophotometer. Sample preparation for electrophoresis and protein transfer was performed according to the Bolt® Blot Module System (Life Technologies). Briefly, cell lysates containing 20 μg of protein were incubated with lithium dodecyl sulfate sample buffer and sample reducing agent for 10 min at 70°C. Subsequently, the samples and protein markers were electrophoresed on a 10% polyacrylamide Bis-Tris Plus gel with MES running buffer and transferred to polyvinylidene fluoride membranes. The transferred proteins were sequentially probed with the primary antibodies listed in Table 1. Secondary anti-goat, anti-mouse, and anti-rabbit antibodies conjugated to horseradish peroxidase (Santa Cruz Biotechnology) were added, and chemiluminescence or color was developed with the SuperSignal West Pico chemiluminescent substrate (Thermo Scientific) or Pierce 1-Step Ultra TMB blotting solution (Thermo Scientific), respectively. The protein bands were visualized by autoradiography or VersaDoc. GAPDH was used as a protein loading control.
Identification details not provided by the manufacturer.
Clone.
Manufacturer's antibody identification.
Cytometric bead array
CCL3, CCL4, CCL5, CXCL9, and total STAT1 were captured by CBA (BD Bioscience, San Jose, CA) as outlined in the manufacturer's manual. Protein levels were then quantified by flow cytometry (BD Fortessa; BD Bioscience, San Jose, CA). Data were analyzed with FCAP array software (v3.0; BD Bioscience, San Jose, CA).
siRNA knockdown of cathepsins B and L
RAW264.7 cells were grown in RPMI-1640 GlutaMAX supplemented with 10% FBS and seeded at 3 × 105 per well in six-well plates. After overnight culture, cell confluent monolayers were treated with siRNA against cathepsin B or L. Confirmation of gene knockdown was assessed by measuring protein content in lysates by Western blot (see Supplementary Fig. S1; Supplementary Data are available online at
Results
Initial reaction of the innate immune system on infection with a virus may decide the fate of the host. BALB/c mice are susceptible to infection, while C57BL/6 are not. We sought to look for differences between these two mouse strains in antiviral innate gene expression profiles following infection. Peritoneal macrophages were derived from both BALB/c and C57BL/6 control mice (noninfected) and ECTV-Mos-infected mice. We used the peritoneal route to derived equal numbers as well as uniformly infected cells. Because this route of infection may kill mice from both strains, cells were collected at 48 h postinfection. This time point was chosen because preliminary data showed no or minor differences in fold transcriptional changes of the genes analyzed at earlier time points. Moreover, a similar observation of lack of transcriptional changes at earlier time points was reported by Sigal and colleagues (47) in relation to type 1 IFN and ISG transcripts. Therefore, we show results indicating innate response during increased virus load in infected cells. It is worthy of note that another study using a highly sophisticated technique has reported analysis of VACV and host cell transcriptomes at earlier time points (48). In addition, we used incomplete thioglycolate with omission of agar and methylene blue to avoid activation of macrophages (23). RNA was isolated and real-time PCR performed with RT2 profiler PCR array assays. Protein expression was measured by Western blot and CBA.
Peritoneal macrophages infected with ECTV-Mos in vivo
First, we sought to confirm that the cells were indeed infected with the virus. Therefore, immunofluorescence was performed on cells from control and infected mice. Figure 1A depicts the staining of control and infected pMΦ with an anti-ECTV antibody, indicating that the majority of the cells were infected at the time of preparation for RT-PCR or protein analysis. This result is similar to that reported in an earlier study by Roberts (29), although those experiments were performed on cultured macrophages. The immunofluorescence evidence was corroborated with conventional PCR amplification of the ECTV-Mos EVM012 gene, where a fragment of ∼439 bp was readily amplified in pMΦ originating from the infected mice of both strains and not in cells from the control animals (Fig. 1B). Although the immunofluorescence showed uniform infection of pMΦ from both strains of mice, there was a difference in the virus titer already at 48 h postinfection (Table 2). To support the claim of uniform cell infection, we confirmed the MFI of stained cells by ImageJ (Fig. 1c). This measurement also showed comparable values between cells from the two mouse strains. This result indicates that although similar level cell infection was apparent, there are differences in cellular response to infection. Following the confirmation of viral antigen in pMΦ from infected mice, we proceeded with the mRNA and protein expression experiments.
Virus titration in ECTV-Mos-infected murine macrophages was performed on Vero cells. Experiments were performed three times with five mice per treatment group. * p ≤ 0.05.
ECTV-Mos, ectromelia virus Moscow strain; SD, standard deviation.
Generalized downregulation of innate genes greater in BALB/c than C57BL/6 mice
Before calculating 2ΔΔCT values, the data were normalized to Hsp90ab1. Hsp90ab1 was chosen for normalization because of its uniform expression across all treatment groups in the experiments. Figure 2 shows the volcano plots and heat maps of the mRNA expression profiles of pMΦ isolated from ECTV-Mos-infected BALB/c and C57BL/6 mice. Supplementary Tables 1 and 2 are essential in understanding the heat maps. The volcano plot combines a p-value statistical test with the fold regulation change, enabling the identification of genes with both large and small expression changes that are statistically significant. In this case, only p-values ≤0.05 were considered. Twofold changes were deemed reflective of genuine changes in transcription and only data with such fold change are shown.
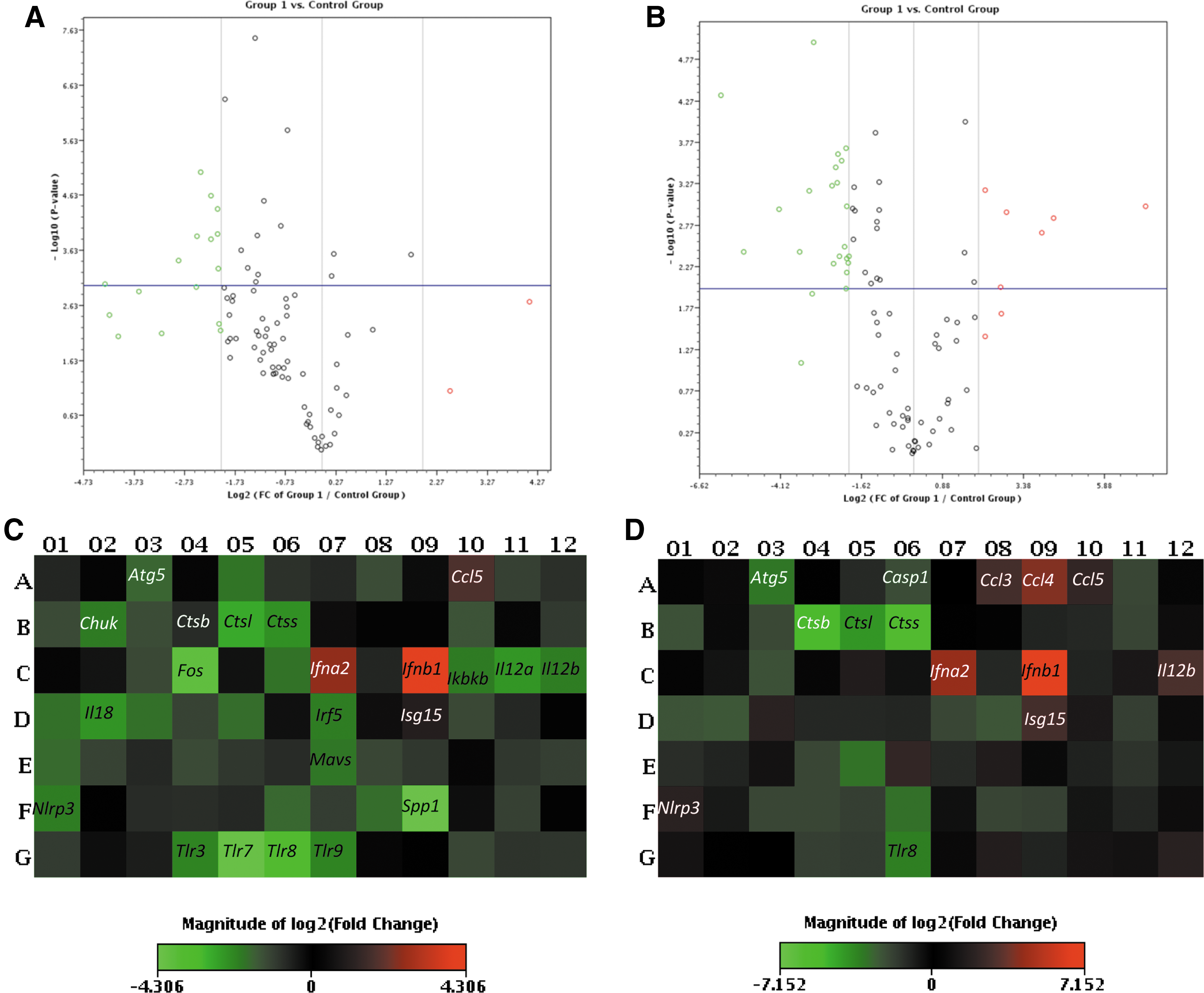
Generalized downregulation of genes in BALB/c than C57BL/mice in response to viral infections; the volcano plots display statistical significance versus the fold change on the y- and x-axes, respectively.
