
Abstract
Chikungunya virus (CHIKV), a member of Alphavirus genus, is responsible for chikungunya fever (CHIKF), which is characterized by the presence of fever, rash, myalgia, and arthralgia. Reemergence of CHIKV has become a significant public health concern in Asian and African countries and is newly emerging in the Middle East, Pacific, American, and European countries. Cytokines, innate (monocytes, natural killer cells) and adaptive immune response (role of B cells and T cells i.e. CD4+ and CD8+), and/or viral factors contribute to CHIKV-induced arthritis. Vector factors such as vector competence (that includes extrinsic and intrinsic factors) and effect of genome mutations on viral replication and fitness in mosquitoes are responsible for the spread of virus, although they are not directly responsible for CHIKV-induced arthritis. CHIKV-induced arthritis mimics arthritis by involving joints and a common pattern of leukocyte infiltrate, cytokine production, and complement activation. Successful establishment of CHIKV infection and induction of arthritis depends on its ability to manipulate host cellular processes or host factors. CHIKV-induced joint damage is due to host inflammatory response mediated by macrophages, T cells, and antibodies, as well as the possible persistence of the virus in hidden sites. This review provides insight into mechanisms of CHIKV-induced arthritis. Understanding the pathogenesis of CHIKV-induced arthritis will help in developing novel strategies to predict and prevent the disease in virus-infected subjects and combat the disease, thereby decreasing the worldwide burden of the disease.
Introduction
C
By 2006, CHIKF epidemic has been observed in the Indian Ocean island of La Reunion, including India, Malaysia, Indonesia, Thailand, Singapore, the United States, and European countries and has affected millions of people (37,62). During the Reunion Island epidemic, 244,000 cases of CHIKV infection were reported (116). CHIKF has also been reported from countries like Italy, France, New Caledonia, Papua New Guinea, Bhutan, and Yemen. CHIKV had caused more than 440,000 cases of disease in more than 20 countries in the Caribbean and Central and South America by July 2014 (95). Thus, in less than 10 years, CHIKV has spread over in 50 countries and reemerged as a true global pathogen and is a significant public health concern.
Fever and debilitating arthralgia are the classical symptoms of the disease (133). Asthenia, headache, vomiting, rash, and myalgia are also the commonly observed symptoms (30). CHIKV infection also leads to the development of neurological symptoms such as encephalitis, myelopathy, peripheral neuropathy, myeloneuropathy, and myopathy (16,117). CHIKV is also an important cause of neurological disorders in children and neonates (54,77). Chikungunya infection also lead to systemic complications and deaths in elderly adults (141). During 2005–2006, 254 deaths were documented during Réunion outbreak (case–fatality rate 1/1,000) due to CHIKF directly or indirectly (112). Excess number of deaths were reported from different parts of India during the CHIKF epidemic of 2006 as compared with the number of deaths reported during the same period in other years ranging from 2002 to 2008 (85,88).
CHIKV infection also leads to a wide spectrum of post CHIKV chronic rheumatic musculoskeletal pain and disorders, including nonspecific polyarthralgia, rheumatoid arthritis (RA)-like illness, undifferentiated inflammatory arthritis, soft tissue rheumatism, seronegative spondylarthritis, or psoriatic arthritis (PsA) (8,22,27). Persistent arthralgia was first suggested by Robinson (118). Fourie and Morrison first reported postarthritic syndrome (39). Brighton et al., reported a high prevalence of chronic multiple arthralgia after 3 years of disease onset (11). It is still not clear how CHIKV infection induces persistent arthritis despite a robust immune response, but CHIKV-induced arthritis has an antigen-specific immunopathologic origin, which is responsible for the development of chronic joint disease (36,53). In this review, we attempt to describe the viral and host factors (human and mosquito) with an emphasis on those mechanisms which are responsible for CHIK-induced arthritis immunopathology.
CHIKV: an arthritogenic virus
CHIKV is arthritogenic in nature and muscle tissues have been proposed to be their targets (32). Infection starts when CHIKV-infected mosquito bites the host and subcutaneously inoculate the virus followed by the initial viral replication and systemic viremic spread to secondary sites. “Primary viremia” symbolizes the acute phase of the infection. Muscle satellite cells and fibrobalsts in the skeletal muscles are the major sites of viral replication, which leads to the damage of striated muscle fibers and subsequent development of myalgia. Acute pahse might be more severe in newborns and elderly people. Acute phase is followed by subacute phase, which is marked by the appearance of virus-specific IgM antibodies and extensive inflammatory response leading to myositis. Subacute phase progresses into the chronic phase when the virus-specific IgG appears. IgM antibodies may also be present in the chronic phase and the presence of IgM antibodies in the chronic phase indicates the persistence of virus latently in muscle cells. The chronic phase is characterized by persistent and recurrent myalgia and arthralgia lasting for months to years. The symptoms during the chronic phase might resemble idiopathic inflammatory myopathies (51).
