
Abstract
Recently identified T memory stem (Tscm) cells have stem-cell-like properties, including long lifespan, self-renewal capacity, and multipotency to differentiate into other memory T cell types. In the study of simian immunodeficiency virus (SIV) infection, selective depletion of CCR5+CD4+ Tscm cells and the high proliferation rate of these cells are believed to be responsible for the pathogenesis of SIV-infected rhesus macaques. Here, we conducted a cohort study to investigate the influence of chronic human immunodeficiency virus (HIV)-1 infection on CD4+ Tscm cell homeostasis, and the effect of antiretroviral therapy (ART) on CD4+ Tscm cells. Chronic HIV-1 infection resulted in a decrease of the CD4+ Tscm cell proportion in HIV-1 patients. The decreased number of CD4+ Tscm cells in HIV-1 patients correlated positively with that of circulating CD4+ T cells. Further, the depletion of CD4+ Tscm cells was inversely correlated with an increased level of T cell immune activation during chronic HIV-1 infection. Prolonged ART recovered the CD4+ Tscm cells, and the dynamic change of CD4+ Tscm cells was in parallel with CD4+ T cell restoration and a decrease in the level of T cell immune activation. We propose that the abnormity of CD4+ Tscm cells may contribute to the pathogenesis and disease progression in HIV-1-infected individuals.
Introduction
M
There is increasing evidence demonstrating that Tscm cells play a crucial role in human immunodeficiency virus (HIV)/simian immunodeficiency virus (SIV) pathogenesis. CD4+ Tscm cells are susceptible to both CXCR4- and CCR5-tropism HIV-1 strains and can be a stable HIV-1 latent reservoir regardless of successful antiretroviral therapy (ART) (2,5,7,10,15,17,29). CD8+ Tscm cells correlate with HIV-1 disease progression (24). Maintenance of HIV-1-specific CD8+ Tscm cells during long-term ART raises the possibility that a combination of HIV-1-specific CD8+Tscm cells and latency-reversing agents may support the elimination of the HIV-1 residual reservoir (30). An SIV study showed that, in contrast to pathogenic SIV-infected rhesus macaques (RMs), the resistance of CCR5+CD4+ Tscm cells to SIV infection and the preservation of these cells are responsible for the nonpathogenic infection in SIV natural hosts such as sooty mangabeys (SMs) (3). ART induces restoration of CCR5+CD4+ Tscm cells in blood and lymphoid nodes of RMs (4). However, the proviral DNA in CD4+ Tscm cells does not decline in RMs after ART, supporting the contribution of Tscm cells to persistent SIV infection (4).
Thus far, the abnormity of CD4+ Tscm cells in HIV-1 infection and the effect of ART on the dynamic change of CD4+ Tscm cells are not well documented. Therefore, we conducted longitudinal analysis of CD4+ Tscm cells in the circulating blood of HIV-1 patients by using an ART-treated cohort. We found that HIV-1 infection resulted in a significant decrease in the proportion of CD4+ Tscm cells. The reduction of CD4+ Tscm cells was correlated with the loss of circulating CD4+ T cells and the high level of T cell immune activation in HIV-1 patients. In addition, prolonged ART induced restoration of CD4+ Tscm cells. Increased CD4+ Tscm cells contributed to the recovery of circulating CD4+ T cells and the decline of T cell immune activation to a normal level. Overall, our study further implicates the important role of CD4+ Tscm cells in the pathogenesis and immune reconstitution of HIV-1 patients.
Materials and Methods
Patients and samples
Twenty-five chronic HIV-1 patients were recruited from the ART-treated chronic patient cohort in Beijing You'an Hospital. Thirty-six age-matched healthy controls (HCs) were also included in this study. No subject had hepatitis B or C virus co-infection.