We analyzed a set of genes encoding proteins that regulate innate immune responses during viral infection in four categories: toll-like receptor signaling, NOD-like receptor signaling, RIG-I-like receptor signaling, and type I interferon signaling. In each category, the genes were grouped into the following functional clusters: receptor and chaperone genes, downstream signaling genes, and response genes.
Gene expression in the TLR signaling category
Surprisingly, a profound downregulation of cathepsins B, L, and S was observed in both strains of mice (p ≤ 0.05) (Fig. 3A). These changes were more pronounced in the C57BL/6 mice than in the BALB/c mice. To the best of our knowledge, this observation has not been previously reported during ECTV infection. It may reflect a direct influence of the virus on cathepsin genes as a mechanism of immune escape. In the chaperone and receptor cluster of the TLR signaling category, all viral pathogen-associated molecular pattern (PAMP)-sensing genes were downregulated in both the BALB/c and C57BL/6 mice (Fig. 3A), with the exception of Tlr9, which was upregulated only in C57BL/6 mice. However, this upregulation was not statistically significant (p > 0.05). Figure 3B shows the fold regulation in the downstream signaling cluster of genes. Of the 27 genes analyzed in this group, only Fos, Irf7, Jun, Map3k7, Mapk8, Tbk1, and Traf3 were differentially regulated; they were upregulated in C57BL/6 pMΦ, compared to BALB/c pMΦ in which these genes were downregulated with the exception of Irf7, which was increased in BALB/c pMΦ and was statistically significant (p ≤ 0.05). The remaining genes in this group were downregulated in both mouse strains. Among the 16 genes analyzed in the response gene cluster (Fig. 4), Ccl3, Ccl4, Cxcl9, Cxcl10, Il12a, Il12b, and Il1b were differentially regulated in infected pMΦ. They were upregulated in cells from C57BL/6 mice opposed to downregulation in BALB/c mice. Only Cxcl9 and Cxcl10 were upregulated in cells originating from infected BALB/c mice and downregulated in C57BL/6 mice. Changes in the differentially expressed genes were statistically significant (p ≤ 0.05) in both mouse strains for Ccl3, Il-12a, and Il-12b, whereas Cxcl10 was not (p > 0.05). An important observation from this group was the upregulation of type I IFN genes, Ifna2 and Ifnb1, at 48 h post-ECTV-Mos infection (Fig. 4) in the pMΦ from both mouse strains. However, statistically significant (p ≤ 0.05) expression was noted for both genes in C57BL/6 mice and only expression of Ifnb1 was statistically significant (p = 0.001) in BALB/c mice.
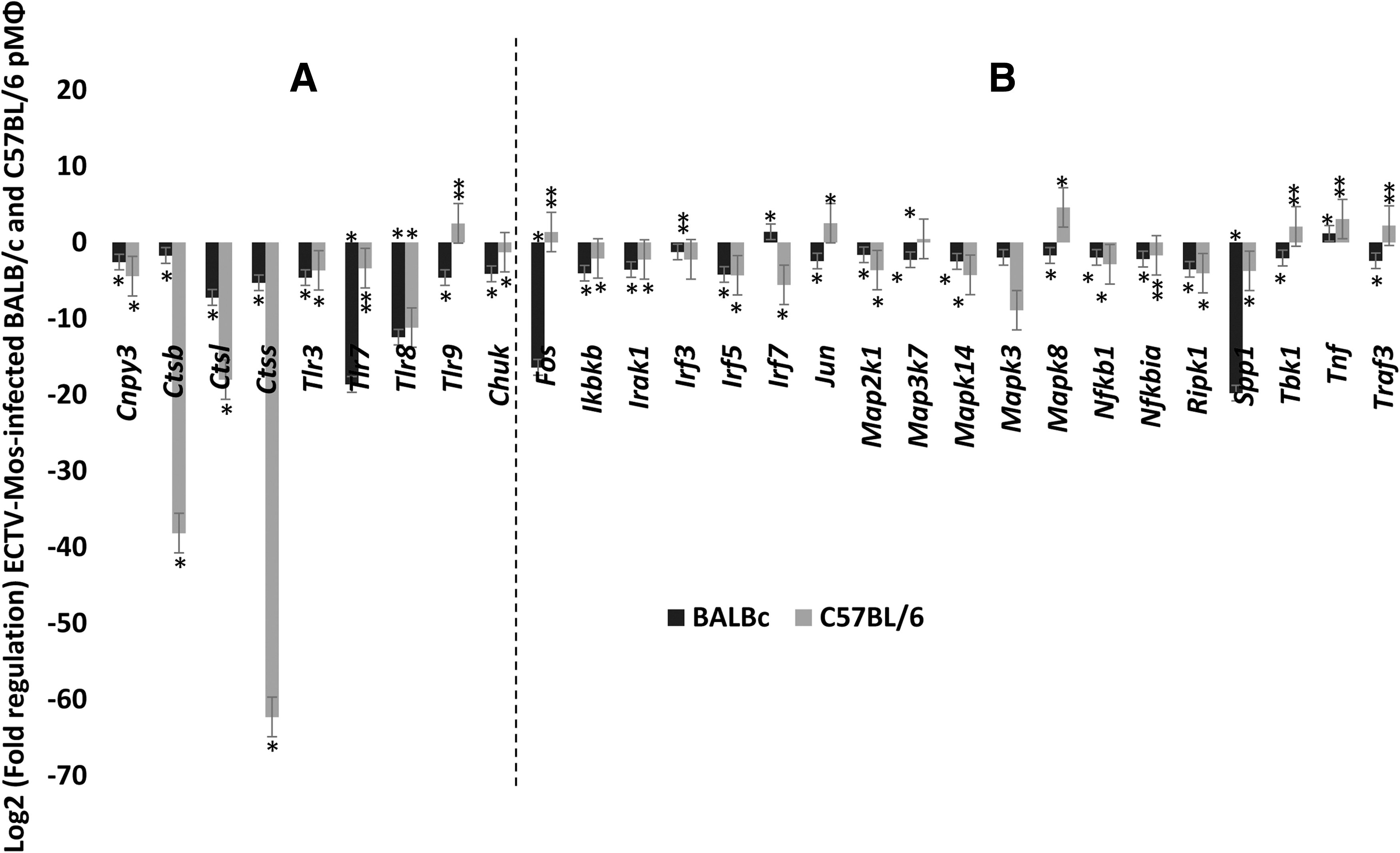
Profound downregulation of cathepsins in C57BL/6 mice following infection with ECTV-Mos.