Pathogenesis of CHIKV-Induced Arthritis
CHIKV inoculated subcutaneously into the host body acquires the lymphatic route to disseminate to various organs. Virus replicates in the skin, host articular and nonarticlular cells, and it spreads to liver, spleen, and kidneys, which are considered as the primary sites of replication (70). It may proceed to bones, muscles, articular tissues, and joints, increasing the severity of the disease. CHIKV cause muscle necrosis as well as infiltration of inflammatory cells (126). Various other cell types are also affected viz monocytes and/or macrophages, dendritic cells (DCs), synovial and dermal fibroblasts, endothelial cells, muscle cells, periosteum, and sometimes keratinocytes (3,47,49,67,94,99,123). Widespread infection in these cells elicit strong immune response and is responsible for acute symptoms (129) (Fig. 1).
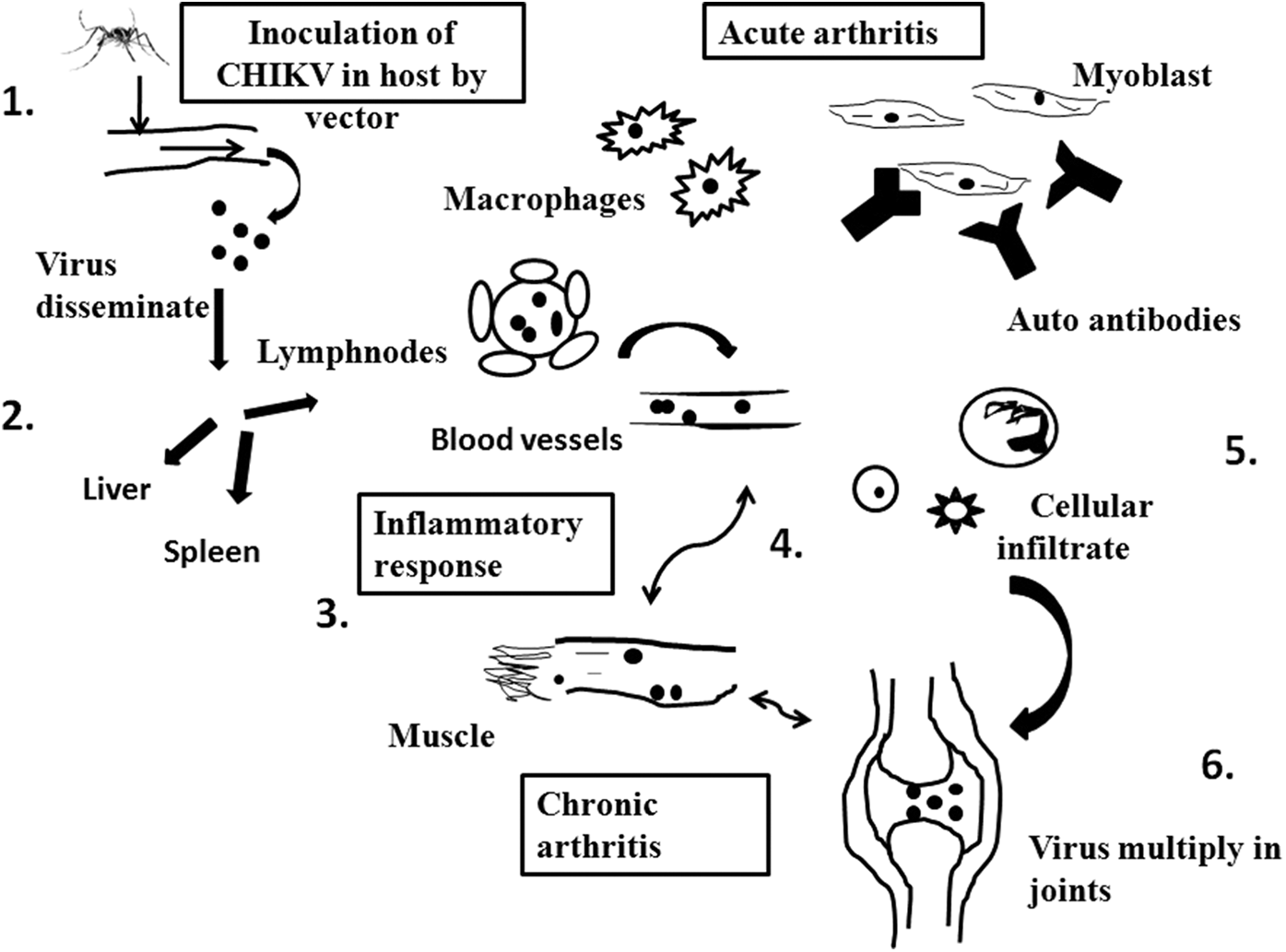
Figure showing the pathogenesis of Chikungunya virus. (1) Chikungunya virus inoculated by the bite of mosquito that is, Aedes albopictus. (2) Thus, viral particles disseminate through skin to blood stream. Then virus spreads to organs such as liver, spleen, and lymph nodes. (3) In acute phase of disease, virus tends to replicate in the target tissue which results in the infiltration of immune cells and macrophages. Proinflammatory cytokines expressed at the site of infection. These induce pain, which are responsible to cause arthralgia or arthritis. (4) In chronic condition, virus remains in target organs and cells. (5) The acute phase of the disease involves virus replication followed by an inflammatory response in the target tissues, which is characterized by an extensive infiltration of lymphocytes, NK cells, neutrophils, and macrophages (the main component). The increase in the levels of several proinflammatory cytokines and chemokines in the site of infection and in the plasma is associated with myositis and arthralgia/arthritis. Also, the secretion of metalloproteinases (MMP) in the joint tissue may contribute to articular damage. (6) Persistence of the symptoms may be related to the persistence of the virus or its products in the target cells with the subsequent accumulation of inflammatory mediators such as IL-6 and GM-CSF. NK, natural killer.