After baseline evaluation, HIV-1 patients began treatment with antiretroviral drugs and were followed up every 3 months. The ART regimen included AZT (Zidovudine)/3TC (Lamivudine)/NVP (Nevirapine) (n = 12) or TDF (Tenofovir)/3TC/EFV (Efavirenz) (n = 13). Blood samples from the HIV-1 patients were collected at baseline and at each followed-up time point. Peripheral blood mononuclear cells (PBMCs) from whole blood were isolated by density gradient centrifugation and stored in liquid nitrogen. All participants in this study provided written informed consent. This study was approved by the Beijing You' an Hospital Research Ethics Committee.
Immunophenotyping
Cryopreserved PBMCs were thawed in RPMI 1640 medium (Invitrogen, Carlsbad, CA) and washed with phosphate-buffered saline (PBS) containing 1% bovine serum albumin. The cells were stained with the following anti-human antibodies purchased from Biolegend (San Diego, CA): CD3 APC-cy7 (SK7), CD4 FITC (SK3), CD8 Percp-cy5.5 (RPA-T8), CD45RA PE-cy7 (HI100), CCR7 APC (G043H7), CD27 PE (O323), CD95 BV421 (DX2), CD38 PE (HIT2), and HLA-DR APC (TU36). At least 50,000 live lymphocytes were analyzed with a BD FACSCanto II flow cytometer (BD Biosciences, San Diego, CA). The final analysis was performed by using Flowjo 7.6.1 software (Tree Star, Inc., Ashland, OR).
Measurements of CD4+ T cell numbers and plasma viral load
Routine CD4+ T TruCount (BD Biosciences) and plasma HIV-1 viral load (pVL, Versant440; Siemens Healthcare Diagnostics, Germany) were measured according to the manufacturers' instructions. The detecting limitation of pVL is 50 copies/mL.
Statistical analysis
All statistical analyses were performed by using GraphPad Prism software v5.0 (GraphPad Software, Inc., La Jolla, CA). Statistical significance between two groups was calculated by using the nonparametric Mann–Whitney U-test, whereas comparisons among ART periods (Fig. 3) were determined by the Kruskal–Wallis one-way analysis of variance (ANOVA) with Dunn's comparison test. Correlations were determined by the Spearman rank correlation test. A p-value of less than 0.05 represented significance.
Results
Loss of CD4+ Tscm cells in HIV-1 infection
Twenty-five chronic HIV-1 patients and 36 HCs were included in our study. The baseline characteristics of HIV-1 patients and HCs are presented in Table 1. Table 2 is the CD4+ T cells and pVL pre-ART or on ART in each individual. All subjects were male. The frequency of CD4+ T cells in HIV-1 patients was significantly lower than that in HCs (p < 0.0001). The total circulating CD4+ T cell count was not compared between HIV-1 patients and HCs because of a lack of CD4+ T cell data from HCs. ART resulted in the decrease of pVL to an undetectable level in all patients. To determine the influence of chronic HIV-1 infection on CD4+ Tscm cells, we compared the frequency of circulating CD4+ Tscm cells in HIV-1 patients and HCs. CD4+ Tscm cells were defined as CD3+CD4+CD45RA+CCR7+CD27+CD95+ (Fig. 1A). As shown in Figure 1B, the CD4+ Tscm cell proportion was decreased significantly by chronic HIV-1 infection compared with HCs (p < 0.0001).

Decreased proportion of CD4+ Tscm cells in chronically HIV-1-infected patients.
HIV, human immunodeficiency virus; NA, not applicable.
p < 0.0001 compared with healthy controls.
NA, not applicable.
TND, pVL <50 copies/mL.