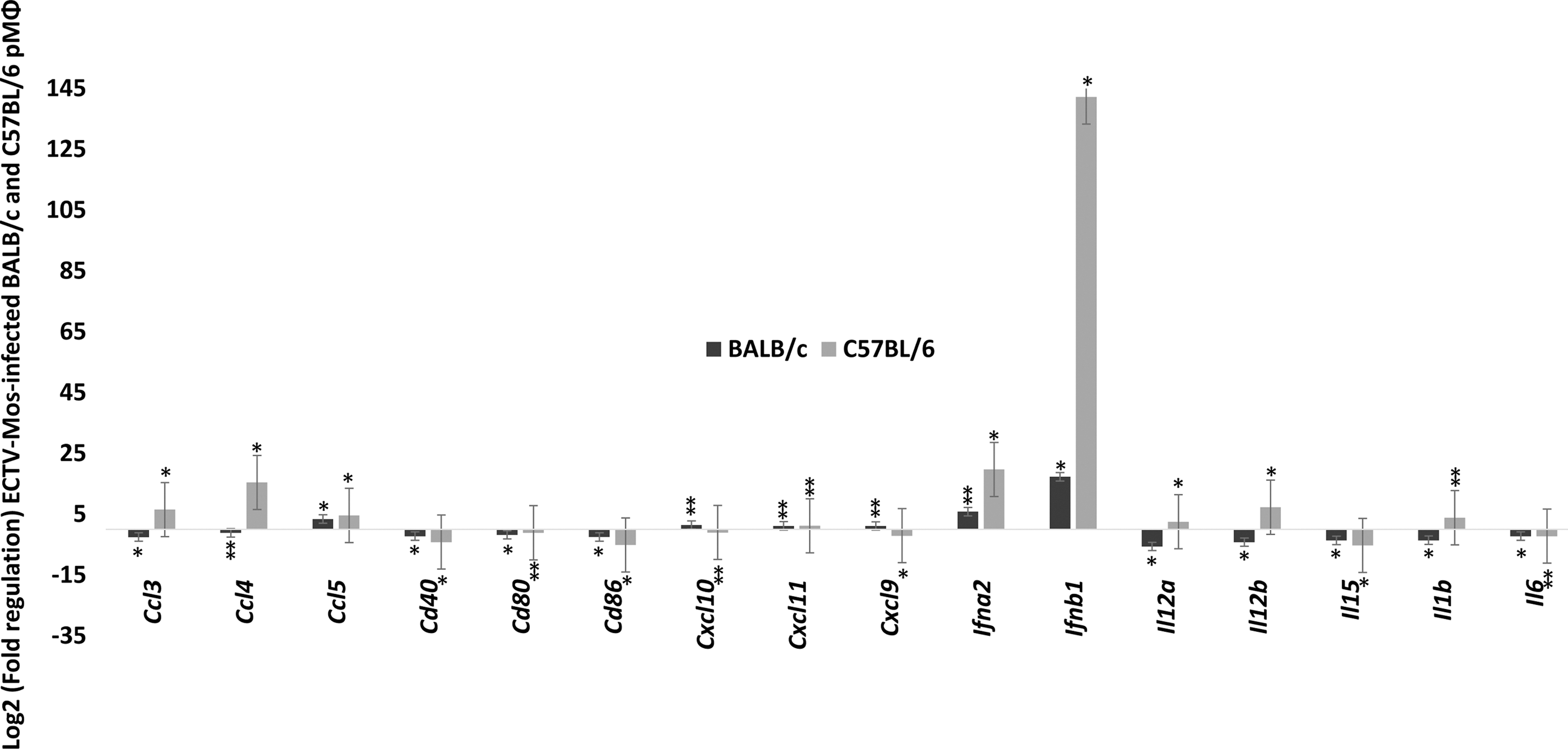
Chemokines and IFNs are highly upregulated in pMΦ on infection with ECTV. Fold regulation of mRNA expression in pMΦ of BALB/c and C57BL/6 mice infected with ECTV-Mos. Response genes in the toll-like receptor signaling group. *p ≤ 0.05, **p ≥ 0.05. Experiments were repeated three times with at least five mice in each treatment group.
Gene expression in the NOD-like receptor signaling category
Two groups of genes were analyzed in the NOD-like receptor signaling category (Fig. 5). Genes such as Nlrp3, Nod2, Mefv, Tbkbp1, and Tank were differentially expressed in the pMΦ from the two strains of mice. The majority of the differentially expressed genes were upregulated in C57BL/6 mice, except for Nod2. Nod2 upregulation was not statistically significant (p = 0.082) in BALB/c mice. The remaining genes were downregulated in both strains.