Acute symptoms of CHIKV infection starts with fever and body ache. Usually fever subsides in 10–15 days, but pain, stiffness, and swelling of the joints, especially in the lower limb around the ankles persist for months to several years (13,69). Polyarthralgia is an important manifestation of CHIKV fever (7) and is the most disabling symptom. Joint pain and inflammation mainly affect small joints in a symmetrical manner (fingers, wrists, and tarsus), but eventually occur in the large joints (knees and shoulders) as well. Arthralgias are generally of symmetric type with distal joints (hands, wrists, feet, and ankles) more commonly affected than proximal (53). Elbow, shoulder, knee, hip, temporomandibular, and sternocostoclavicular joints are also involved (22,130). CHIKV destroys collagen and hinders connective tissue metabolism in cartilage and joints, which leads to severe arthritis. This may also cause necrosis and fibrosis. During acute phase of infection, elevated levels of proline, hydroxyproline, and mucopolysaccharides are secreted in patient's urine, which is a biochemical evidence of cartilage damage (78).
Chronic phase is normally characterized by fluctuations in intensity and relapses affecting the same joint that were affected during the acute phase. Chronic phase is reported as both migratory, where in the pain spreads from one joint to another (56), and nonmigratory (53) and is generally less severe than the acute phase. CHIKV infection leads to restricted movement and quality of life. Such patients do not respond to commonly used analgesics, suggesting that the nature of chronic pain may be nociceptive and neuropathic (55).
Most reports published after the discovery of CHIKV were about the epidemiology and virus genotyping. There were very few reports on immune response and pathogenesis of CHIKV-induced arthritis. CHIKV-induced arthritis could be attributed to the host factors and/or critical mutations in the viral genome (131). The present review is focusssed on viral and host factors that are responsible for pathogenesis of CHIKV-induced arthritis.
Vector Factors
Vector competence that is, ability of a vector to acquire a pathogen and successfully transmit it to another susceptible host is influenced by some intrinsic (e.g., vector and viral genetics, vector competence, and vector life history traits) and extrinsic (e.g., temperature, rainfall, and human land use) factors that in turn influence the virus activity and mosquito biology in many ways (66). A. aegypti or A. albopictus both are invasive and responsible for the spread of CHIKV (80). Invasive species are of greater concern because they have potential effects on native species and ecosystems and/or human activities and health (59). Although A. aegypti is the primary classical vector for CHIKV, both the species are known to be susceptible to CHIKV infection. A. albopictus is quiet and diurnal and easily flourish in both urban and rural atmospheres. It's eggs are resistant to dry conditions. These are the possible reasons for A. albopictus being an important vector for CHIKV. CHIK epidemic in Reunion island in 2005–2006 was due to A. albopictus and identified as a chief vector in Europe, whereas A. aegypti was the important vector for carrying CHIKV in India (65). A. albopictus from La Reuion was found to be more efficient in disseminating CHIKV as compared with A. Albopictus from Vietnam (149). Although vector factors are known to affect the transmission of CHIKV, there is no direct correlation between the occurrence of CHIKV-induced arthritis and vector factors.
Viral Factors
Viral factors influencing infectivity, fitness of the virus, and its replication in vectors
Mutation in CHIKV genome contributes to its spread by mosquitoes. E1 is a class II viral fusion protein that mediates viral entry at low pH. It affects viral fusion, assembly, and/or cell tropism. During the 2005–06 La Reunion epidemic, an alanine to valine mutation was observed in E1 gene at the 226 position from the CHIKV isolates. Furthermore, studies suggested that A226 V mutation increased fitness of CHIKV in A. albopictus mosquitoes. CHIKV with A226 V mutation replicated to high titres in C6/36 cells depleted of cholesterol suggesting that adaptation of CHIKV to A. albopictus mosquitoes is associated with CHIKV dependence on cholesterol in the target cell membrane (146). However, further studies reported that CHIKV adaptation to A. albopictus mosquitoes is not related to cholesterol dependence or requirement of lower pH threshold for fusion reaction (148).
In the Malaysian outbreak during 2008, it was proved that A226 V mutation enhances transmissibility of CHIKV by A. albopictus mosquitoes (125). Wu et al. suggested a similar finding that CHIKV with an E1-A226 V was responsible for the 2010 outbreaks in China (155). Likewise, many mutations, such as M269 V, D284E, P294 L, S295F, A316 V, V322A, and C328 W, were observed in the membrane fusion glycoprotein E1 in isolates responsible for higher virulence (131).
A recent study has shown that nuclear localization sequence (NoLS) in the amino-terminal region of capsid protein is essential for replication and mutations in NoLS attenuates viral replication (142). Recently, some mutations identified in E2 glycoprotein also regulate CHIKV adaptation to its mosquito hosts (147). Although these mutations enhance the viral fitness, its replication and transmissibility by the vector, and their relevance to arthritis induced by CHIKV infection, remains unknown and needs to be answered in future studies.