Baseline CD4+ Tscm correlate with disease progression markers
To investigate the significance of CD4+ Tscm cells in HIV-1 pathogenesis, we next evaluated the correlation between baseline CD4+ Tscm cells in HIV-1 patients and HIV-1 disease progression markers, including the total circulating CD4+ T cell count, pVL, CD4/CD8 ratio, and the level of T cell immune activation. We found that both the frequency and count of baseline CD4+ Tscm cells in HIV-1 patients were positively correlated with the total circulating CD4+ T cell count (r = 0.36, p = 0.04, r = 0.67, p = 0.0002, respectively, Fig. 2A, B). In addition, baseline CD4+ Tscm cell count was also positively correlated with CD4/CD8 ratio (Fig. 2C, D). However, there was no correlation between baseline CD4+ Tscm cells and pVL in HIV-1 patients (data not shown). In addition, the level of T cell immune activation is an important disease marker in HIV-1 pathogenesis. Here, co-expression of CD38 and HLA-DR on the surface of CD8+ T cells was used as the T cell immune activation marker. The number of CD38+HLA-DR+CD8+ T cells in HIV-1 patients was much higher than that in HCs (p < 0.0001, Fig. 2E). A significantly negative correlation between CD38+HLA-DR+CD8+ T cells and the CD4+ Tscm cell count was observed at baseline in HIV-1 patients (r = −0.45, p = 0.03, Fig. 2F, G).
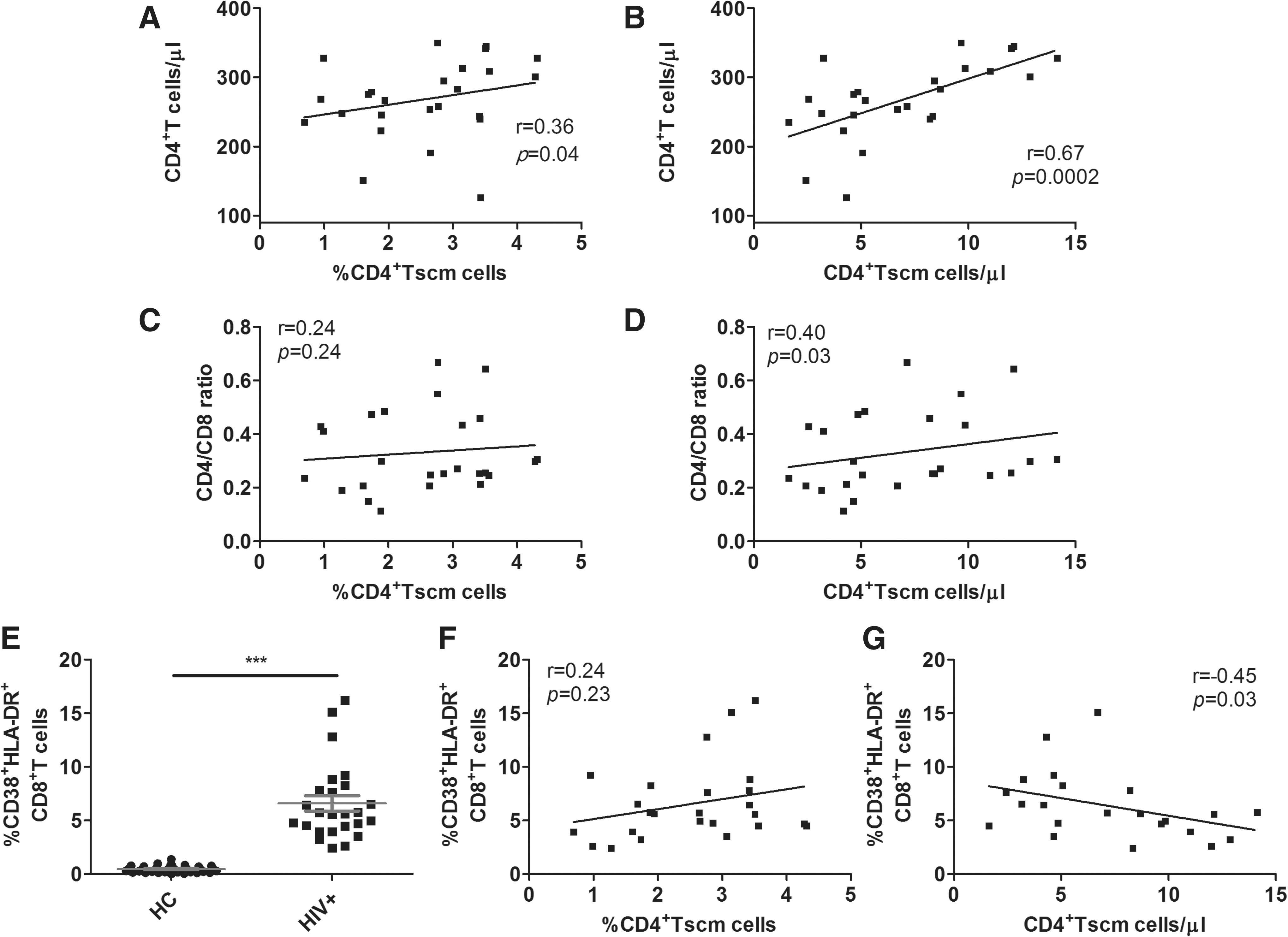
CD4+ Tscm cell counts are correlated with disease progression markers. Correlation between baseline CD4+ Tscm cells and CD4+ T cells