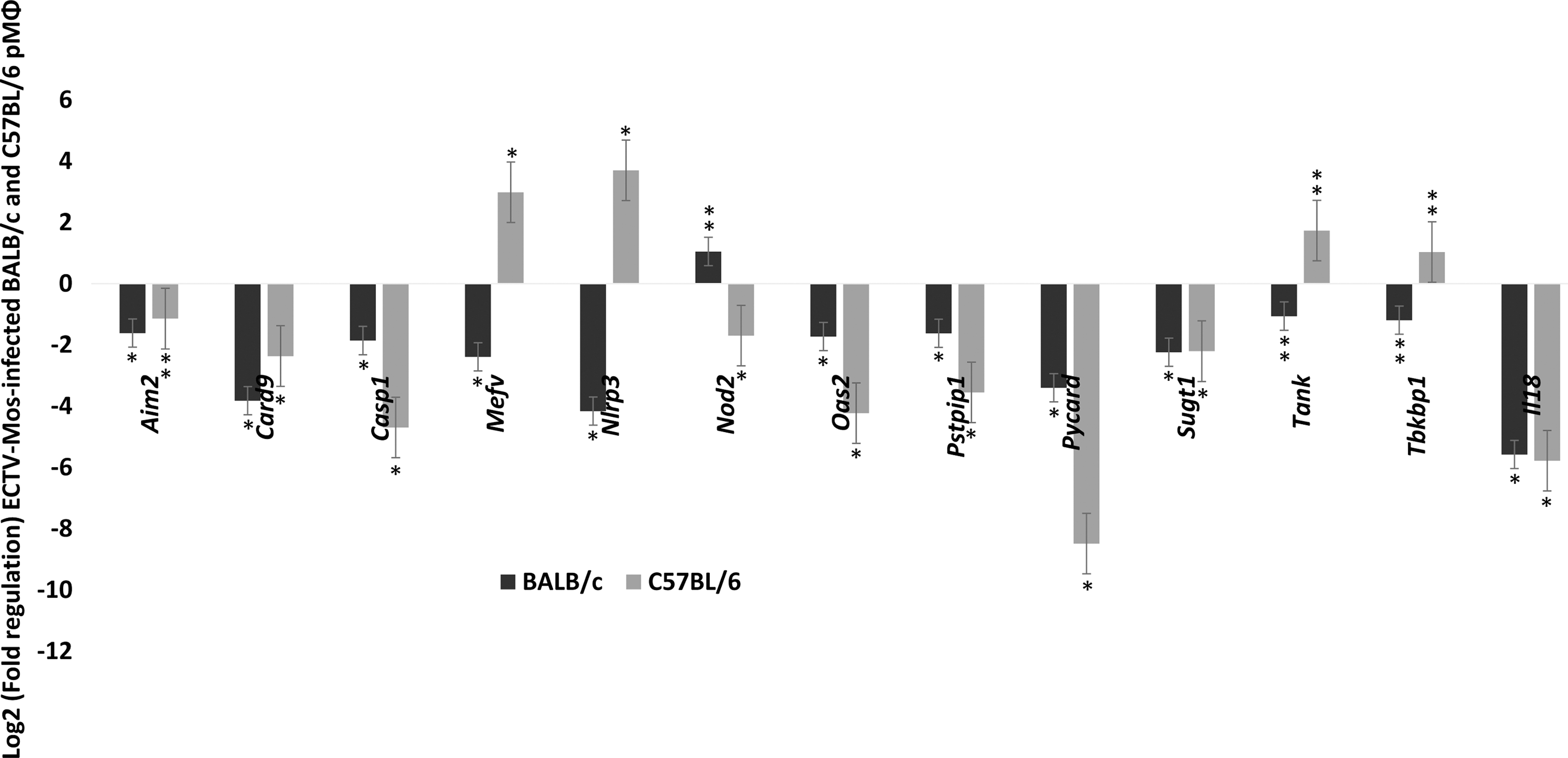
NOD-like receptor signaling genes are downregulated in both BALB/c and C57BL/6 mice. After ECTV-Mos infection, fold regulation of mRNA expression in pMΦ of BALB/c and C57BL/6 infected with ECTV-Mos. Receptors, signaling genes, and responsive genes. Experiments were repeated three times with at least five mice in each treatment group. *p ≤ 0.05, **p ≥ 0.05.
RIG-like receptor gene expression following infection
A total of 37 genes in the RIG-I-like receptor signaling category were examined. Only one gene in the receptor and chaperone cluster (Fig. 6A) was differentially expressed; Trim25 was upregulated in C57BL/6 mice compared to BALB/c mice where it was downregulated. The majority of the genes in the downstream signaling cluster were downregulated in both strains of mice (Fig. 6B).
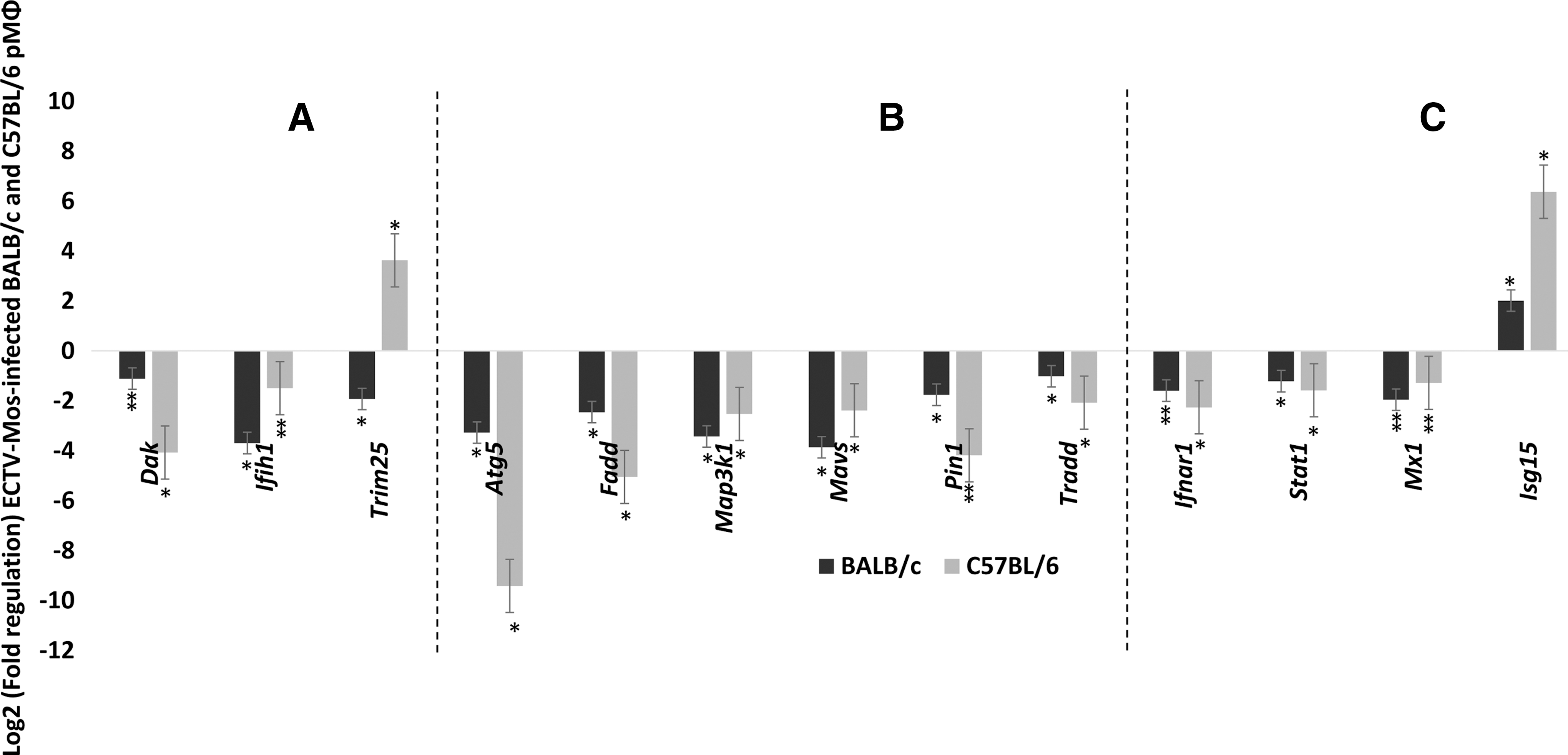
IRIG-I-like receptor signaling is downregulated in pMΦ of BALB/c and C57BL/6 mice after ECTV-Mos infection.
Type I IFN signaling gene expression
The four important genes in this group were downregulated (Fig. 6C). The result for Mx1 was expected because this gene has a mutation in standard C57BL/6 and BALB/c laboratory mice (36). However, Isg15 was highly expressed in infected cells from both mouse strains (p ≤ 0.05) (Fig. 6C).
Differentially expressed genes
Among all the genes expressed differentially between the two mouse strains in all gene categories, only 14 genes reached statistical significance (p ≤ 0.05) (Fig. 7). All the 14 were upregulated in C57BL/6 mice compared to downregulation in BALB/c mice.