Viral Factors Contributing to the Pathogenesis in Human Host
Viral factors are involved in the pathogenesis of CHIKV-induced arthritis. Viral load during infection might contribute to the development of arthritis. In a study conducted in cynomolgus macaques, inoculation of high doses of virus resulted in arthritis, meningoencephalitis, and death; intermediate dose led to development of fever; and at lower doses, clinical symptoms were not observed (70). Higher viral load in serum samples at the time of referral to clinic was associated with chronic status in elderly patients in a cohort of CHIKV-infected patients (53). These studies suggest that a higher viral load during acute stage of the disease might contribute to the development of chronic symptoms.
CHIKV survives in macrophages for a long time in nonhuman primates (70), indicating that CHIKV might establish chronic infections that evade immune responses. CHIKV can escape neutralizing antibody responses by undergoing genetic mutations that abolish antibody binding, or by indirect evasion strategies, such as cell-to-cell transmission (73). A recent study has reported that the pathogenic CHIKV strain (AF15561) was able to evade E2 domain neutralizing antibodies to establish persistence. A highly conserved glycine at position 82 in the A domain of the E2 glycoprotein impedes clearance and neutralization of multiple CHIKV strains. Attenuated CHIKV strain 181/25 was cleared from joint-associated tissues within 4 weeks of infection as compared with the pathogenic parental strain AF15561 (46). It is unclear how CHIKV exploited these mechanisms to persist in the host body.
In addition, CHIKV infection induces autophagy in host, which enhances the replication of viral RNA in the host (67). CHIKV activates endoplasmic reticulum stress and oxidative stress pathways to induce autophagy. Studies have suggested that many viral proteins are known to mimic host protein motifs and can interact with other host proteins to exploit the cellular infrastructure of the host (28). This is also seen in case of HIV and dengue virus infections (33,34). This phenomenon is also observed in nonviral diseases (29). Mapping the interaction between nonstructural viral proteins and cellular factors is important to understand how virus hijacks the cellular machinery of the host since such interactions are responsible for blocking the antiviral response to achieve genome replication and production of new virion particles (9).
Host Factors
Innate immune response
Innate immune response is not specific as it provides a first line of defense to the body. Monocytes, macrophages, natural killer (NK) cells, and DCs are the important mediators of innate immune response and hence they have been implicated in the immunopathogenesis of many viral diseases (26). These cells being located in the circulation and peripheral tissues are attractive targets for the virus and assists viruses in dissemination.
Monocytes/macrophages
Macrophages are the main infiltrating cell type in infected tissues post alphaviral infections. Macrophage and monocyte infiltration is commonly seen in CHIKV infection in mice, monkeys, and humans (41,53,70). They act both by protective and pathogenic pathways (79). Studies in experimental animals and patients showed that CHIKV infects monocytes/macrophages (41). Nakaya et al., have shown that upregulated expressions of genes associated with macrophage recruitment and activation in mice model during peak arthritis (96). Activated macrophages release a series of inflammatory mediators viz prostaglandins, tumor necrosis factor-α (TNF-α), and interleukin-6 (IL-6), which results in tissue damage, inflammation, and arthritis (136). CHIKV persists in monocytes through an apoptosis-assisted infection mechanism. CHIKV by hiding itself in apoptotic blebs can infect neighboring cells without triggering any antiviral response (58).
A study in macaques reported that CHIKV was detected even at 19 days postinfection in macrophages indicating viral persistence in these cells and might be responsible for the chronic symptoms observed in a percentage of human CHIKV cases (70). A study in mice has reported that CHIKV infection and the associated musculoskeletal inflammatory lesions activated M2 macrophages expressing higher levels of arginase 1 (Arg 1) enzyme. M2 macrophages are immunosuppressive in nature and are usually activated during wound healing response. Arg 1-expressing macrophages also suppressed T cell proliferation. Thus, it is possible that the immunosuppressive macrophages activated in musculoskeletal lesions might facilitate CHIKV persistence (134).
NK cells
NK cells respond to viral infection in early phase through interferon (IFN)-γ production, cytokine secretion, and cytolysis of infected cells (10). NK cells were shown to be highly activated in humans during CHIKV infection (53). NK cells provide protection against many viruses, including human cytomegalo virus, influenza virus, hepatitis viruses, and human immunodeficiency virus-1 (HIV). By secreting perforins and granzymes, NK cells mediate cytolysis (138).
The role of NK cells in CHIKV infection is still unexplored. Recent studies have reported that in an acute phase of CHIKV infection, NK cell repertoire is modulated to form a subset of NK cells that are unresponsive to cytokine stimulation, but with increased cytolytic ability. NK cells were also impaired in their ability to produce IFN-γ, expressed CD94/NKG2C, and inhibitory killer immunoglobulin-like receptors. Patients with higher viral loads showed prolonged persistence of NK cells expressing NKG2C and all developed chronic arthralgia suggesting a role for NK cells in chronic arthritis (108,109). In mice infected with CHIKV, NK cell infiltration was observed in arthritic feet (41).