Dynamic change in CD4+ Tscm cells during long-term ART
A recent study showed that ART induces partial restoration of the proportion of CCR5+CD4+ Tscm cells but not total CD4+ Tscm cells in both PBMCs and lymph nodes during SIV infection (4). Therefore, we investigated the effect of ART on CD4+ Tscm cells in HIV-1 patients and the correlation between the change of CD4+ Tscm cells and disease markers. As expected, compared with the baseline, the circulating CD4+ T cell count increased gradually (p < 0.0001, Fig. 3A). Long-term ART resulted in the decrease of pVL to an undetectable level in all patients (p < 0.0001, Fig. 3B), and the level of CD38+HLA-DR+CD8+T cells was also reduced significantly (p < 0.0001, Fig. 3C). The CD4+ Tscm cell count was also increased at 144 weeks post-ART compared with at baseline (p < 0.01, Fig. 3D). Overall, the dynamic change in the CD4+ Tscm cell count in response to ART was similar to the change in circulating CD4+ T cells, and it was opposite to the change in the CD38+HLA-DR+CD8+ T cell level. Analysis of individual patients also demonstrated this close relationship between CD4+ Tscm cells and the circulating CD4+ T cell count or the level of T cell immune activation during prolonged ART (Fig. 3E, F).
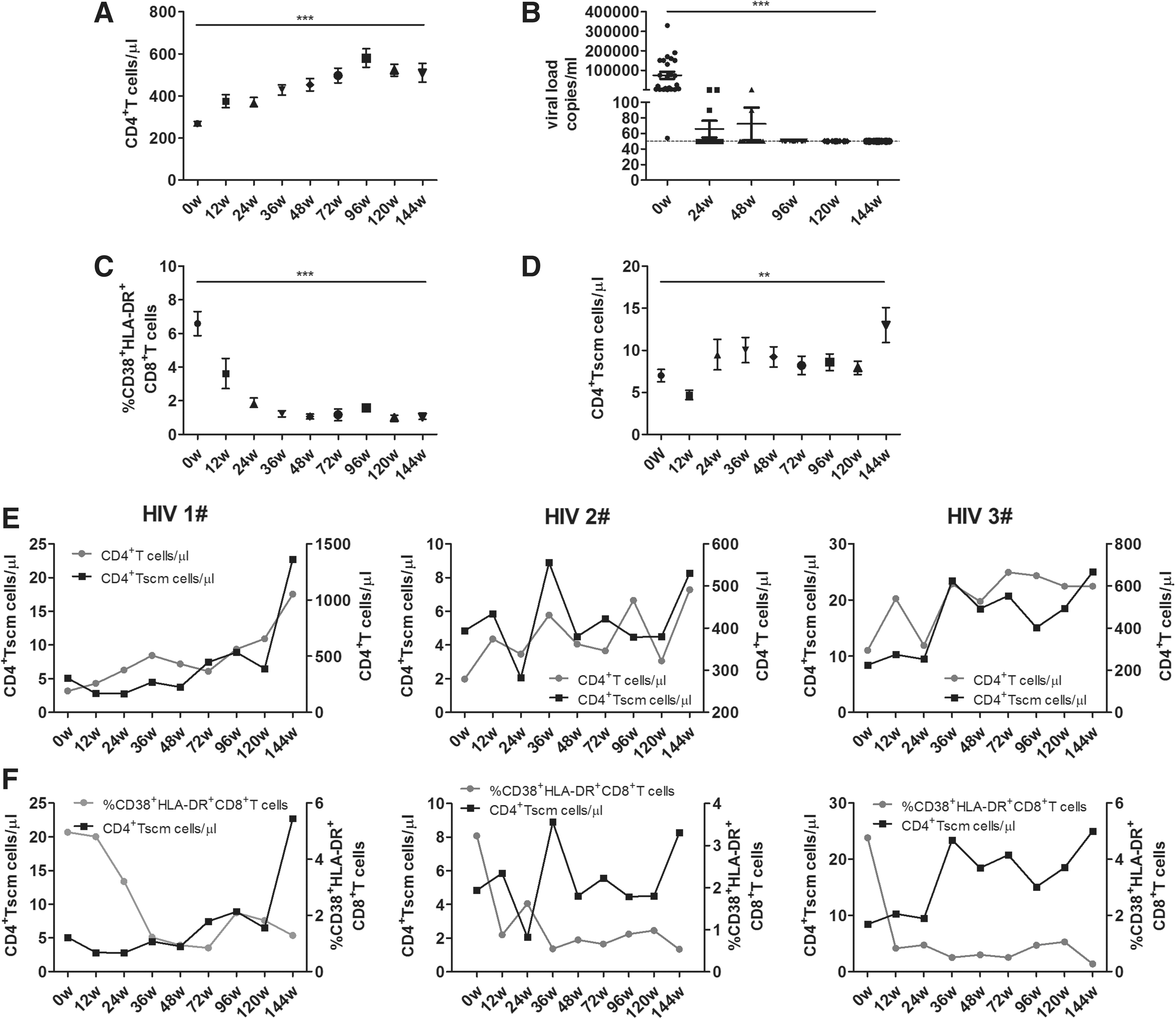
Response of CD4+ Tscm cells to ART in chronically HIV-1-infected patients. A total of 25 HIV-1 patients accepted stable ART for at least 2 years. Blood samples were collected at each followed-up time point. Changes in the CD4+ T cell count