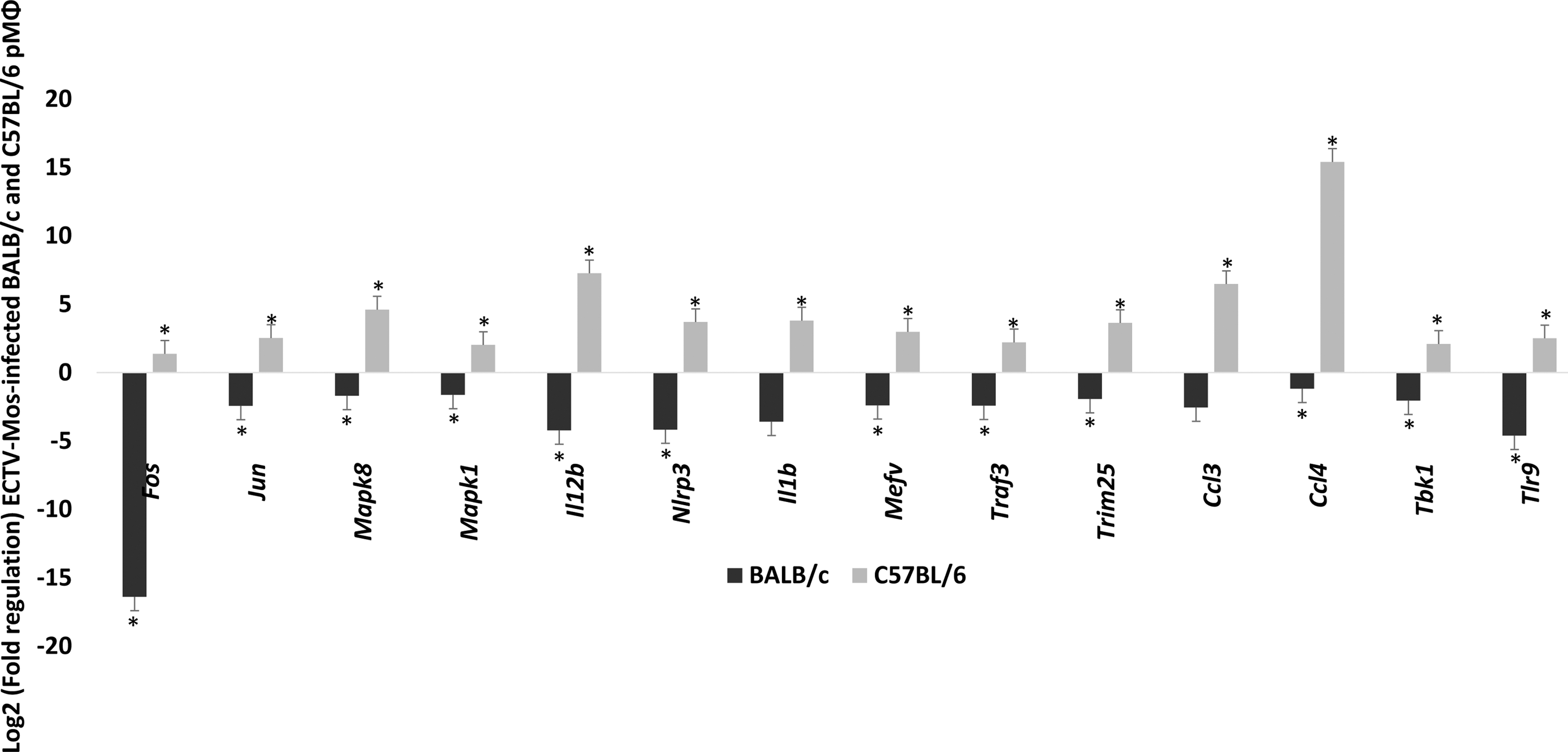
Differentially expressed genes. C57BL/6 mice have an advantage of 14 genes that are upregulated compared to BALB/c mice. *p ≤ 0.05.
Protein expression measured by Western blot
The mRNA expression patterns shown here would only be reliable if they correlated with protein expression in the studied cells. However, it is known that mRNA expression does not always correlate with protein expression in mammalian cells (41). To attempt a correlation between the observed mRNA expression levels and protein expression, we measured the expression of selected proteins in pMΦ originating from ECTV-Mos-infected and control mice of both strains. We measured all proteins, secreted or nonsecreted, with Western blot so as to avoid subculturing harvested cells before ELISA or flow cytometry and therefore unnecessary activation of cells. Of the significantly differentially expressed mRNAs, only five genes were confirmed to have a protein expression pattern that visually correlated with the mRNA profile. In Figure 8A, we show that NLRP3 was downregulated in BALB/c pMΦ from ECTV-Mos-infected mice, but upregulated in C57BL/6 pMΦ from ECTV-Mos-infected mice. Nod2 was upregulated in infected pMΦ from BALB/c mice, but downregulated in pMΦ from infected C57BL/6 mice. Although Nod2 and NLRP3 have been shown to have a functional synergism in other systems (9), their protein expression profile during infection with a poxvirus may not support cooperation of these proteins. Similarly, IRF7 was upregulated in BALB/c mice infected with ECTV-Mos, while it was downregulated in the similarly treated C57BL/6 mice. Conversely, MAPK8 was downregulated in BALB/c pMΦ from infected mice and upregulated in C57BL/6 mice. Although they were not differentially expressed in the two mouse strains, the cathepsin genes' mRNA expression matched the protein expression pattern.