Role of NK cells in rheumatic diseases is not clear, but NK cells activate the pathways leading to a range of proinflammatory cytokines (such as IFN-γ, receptor activator of nuclear factor kappa-β ligand, macrophage colony-stimulating factor) that may worsen RA pathology (1). Elsewhere, it is reported that higher levels of IL-15 (which stimulates NK cells) was observed in acutely infected CHIK patients (97). Thus, theoretically NK cells may exacerbate the disease (31). A recent study has reported a reduced frequency of perforin-positive NK cells and higher frequency of TNF-α-positive NK cells in chikungunya patients with chronic arthritis as well as in RA suggesting the possible utility of anti TNF-α therapy in CHIKV patients with arthritis (144).
Role of host proteins in CHIKV infection
Vimentin, actin, actin-related protein, myosin, and tropomyosin were among the differentially expressed cytoskeleton-associated proteins found in mouse model of CHIKV infection. Expression of vimentin was also observed on the surface of activated macrophages and apoptotic neutrophils. The probable reason for the increased expression of vimentin could be due to the increased neutrophil infiltration in CHIKV-infected muscles as observed in histological analysis (32). Four receptors of human host CD209, laminin receptor, and 60-kDa neural cell adhesion molecules have been identified to act as attachment receptors for CHIKV (76,124). Rana and group, suggested that five members of the 14-3-3 protein family interact with CHIKV nsP4 (114). They also discovered 15 cellular partners which interact with CHIK nsP3.
Adaptive immune response against CHIKV infection
Adaptive immune response against CHIKV infection is efficient and more specific to immune response (35). Elderly people have a weaker adaptive response and hence they are more prone to CHIKV infection and the disease become more severe in such patients (53). Both T cells and B cells play an important role in the immune response against CHIKV and in the pathogenesis of arthritis associated with CHIKV.
Role of B cells and antibody-mediated immune response
Antibodies are known to play an important role in the immune response against CHIKV. Anti-CHIKV IgM antibodies appear around 4th day postonset of infection and was detectable up to 6 months and mainly contribute to neutralization of the virus up to 10 days (23,25,60). In contrast, few studies have detected long-lasting CHIKV-specific IgM more than 12 months (53,84). The observation of long-lasting CHIKV-specific IgM antibodies in a minor proportion of patients suggests the persistence of viral antigens in sites which have no access to antibodies.
CHIKV-specific IgG antibodies appear around 4–20 days and take over the neutralizing activity and were observed even after 20 months (23,25). IgG3 was the predominant isotype of IgG antibodies and were directed mostly toward a particular epitope in the E2 protein termed as E2EP3 and are associated with neutralization activity (60,61). Patients with early IgG3 response (<10 days), although, had higher viremia and developed severe disease in the acute phase recovered fully, whereas patients with late IgG3 response (>10 days) had low viremia, but developed persistent arthralgia. This suggests that higher viremia induces an early IgG3 response leading to complete clearance of virus, whereas late IgG3 response results in incomplete clearance of virus leading to persistence of virus in hidden sites and hence persistent arthralgia (61).
A study from India has reported that earlier seroconversion to neutralizing IgG response was associated with better prognosis at the postacute phase (57). Hawman and group suggested that in persistently viremic mice that were deficient in B cells, injected antibodies could clear the CHIK viremia only, but CHIKV RNA still remained in tissues (45). Mice lacking B cells developed viremia that persisted until 1 year and exhibited increased CHIKV acute disease (81). This supports the role of B cells and neutralizing antibodies in viral clearance and prevention of persistent arthralgia.
Contrary to the protective role of IgG antibodies, few studies have reported an association between increased level of CHIK-specific IgG and long-term rheumatic manifestations (42,93). Autoantibodies were detected in a patient infected with CHIKV suggesting the association of CHIK virus with autoimmune diseases (82). In contrast to the above study, a longitudinal follow-up study for 36 months involving 180 patients could not identify any positive association between markers for autoimmune diseases and long-term arthralgia in CHIKV-infected patients (128).
Role of T cells
Role of T cells in the pathogenesis of CHIKV infection has been based on few clinical studies and investigations in mouse models. Investigation of CHIKV-infected patients in the acute phase revealed that T cells were elevated in numbers (53,152). CD8+ T cell response was observed in the earlier phase of the disease, whereas CD4+ T cell participation was observed during the late phase of virus infection (45). It was also shown that CD4+ and CD8+ T cells are infiltered in inflamed joints of mice infected with CHIKV (41). Teo and coworkers showed that CD4+ T cells, and not CD8+ T cells, were responsible for joint swelling and inflammation.
Cytokine and chemokine profiling during CHIKV infection suggested that CD4+ T cells could mediate inflammatory pathway through Th-17 and Th1-related mechanisms (143). Another study in IFN-γ knockout mice revealed a role for IFN-γ in CHIKV-induced arthritis (96). Thus, it is possible that CD4+ T cells might mediate joint inflammation through multiple mechanisms (143). A study carried out in C57BL/6J mice has reported that CHIKV infection led to an increase in the γδ T cells in the infected foot and draining lymph nodes and were associated with the production of proinflammatory cytokines and chemokines. Mice lacking γδ T cells showed exacerbated disease (79). Selective expansion of regulatory T cells (T regs) prior infection reduced CHIKV-induced joint pathology by induction of anergy in CHIKV-specific CD4+ effector T cells (74). Thus, the above studies provided the evidence for the role played by T cells in the pathogenesis of CHIKV-induced arthritis.