Discussion
CD4 depletion is a hallmark of HIV/SIV infection. The pathogenic mechanisms of CD4+ T cell depletion have been extensively studied in the past 3 decades. A better preserved CD4+ Tcm cell compartment is a key mechanism that is responsible for HIV/SIV pathogenesis (6,20,22), and CD4+ Tcm cell restoration is a critical parameter for immune reconstitution after ART (14,21,23). A novel memory T cell subset, termed as Tscm cells, has been recently identified. These Tscm cells have superior proliferation and self-renewal capacity, and they are precursors of other T memory cell types (11). The dynamics of CD4+ Tscm cells have been well investigated in pathogenic and nonpathogenic SIV infection of NHP models in the past 2 years (3,4). Therefore, in this study, we focused on how CD4+ Tscm cells are affected by HIV-1 infection and the contribution of CD4+ Tscm cells to HIV-1 disease progression.
The main findings of our study are that chronic HIV-1 infection results in a reduction of CD4+ Tscm cells, and the loss of these cells positively correlates with a CD4+ T cell decrease and inversely correlates with the level of T cell immune activation. In addition, ART restores CD4+ Tscm cells, and the dynamic change in CD4+ Tscm cells is closely related to that of CD4+ T cells and T cell immune activation in ART-treated HIV-1 patients.