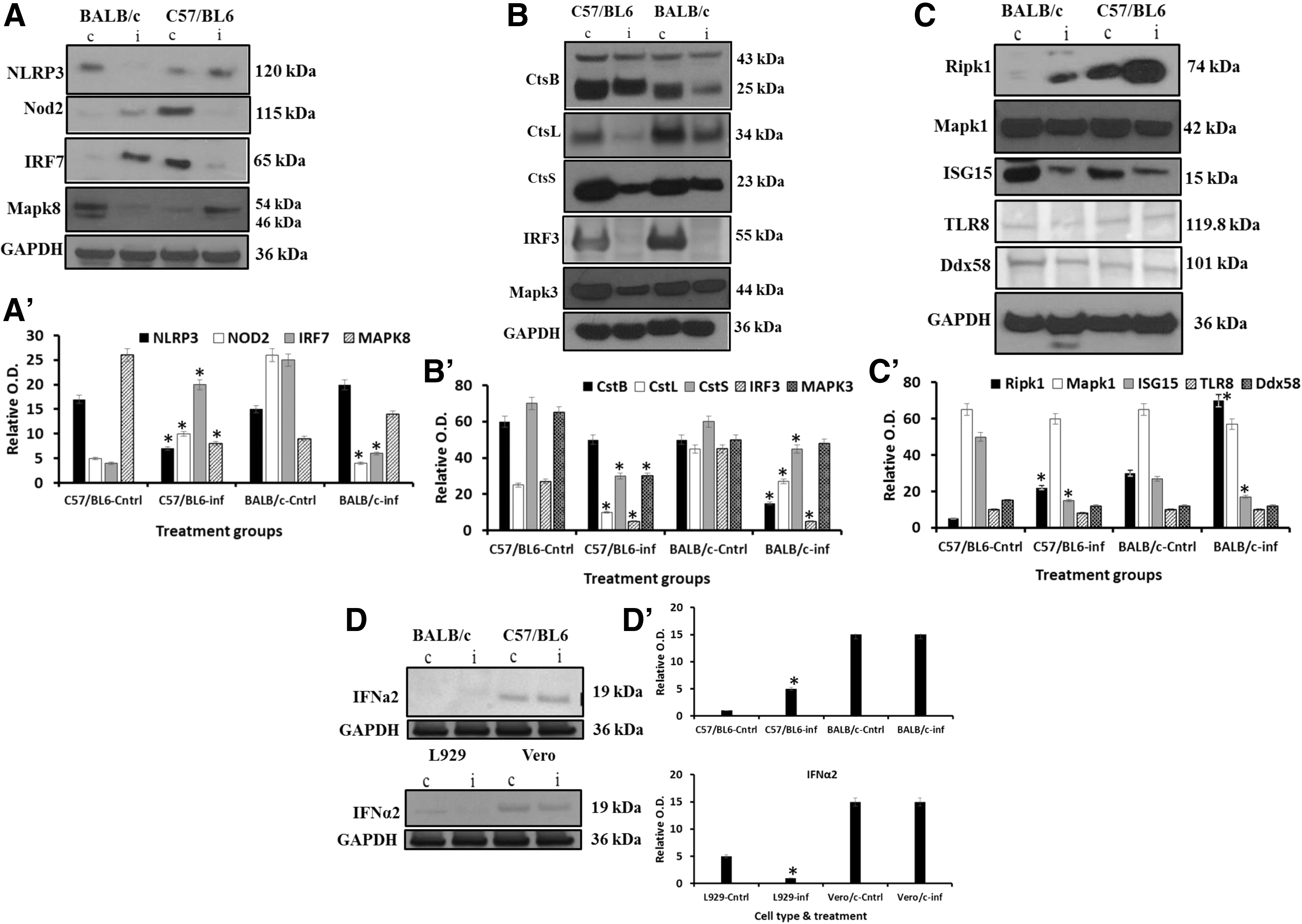
Protein expression of selected genes in pMΦ of BALB/c and C57BL/6 mice infected with ECTV-Mos. Western blot analysis of
Cathepsins B, L, and S (Fig. 8B) were all considerably downregulated in murine pMΦ from ECTV-Mos-infected mice. Cathepsin B, which usually occurs in two forms, a proenzyme and an activated form, only exhibited a reduction in the expression level of the activated form, while the proenzyme remained unchanged. Among the genes selected for protein expression confirmation, the mRNA expression coincided with protein expression in the cases of IRF3 and MAPK3 (Fig. 8B) in both mouse strains. Although the mRNA expression of IRF3 was not significantly downregulated in BALB/c mice (p ≥ 0.333), the protein level appeared to be appreciably reduced, such that we could not detect the protein band by Western blotting (Fig. 8B). In this case and other cases of mRNA and protein level discrepancy, we anticipate proteins could have been degraded by virus or simply short half-life of proteins. As for MAPK3, the statistically significant (p ≤ 0.05) downregulation of mRNA expression did not completely correspond to an equally reduced amount of protein expression, particularly in BALB/c mice, although it was evident that viral infection negatively affected the expression level.
We could not reconcile the protein expression with the mRNA expression for Ripk1, Mapk1, and Isg15. The mRNA expression was reduced for Ripk1 (p ≤ 0.004 and p ≤ 0.009 in C57BL/6 and BALB/c mice, respectively; Fig. 3B), while the protein expression was increased compared to noninfected murine pMΦ (Fig. 8C), particularly in C57BL/6 mice. We do not have a valid explanation of this occurrence. However, it is likely that different mechanisms involving cis-acting and trans-acting mechanisms may generate a big repertoire of systems that enhance the synthesis of proteins from a certain copy number of mRNA molecules (24). Mapk1 mRNA was differentially expressed; it was upregulated in C57BL/6 (p ≤ 0.02) mice but downregulated in BALB/c mice (p ≤ 0.003) (Supplementary Tables S1 and S2). However, the protein expression was reduced in both strains as determined by Western blotting (Fig. 8C). As for Isg15, the mRNA expression was significantly upregulated in both strains of mice (p ≤ 0.006 and p ≤ 0.009 for BALB/c and C57BL/6, respectively) (Fig. 6A), but the protein expression was downregulated in both strains following infection with ECTV-Mos (Fig. 8C). No difference in protein expression was observed for TLR8 or Ddx58 (RIG-I) in both strains (Fig. 8C); however, the mRNAs were downregulated (Fig. 3A and Supplementary Tables S1 and S2), although the decrease was not statistically significant (p ≥ 0.079 and p ≥ 0.43, respectively).
It was not surprising that IFNα mRNA was significantly higher (p ≤ 0.0014) in C57BL/6 mice despite the notion that poxviruses inhibit expression of this important innate cytokine. Rubio et al. (30) have previously shown the increase in expression of this gene. The protein expression did not reflect the mRNA expression in pMΦ infected with ECTV-Mos (Fig. 8D). In BALB/c mice, the mRNA expression increased, but it was not statistically significant, and we could not detect the protein by Western blotting in either infected or control mice (Fig. 8D). This is likely because IFNα is a secreted protein.
CBA measurement of protein expression
For the proteins that did not have a suitable Western blot antibody, we attempted a CBA to assess the protein expression. We assessed CCL3, CCL4, CCL5, CXCL9, and total STAT1 levels. We did not observe any significant differences in protein expression between mouse strains or the different treatments for CCL5 (Fig. 9). In this particular case, the mRNA expression in both mouse strains was significantly increased. Although CCL3 and CCL4 were differentially regulated at the mRNA level (Fig. 4), the protein expression was increased in C57BL/6 mice infected with ECTV-Mos, while no significant changes were observed for either protein in BALB/c mice (Fig. 9). Stat1 mRNA did not change for either mouse strain during infection. However, the protein quantity in pMΦ increased in both infected mouse strains (Fig. 9).