Role of osteoblasts in CHIKV-induced pathogenesis
Bone-forming osteoblasts (OB) and bone-resorbing osteoclasts (OC) are involved in maintaining the bone homeostasis. OBs express receptor activator of nuclear factor-κβ ligand (RANKL) and osteoprotegerin (OPG) that are involved in bone remodeling. Higher RANKL/OPG ratio favors the formation of osteoclasts from monocytic precursors. CHIKV infects OBs and is known to induce the production of IL-6 and RANKL and suppress OPG production. Hence, altered RANKL/OPG ratio leads to bone loss and arthralgia (98) (Fig. 2). Similar phenomena have been observed in RRV infection of OBs (18,19). CHIKV is also known to infect primary human fibroblast-like synoviocytes (HFLS) that are present in the synovial membrane lining the synovial joints. CHIKV infection of HFLS induces the production of RANKL,IL-6, IL-8, and monocytes chemotactic protein-1 (MCP-1; CCL-2) that recruit and induce the differentiation of monocytes into OCs (111). These studies suggest the role of interaction between the cells in the joints and bones and CHIKV in inducing arthralgia/arthritis.
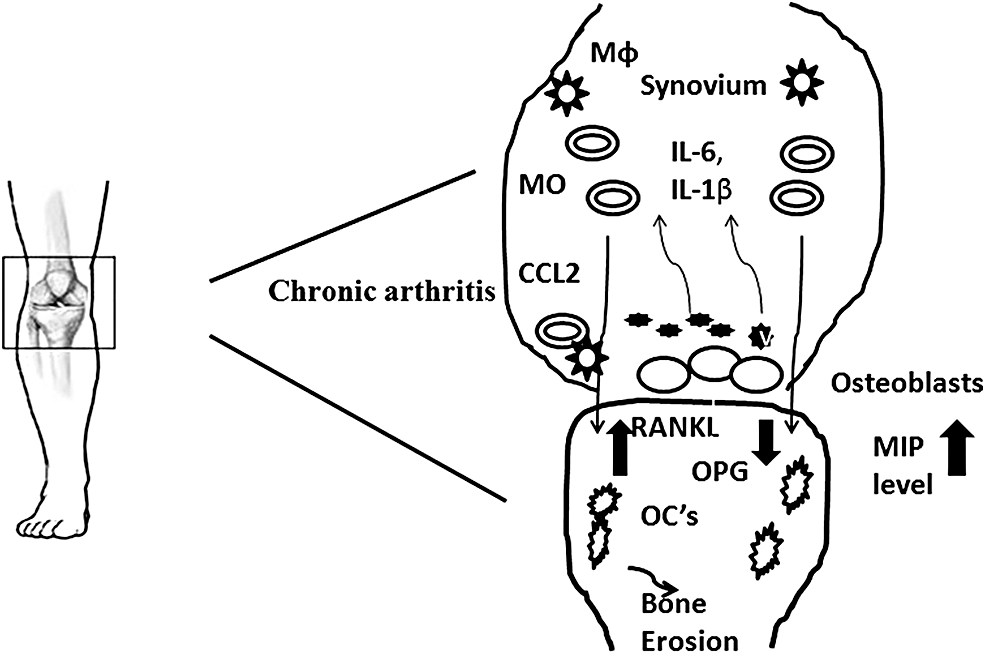
Figure showing the progression of CHIKV-induced arthritis. Virus attacked OB cells and secrete IL-6, IL-1β, CCL-2 (MCP-1), and MIPs. Monocytes and macrophages are stimulated by chemoattractive gradients such as CCL-2. Increase in RANKL/OPG ratio, osteoclast cells are produced, leading to the damage to bone and hence developing arthritis. CHIKV, chikungunya virus; MIP, macrophage inflammatory proteins; OB, osteoblast.
Cytokines and chemokines in CHIKV infection
Cytokines/chemokines play a vital role in viral immunopathology.
Type I interferons
Type I IFNs represent one of the most important antiviral responses. IFNs are capable of limiting viral replication/dissemination, or they may induce potent host immune response by regulating proinflammatory cytokines (153). In vitro studies have shown that infected nonhematopoietic cells sense viral RNA in a Cardif-dependent manner and control infection by producing type I IFNs (127). Levels of IFN-α were found to be elevated in patients with higher viral load compared with those with lower viral load suggesting that higher viral load induces IFN-α production (24). Elsewhere, it is also reported that during acute phase of CHIKV infection, type I IFN expression was upregulated in sera and tissue samples from patients (24,152). Similar findings were observed in primates, mice, and zebrafish (89,101,120). Moreover, it was observed that IFN-α/β, along with antiviral antibodies, protected mice against CHIK infection and severity of disease (41,129,137).
Studies conducted on chikungunya-infected patient cohorts also suggested higher expressions of IFN in sera and inflamed tissues during the inflammatory phase of CHIK infection (21). All these studies suggest that type I IFN is an important biomarker, which is upregulated during CHIKV infection. NK cells produces large amount of IFN-γ, which helps in the maturation of other immune cells. They also activate T helper cells and were shown to possess potent antiviral activities (139).