CD4+ Tscm cells are more susceptible to R5-tropsim HIV strains than other memory T cell subsets (5). Compared with healthy SIV nonpathogenic SMs, RMs have a higher level of CCR5 expression, and selective infection and depletion of CCR5+CD4+ Tscm cells are observed in SMs, although SMs exhibit preservation of total CD4+ Tscm cells (3). In this study, a decrease of CD4+ Tscm cell was also found in HIV-1 patients (Fig. 1B). Therefore, the infection and loss of CD4+ Tscm cells could be a novel mechanism of HIV/SIV pathogenesis. In addition to the loss of CD4+ Tscm cells by direct virus infection, whether bystander apoptosis leads to CD4+ Tscm cell exhaustion should be investigated further.
Previous studies have demonstrated that stability of the CD4+ Tcm cell compartment is a key determinant in the timing of disease progression in HIV/SIV infection (20,22). Because of the ability to differentiate into other T memory cell types, CD4+ Tscm cells may also contribute to the maintenance of T cell homeostasis and disease progression of HIV/SIV infection. A comparative study between viremic non-progressors (VNPs) and putative progressors (PPs) revealed that VNPs preserve CD4+ Tscm cells with a higher proliferative potential compared with PPs (17). In addition, the limited infection of CD4+ Tcm and CD4+ Tscm cells is associated with the lack of progression in VNPs (17). In SIV-infected pathogenic RMs, increased CD4+ Tscm proliferation is the key feature distinguishing them from nonpathogenic SMs (3). The higher level of CD4+ Tscm cell proliferation inversely correlates with the proportion of CD4+ Tcm cells (3). In our study, a positive correlation was found between CD4+ Tscm count and CD4+ T cells pre-ART. Exhaustion of CD4+ Tscm cells may disturb the T cell homeostasis in HIV-1 or pathogenic SIV infections, and the CD4+ Tscm cell proliferation, at least in part, maintains T cell homeostasis and compensates for the loss of other memory T cells. Therefore, we proposed that the reduction of CD4+ Tscm cells may be responsible for the loss of CD4+ T cells during chronic HIV-1 infection. Further, CD4+ Tscm cells have a robust interleukin-2 secretion ability compared with other T memory cell types, which can help CD4+ Tscm cells to maintain their own homeostasis through proliferation (9,11).
Correction of the abnormity in CD4+ Tscm cells by ART administration was observed in our study and an SIV study, thus improving T cell homeostasis and immunological responses to HIV/SIV infection (Fig. 3) (4). However, ART cannot decrease SIV DNA in CD4+ Tscm cells in RMs, indicating persistent infection in CD4+ Tscm cells under ART (4). Consistent with RMs, the level of HIV-1 DNA in CD4+ Tscm cells does not decrease in HIV-1 patients (2). Therefore, a novel approach to reduce the CD4+ Tscm cell contribution to viral persistence should be developed to improve the outcome of HIV-1 treatment.
Decrease of T cell activation after ART has been well documented in many publications (16,25,28). Here, we observed a negative correlation between CD4+ Tscm cells and T cell activation at baseline, and an opposite dynamic change of them during long-term ART (Fig. 3). A study showed that the high frequency of CD4+ Tscm cells in the circulation expresses gut-homing marker α4β7 (24). Therefore, a potential mechanism of protective effect on T cell immune activation by CD4+ Tscm cells might be the maintenance of gut barrier function and the reduction of microbial translocation by the migration of α4β7+CD4+ Tscm cells from blood to gut mucosa.
In conclusion, this study provides evidence that HIV-1 infection induces the reduction of CD4+ Tscm cells, and CD4+ Tscm cells play an important role in both HIV-1 pathogenesis and disease progression.
Footnotes
Acknowledgments
The authors thank Wei Feng, Wei Xia, and Yuefang Zhou for the study cohort maintenance.
This study was supported by the National Natural Science Foundation of China (81501732 to XL), Beijing Municipal of Science and Technology Major Project (D161100000416003 to HW), the Funding for Chinese overseas talents returning to China in 2016 (to BS), and the Beijing Key Laboratory for HIV/AIDS Research (BZ0089).
Author Disclosure Statement
The authors of this article report no conflicts of interest.