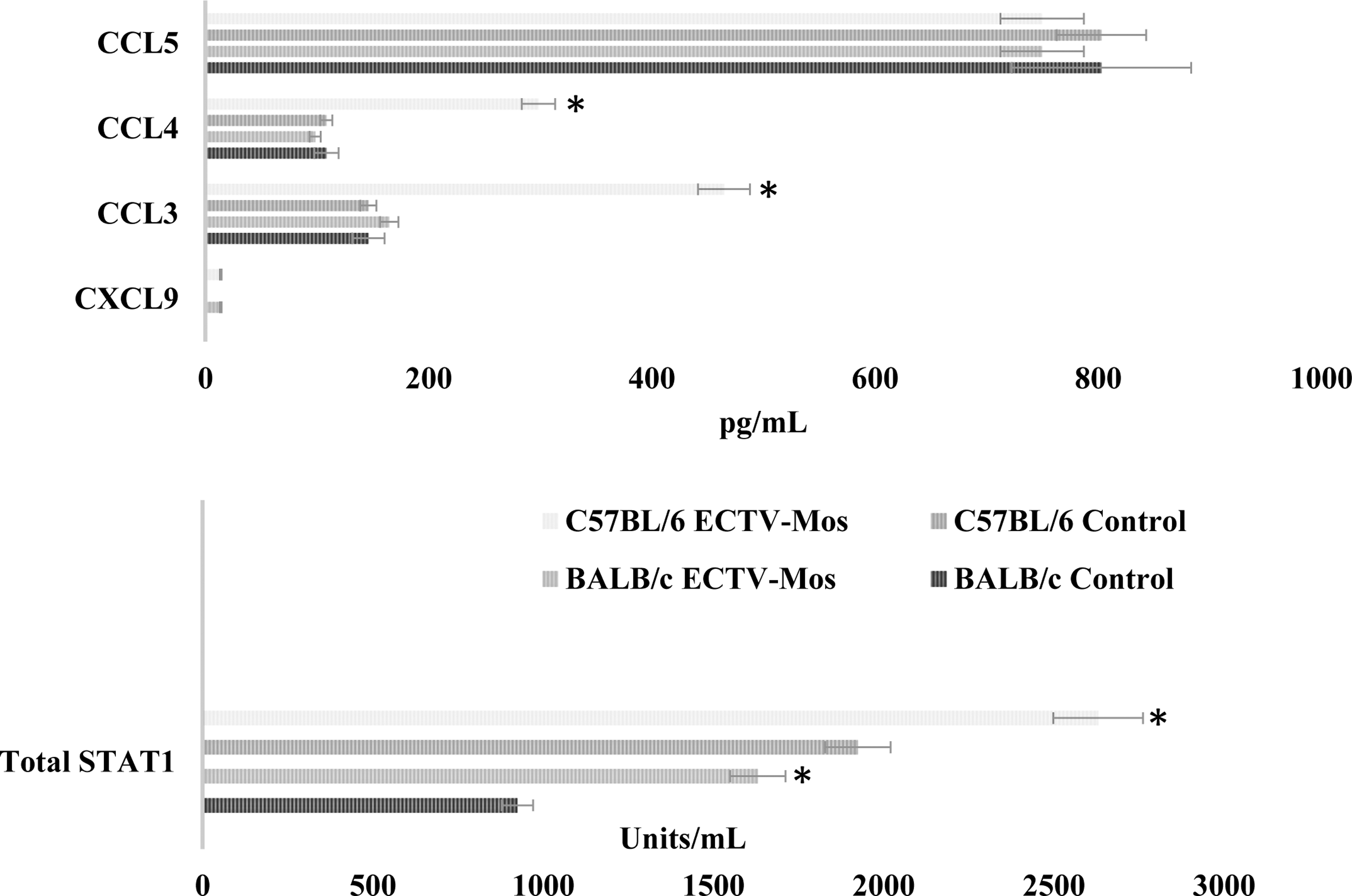
Protein expression levels for selected genes in pMΦ of infected and noninfected BALB/c or C57BL/6 mice. On collection of the cells as described in the Materials and Methods section, the cell lysates were prepared and the CBA assay performed according to the manufacturer's instructions. *p ≤ 0.05. pMΦ prepared at 48 h postinfection. Experiments were repeated three times with at least five mice in each treatment group.
siRNA knockdown of cathepsin B or L increases ECTV titer
To follow up on the implications of the observed downregulation of cathepsins B, L, and S, we assessed the immediate effect on the replication of ECTV in a macrophage cell line RAW264.7. As described in the Materials and Methods section, Table 3 shows the titers obtained in cells infected with ECTV after knockdown of cathepsin B or L. Although ECTV replicated in all cells irrespective of treatment, a more efficient replication was evident by higher titers in cells with knockdown of cathepsin B or L.
Supernatants were collected after 48 h and assayed for virus titer as described earlier. Experiments were repeated at least three times. * p ≤ 0.05 in comparison to control infected cells, **result not statistically significant in comparison to control infected cells, p > 0.05.
Discussion
We used RT2 profiler PCR arrays, Western blotting, and CBAs to compare the mRNA and protein expression patterns of innate immunity genes between pMΦ originating from C57BL/6 and BALB/c mice lethally infected with a murine poxvirus. The results show dramatic downregulation of pathogen-sensing mechanisms following infection, including the downstream signaling components of innate immunity. At the molecular level, many innate immunity genes were negatively affected in BALB/c mice compared to C57BL/6 mice. BALB/c mice are highly susceptible to murine ECTV compared to C57BL/6 mice. Knowledge of the host response elicited by poxviruses is critical for understanding of virus antiviral escape mechanisms and for designing new poxvirus vectors with improved immunogenicity. This study contributes valuable insight to molecular mechanisms of pathogenesis, specifically what genes may participate in the susceptibility to lethal poxvirus infection.
At least 14 genes are significantly differentially regulated between the two mouse strains. Some of the genes among the 14 have been studied in more detail either in VACV infection or other virus systems. For instance, Wang et al. (44) showed that Mapk8, a terminal kinase, is involved in generation of CD8+ T cell responses to ECTV. Mice deficient in JNK1/2 were not able to develop a CD8+ T cell response. Moreover, Hu et al. (20) also showed increased oncolytic VACV replication in murine embryo fibroblast in the absence of JNK1/2. Using a cell line 293T, Schweneker et al. (35) showed sustained stimulation of Mapk1 (ERK1) on infection with modified Ankara VACV or chorioallantoic VACV. A mechanism has recently been described where C6, a virulence factor from VACV, binds Tbk1 adaptor proteins and inhibits activation of IRF3 (40). In addition, IFN responses could be hampered by the reduction of Tbk1, since Tbk1 knockout macrophages failed to induce an IFN response (26). Ddx3x is targeted by VACV K7 leading to inhibition of IRF3 activation (34).
Myxoma virus infection of human monotypic THP-1 cells activates NLRP3 and leads to production of IL-1b, which is consistent with our finding in C57BL/6 at the mRNA level. Suppression of NLRP3 would entail reduction of the IL-1b, thus reducing the capability to mount an inflammatory response as has been reported by Delaloye et al. (12) in NLRP3 (NALP3)-deficient mice. Our study confirms this observation in BALB/c mice, where downregulation of Nlrp3 led to downregulation of both Il1β and Il18 genes. However, we found a discrepancy in the expression of Il18, which was downregulated as opposed to upregulation of Nlrp3 and Il1β in C57BL/6 mice. This difference may relate to susceptibility and resistance to ECTV infection of the two mouse strains. The direct role of Mefv in viral infection has not been described; however, its role may be closely associated with the function of the NLRP3 inflammasome. TRIM25 has not been assigned a role in poxvirus infection, but in other virus systems it appears to induce the Lys 63-linked ubiquitination of RIG-I, which is crucial for the cytosolic RIG-I signaling pathway to elicit host antiviral innate immunity (14). It is known that chemokines are affected by poxvirus infection (3), which was highly visible in studies in the BALB/c mice both at the mRNA and protein level. Collectively, these genes were downregulated in BALB/c mice, and therefore, it is highly probable that this difference could tip the balance toward susceptibility to lethal infection by ECTV-Mos, while C57BL/6 have more chances to mount a protective immune response. It is important, however, to note that our conclusion may be regarded to be general, mostly because of the in vivo character of this study, which presents some shortcomings to the approach. For instance, (i) the difference in susceptibility between the two mouse strains means that the virus titer at the time of analysis will be different in the two mouse strains, (ii) the degree of virus-induced cell death will be different between the two mouse strains, (iii) we cannot precisely distinguish between effects of infected cells and those induced by cytokines in uninfected cells in the harvested sample.