It is suggested that the cytokine profile in CHIKV infection is similar to those secreted during arthritis (63,92). Studies have suggested that IL-1β, IL-6, and Regulated on Activation in Normal T Cell Expressed and Secreted (RANTES) were linked with the acute phase of CHIKF (63). IL-1β and IL-6 account for the high-grade fever in CHIK (97). In addition to this, high expression of IL-6 has also been implicated in bone loss in other bone disorders such as RA (75) and bone metastasis (157). Patil et al. showed that levels of proinflammatory cytokines of Th1 group were found to be increased in a mouse model of CHIKV (106). Similar findings were also reported by many researchers (18,21,24,63,97).
Interferon gamma-induced protein 10 (IP-10), and macrophage inflammatory proteins (MIP) had increased plasma levels during CHIKV infection (53). These chemokine signals through the same chemokine receptor (CXCR3) may activate and direct the migration of T cell subset (113). Elevated levels of CCL-2, IL-6, and IL-8 have been reported in the synovial fluid of CHIKV-infected patients. Monocyte chemoattractant protein (MCP)-1, MIP-1α, and MIP-1β were increased during the chronic phase of CHIKV infection (107). Higher levels of MCP-1 have been observed in patients with CHIKV and RRV-induced arthritis. MCPs play an important role in recruiting the inflammatory cells to injured tissue (3). MCPs also play a pathogenic role in mediating arthritis and myositis (121). MCPs may also cause exacerbated bone loss. RANTES regulates protective immunity to viral infections. Lower levels of RANTES were observed in chronic phase of CHIKV infection (14,63). These chemokines induce direct movement of blood monocytes or macrophages. CHIKV infection of OBs increases the ratio of RANKL/OPG that is responsible for differentiation of monocytes into bone-resorbing osteoclast (OC) cells contributing to arthritis (19).
Host genetics and chikungunya-induced arthritis
Two studies from India have reported the association of human leukocyte antigen (HLA) alleles with chikungunya infection. A study from western India has reported the association of HLA-DRB1*11 and HLA-DRB1*11-HLA-DQB1*03 haplotype with resistance to chikungunya infection and HLA-DRB1*04-HLA-DQB1*03 haplotype with susceptibility to chikungunya infection (144). A study from Andaman islands reported a lower frequency of HLA-DQB1*03:03 allele and HLA-DQB1 alleles with glutamic acid at the 86th position of peptide-binding pocket in CHIKV-infected patients suggesting their influence on infection and pathogenesis (14). The same group from Andaman islands also reported that single nucleotide polymorphisms (SNPs) in OAS gene cluster that codes for various oligoadenylate synthetases, an important part of innate immune response, influenced the risk of developing clinical symptoms in CHIKV-infected patients (15).
A study on CHIKV-infected patients belonging to multiple ethnicities reported that a noncoding SNP in the TLR3 gene, rs6552950, is associated with disease severity and CHIKV-specific neutralizing antibody response (50). However, none of study has looked at the association of host genetic factors with persistence of arthritis for longer periods in CHIKV-infected patients, except for one. A study from Reunion island reported that 66.6% of CHIKV-infected patients with RA had HLA-DRB1*04 or HLA-DRB1*01 alleles. However, this study lacked appropriate controls to conclude the association between arthritis in CHIKV-infected patients and HLA alleles (8).
Various studies carried out in different populations have reported the association of HLA-DRB1*04, HLA-DRB1*01, and HLA-DRB1*10 alleles with susceptibility to RA (6,71,86,156). Moreover, it has been shown that HLA-DRB1 alleles with common amino acid pattern, RAA, also called shared epitope (SE), in positions 72–74 of the third hypervariable region of the DRB1 chain was reported to be associated with RA susceptibility in different populations. This association of HLA-DRB1 alleles coding for SE with RA is further influenced by the presence of aminoacids in the positions 70 & 71 (4,43). A recent study has reported that SNPs in the genes coding for RANK/RANKL/OPG were associated with the presence of anticitrullinated antibodies and erosion in RA (122). It is possible that in subjects with susceptibility alleles for RA, CHIKV infection acts as a trigger for the development of arthritis or CHIKV, independent of these susceptibility factors, might induce arthritis. Furthermore, studies are needed in CHIKV-infected patient cohorts with and without persistent arthritis. This could lead to the identification of genetic markers that predispose to arthritis in CHIKV-infected patients.
Preventive and Therapeutic Approaches Against CHIKV Infection and CHIKV-Induced Arthritis
Therapeutic approaches
Nonsteroidal Anti-Inflammatory Drugs (NSAIDs), pain killers, and corticosteroids are important drugs of choice against arthritogenic viruses, including CHIKV (137,104). These treatments are opted based on symptoms, although these often provide only partial relief (135), as neither effective licensed vaccine nor specific anti-CHIKV therapy and antiviral drugs are available in the market (5,48,145). Methotrexate, sulfasalazine, and paracetamol are used frequently to alleviate the symptoms (40). However, these drugs cause serious side effects. Ribavirin and chloroquine have been reported to have useful relieving effects during CHIK fever (2,100,115). A mixture of Methotrexate and hydroxychloroquine was effective against CHIKV-induced arthritis (102). Chloroquine phosphate significantly improved Ritchie articular index and morning stiffness in an open pilot study with 10 patients (12).