In the results reported here, we show a profound reduction in both mRNA and protein expression of cathepsins B, L, and S following infection with ECTV-Mos. Currently, studies are underway to investigate if this allows the virus to rapidly replicate in infected cells or if it is a cellular strategy to abate viral replication. Cathepsins B and L have been reported to play an essential role in the cellular entry of Ebola virus, which is mediated by its surface glycoprotein (33). In that study, cathepsins B and L appeared to convert the 130 kDa virus glycoprotein on the Ebola pseudovirions to a 19 kDa molecule, which apparently enhanced infection. In relation to the poxvirus used in our studies, we speculate that this may be an innate cellular mechanism to control viral infection because higher fold changes were observed in C57BL/6 mice than in BALB/c mice. C57BL/6 mice are resistant to infection, whereas BALB/c mice are highly susceptible. The replication of ECTV following transient removal of cathepsin B or L using siRNA shows that ECTV may downregulate cathepsins to its benefit. The cells with minute expression of cathepsin B or L had higher virus titers indicating that the virus replicated efficiently. The likelihood of cathepsin involvement in innate immune response is well explained in reports by Conus et al. (10) and Conus and Simon (11), where cathepsins influence apoptosis of neutrophils, affect toll-like receptor signaling, and activate certain proinflammatory cytokines. Cathepsins such as CatS and CatB are implicated in the generation of TAP-independent MHC class I cross-presentation (37). Moreover, Wang et al. (43) have reported the reduction in expression, both at the mRNA and protein level, of CatS, CatB, and CatD following infection of B cells with VACV. In those studies, the implication was that these enzymes played a role in antigen and Ii chain processing. Also, cathepsin B is involved in the trafficking of TNF-α-containing vesicles to the plasma membrane in macrophages (16). In light of these findings, the profound reduction in cathepsin expression in C57/BL6 mice is puzzling, particularly that virus titers in these mice were lower compared to BALB/c mice. One possibility is that the complexity of networks induced by poxvirus infection leading to this viable antiviral mechanism being downregulated makes distinct host activators necessary to ensure effective inhibition of poxvirus replication. Another possibility is that downregulation of cathepsin may indicate yet another unidentified antiviral mechanism engaged by poxviruses. Either way this aspect requires further study.
The TLR genes were equally downregulated in both mouse strains, although more receptor and chaperone genes were downregulated in BALB/c mice compared to C57BL/6 mice. These results are contrasted by the upregulation of response genes at the mRNA level, such as type I IFN (Ifnα2 and Ifnβ1), in both strains of mice. In BALB/c mice, this can partially be explained by the maintenance of Irf7 at an upregulated level. IRF7 has been reported to activate Ifnα and Ifnβ (19). Similarly, Cao et al. (7) showed a reduction in the expression of TLR7 and TLR9 in human plasmacytoid DCs infected with VACV.
Innate immunity is often stimulated by the sensing of PAMPs. For viruses, cytosolic pathogen recognition receptors (PRRs), in addition to TLR9, are the most important for detecting PAMPs. RIG-I and MDA5 detect viral RNA or RNA intermediates, leading to the activation of IRF3/IRF7 and NK-κB signaling (1). In addition to TLR9, the viral sensing system in C57BL/6 mice was highly downregulated; however, some of the downstream signaling molecules were upregulated, which appeared to strengthen the type I IFN pathways and proinflammatory cytokine maintenance, as evidenced by the mRNA expression pattern of the response gene cluster. It appears that RNA intermediates may not be sensed properly, and it is likely that the type I IFN pathway is induced through dsDNA sensors as described by Takaoka et al. (38). The genes encoding dsDNA sensors were not included in the gene set analyzed in our studies. The Ifih1 gene encodes MDA5, an RIG-I-like receptor with helicase enzymatic activity (21), and it has been implicated in blocking viral infection through the induction of IFN-β.
The activation of type I IFN appeared intact only in C57BL/6 mice based on the upregulation of Tbk1, Tnf, Irf3, Ikbkb, and Traf3. These genes have been described to be important for the induction of type I IFN in macrophages and DCs (39). The failure to detect type I IFN in BALB/c mice or increases in this protein in C57BL/6 mice could be explained in terms of production of this type of IFN, which is a secretory protein. Only plasmacytoid DCs are defined as natural IFN-producing cells. The canarypox virus (ALVAC) used in the studies conducted by Harenberg et al. (17) on monocyte-derived DCs enhanced the expression of ISG15 at both the mRNA and protein levels. In murine pMΦ from both C57BL/6 and BALB/c mice, we could not reconcile the mRNA and protein expression; the mRNA was upregulated, while the protein was downregulated. However, the expression of ISG15 protein may be dependent on type I IFN (27). We do not show evidence of enhanced expression of type I IFN protein.
A profound inhibitory influence was exerted on the NOD-like receptor signaling pathway in peritoneal macrophages during infection with ECTV-Mos. In the NOD-like category, only Nod2 was unaltered, while the remaining 13 genes analyzed were significantly downregulated. On the contrary, Nod2, a molecule that functions as a cytoplasmic viral PRR, was upregulated in BALB/c mice. It does not appear to be a sensitive monitor of RNA intermediates during poxvirus infection. Similarly, Sabbah et al. (31) showed no effect of VACV activation on Nod2.
Of the 37 RIG-I-like signaling components in the antiviral response, only two responsive genes, Ifna2 and Ifnb1, were upregulated. Our results relating to BALB/c mice partially agree with those of Harenberg et al. (17), although the previous study was conducted on human DCs using a canarypox virus, ALVAC. They reported an increase in the gene expression of RigI, Irf7, Cxcl10, and Isg15. This would be in accordance with our observed increase in type I IFN in BALB/c mice. However, this mouse strain has been deemed highly susceptible to ECTV. It is possible that the receptors exhibit multiple functions during the response to viral infection, when RNA intermediates are produced during replication.
While we did not assay the phagocytic capacity of infected pMΦ in these studies, we speculate that this function could have been impaired to some extent because of the significant downregulation of at least three cathepsin proteins. Earlier, Gledhill et al. (15) showed reduced phagocytosis during infection with the viruses that produce hepatitis, including ECTV.
Conclusion
In conclusion, we have observed a massive remodeling of the host transcriptome in pMΦ originating from mice infected with a poxvirus. Although peritoneal macrophages, in this particular infection, are not in the forefront of mediators of a protective immune response, the results show how a single virus is able to affect numerous host genes to downregulate innate immune responses. With at least a 14-gene disadvantage, these data provide molecular evidence of the increased susceptibility of BALB/c mice to infection with ECTV-Mos. We have identified a drastic change in the expression of cathepsins. These genes will be used in functional studies to understand the contribution of macrophages to the pathogenesis of poxvirus infection.
Ethics Approval
The 3rd Local Ethics Committee for Animal Research at the Warsaw University of Life Sciences-SGGW (Permit number 41/2010) approved the experimental protocols.
Footnotes
Acknowledgments
This work was supported by the NCN Grant NN401015240 to F.N.T. and Centre for Integrative Mammalian Research, Ross University School of Veterinary Medicine. The funding bodies did not play a role in the design of the study, collection, analysis, interpretation of data, and in writing the manuscript.
Authors' Contributions
P.D. performed the in vivo experiments; L.S.-D. performed all immunofluorescence studies; M.M. and M.B. performed the in vivo experiments; Z.N. performed RT-PCR experiments and interpreted RT-PCR data; F.N.T. conceived the study, designed, performed, and interpreted RT-PCR data and drafted the manuscript, coordinated the work, and revised the manuscript critically for important intellectual content.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.