Anti-IL6 therapy, TNF antagonist, anti-RANKL therapy, and direct targeting of Th 17 cells and cytokines have been proposed as potential immunotherapies against Alphavirus-induced arthritis (20). Agents such as pentosan polysulfate and bindarit, an inhibitor of MCP-1, have shown promising results in mice models in reducing Alphavirus-induced arthritis and bone loss (20,52).
Along with these traditional and chemotherapeutic agents, some other alternatives have also been reported. RNA interference (RNAi)-mediated inhibition of CHIKV has been suggested by some groups (27,103). These studies suggested that siRNA's targeted against the nsP3, E1, and nsP1, E2 genes, respectively, inhibited the replication of CHIKV in mammalian cells. Likewise, plasmid-based small hairpin RNA's (shRNA's) have also shown promising results in silencing replication of CHIKV in mammalian cell lines. CHIKV replication inhibition was reported in suckling mice pretreated with shRNA E1 (72).
In another approach, inhibiting the Manose-binding lectins (MBL) pathway in mice prevented the Ross River virus-induced arthritis/myositis (44). Inhibitory agents, which target the MBL pathway, may be helpful in the treatment of CHIK-induced arhtirits. RNA seq analysis during acute and chronic arthritis in a CHIKV-infected adult mouse model identified granzyme A, present in cytolytic T cells, as an important component of arthritis inflammation. Further studies revealed that granzyme A knockout mice infected with CHIKV showed pronounced reduction in arthritis. Moreover, inhibition of granzyme A in CHIKV-infected wild-type mice also reduced arthritic inflammation. These results suggested that targeting granzyme A could be beneficial in treating CHIKV-induced arthritis (154).
Gene profiling/gene set enrichment analysis showed that there are close similarities in the differentially expressed genes in the CHIKV arthritis model and in RA. Thus, new drug regimens being developed for RA might also be useful in CHIKV-induced arthritis (96). A recent study investigated the effect of eight food and drug administration (FDA)-approved antirheumatic drugs on joint swelling in CHIKV-infected mice and identified that a monoclonal antibody (MAB) against cytotoxic T lymphocyte antigen 4 (CTLA4) termed as abatacept and a janus kinase inhibitor, tofacitinib, reduced joint swelling without affecting viral RNA levels. When abatacept was given in combination with an anti-CHIKV MAB (4 N12), joint swelling was completely resolved with partial reduction in viral RNA levels (90). This study suggested that targeting T cell activation might form a potential therapy against CHIKV-induced arthritis.
Preventive approaches
Many CHIKV vaccine candidates have been reported, which includes inactivated virus formulations (41), live attenuated virus vaccines (105), chimeric virus vaccines (150), DNA vaccines (83), T cell-based peptide vaccine (140), a recombinant adenovirus vaccine (151), subunit protein vaccines (68), and a virus-like particle (VLP) formulation (64). Chang et al. have tried Chikungunya VLP vaccine in an open-label clinical trial. This trial has shown satisfactory results with regard to safety, tolerance, and immunogenicity (17). An insect-specific Alphavirus, Eilat virus (EILV) cDNA, has been utilized as a vaccine platform to express CHIKV structural proteins and was found to elicit robust immunity against CHIK in mouse and non-human primate models (38).
Considering these facts, mosquito control and public health response measures are very important to check the spread of the disease. Traditional mosquito repellents or insecticides are costly on mass scale and labor intensive as well (91). Resistance against a particular insecticide minimizes its use in vector control. Transgenic A. aegypti engineered with a late-acting lethal genetic system is a novel approach for vector control (110). Wolbachia as a novel biological control method is of interest. Mosquitoes infected with intracellular bacterium Wolbachia are resistant to infection. Surveillance and early detection of CHIKV outbreaks are important steps to stop the transmission cycle. Avoiding mosquito bites by wearing protective clothing, and maintaining the hygiene and cleanliness are advisable to limit human contact with vector and further spread of CHIK.
Conclusion
CHIK virus is currently generating an epidemic of chronic arthritis worldwide. In recent years, there have been explosive outbreaks of CHIK fever in several parts of the world and is responsible for morbidity in a very short time period. CHIKV-induced arthritis resemble RA in various aspects. Available knowledge indicates that immunosuppressive macrophages, IgG3 antibodies, activation of CD4+ T cells and associated inflammatory response, and possible persistence of virus in hidden sites contribute to the development of CHIKV-induced arthritis. Studies in mouse models have revealed that novel drug regimens/MABs used for the treatment of RA or drugs/MABs used in reducing inflammation in combination with anti-CHIKV MABs are effective in treating CHIKV-induced arthritis, and there is urgent need for clinical trials in humans. Thorough understanding of CHIKV-induced arthritis might pave the way for identifying biomarkers to predict arthritis in CHIKV-infected patients and to develop efficient new drugs to treat CHIKV-induced arthritis.
Footnotes
Acknowledgment
Sarika Amdekar would like to acknowledge Science and Engineering Research Board (SERB) for providing the research grant (SERC/LS-58/2012).
Author Disclosure Statement
The authors declare that there are no conflicts of interest regarding the publication of this article.