
Abstract
The objective of this study was to develop a new antigen delivery system using an alphavirus replicon particle (RP) to induce humoral antibody responses against bovine viral diarrhea virus (BVDV) recombinant antigen produced from envelope glycoprotein E2. An alphavirus RP expressing the E2 glycoprotein of BVDV was used for immunization of pigs. A fluorescent microsphere immunoassay (FMIA) has been applied to detect BVDV E2 antigen-specific antibody isotype in pig immunized with alphavirus RP. Full-length BVDV E2 (aa 1–375) was cleaved into several pieces, eight E2 DNA fragments, including full-length DNA, were cloned into expression vector pHUE, and the recombinant proteins expressed in BL-21 (DE3) Escherichia coli. After successful conjugation of purified proteins with microsphere beads, a multiplex FMIA platform was constructed, and BVDV E2 alphavirus-based RP-immunized animal serum samples were tested in the presence of bead-bound antigen targets. The results were represented as mean fluorescence intensity (MFI); the MFI values were converted to sample value/positive value (S/P) ratios. BVDV E2 (aa 1–183) showed the highest MFI values of eight recombinant E2 fragments when the specific activity of each fragment was tested. In immunized animals, data for BVDV E2-specific IgA, IgG, and IgM in serum and only IgG and IgA in oral fluids were recorded. The MFI values for the positive serum sample showed a 100-fold increase compared with the negative serum sample. Antibody isotype to BVDV E2 antigens showed that IgG > IgM > IgA in serum, whereas IgG > IgA > IgM in oral fluids. The data presented in this study suggested that boosting with the same doses of alphavirus RP in 3-week intervals may potentially enhance antibody response. The experimental results demonstrate that alphavirus RP-expressing BVDV E2 antigen induces antibody response in pig.
Introduction
B
Research has developed a variety of diagnostic strategies that are useful in the control of BVDV infection. These strategies include multiple different diagnostic tests to identify persistently infected (PI) cattle, develop vaccine to maximize immunity, and ensure biosecurity to prevent additions of PI animals. Detection and elimination of animals PI with BVDV are essential for the control of BVD and eradication of BVDV. There are several immunological diagnostic assays for detection of BVDV; however, a substantial improvement in sensitivity, specificity, and affordability of these assays is required.
A variety of different diagnostic tests for routine diagnosis of BVDV have been developed, including virus isolation, enzyme-linked immunosorbent assay (ELISA), immunohistochemistry of skin, serum virus neutralization test, indirect fluorescence antibody test, and polymerase chain reaction for serum (8,19,27). The virus neutralization test (VNT) is the gold standard for antibody detection against BVDV (9). ELISA-based assays are alternatives to gold standard techniques, such as the VNT and virus isolation. Additionally, antibody detection techniques have advantages for the early detection of viral infection (28). However, ELISA has long since been the most common technique for detection of antibody against E2 and Erns of BVDV (6,16). E2 and Erns are glycoproteins found on the surface of the virion. A 53 kDa major structural glycoprotein is encoded by the BVDV E2 gene that might have a majority of neutralizing epitopes (15). E2 is immunodominant in eliciting antibody response to virus infection, whereas Erns plays an important role in immune response.
Currently, recombinant or vectored vaccines are not commercially available. To reduce catastrophe caused by BVD in infected cattle, vaccination with modified live vaccine or inactivated vaccine is the approach used by producers (25). Recently, an alphavirus replicon particle (RP) expressing the E2 glycoprotein of BVDV in cattle was developed to evaluate BVDV-neutralizing activity (22). In previous studies, a replicon-based expression system was generated from the alphavirus Venezuelan equine encephalitis (VEE) (7,24). VEE RP has been tested against foot-and-mouth disease virus (FMDV) in cattle and potential protection was reported (26).
Recently, a fluorescent microsphere immunoassay (FMIA) or Luminex® has been developed, which can be used for detection of antibody response to several antigen targets in a single platform (13). In other studies, an alternative secondary conjugate-based FMIA has been successfully utilized to detect porcine reproductive and respiratory syndrome virus and porcine circovirus type 2 (PCV2)-specific IgG antibodies (12). In our previous studies, we established a multiplex FMIA for detection of CSFV-specific IgA, IgG, and IgM antibodies in pig immunized with alphavirus-expressed CSFV E2 antigens (11). In the present study, an alphavirus RP expressing the BVDV E2 antigen-induced antibody response and detection of BVDV E2 antigen-specific antibody were tested using FMIA.
Materials and Methods
Oral fluid and serum samples after immunization
Ten weanling, large, white, and Landrace crossbred domestic pigs, 3 to 4 weeks of age, were immunized with RP expressing the E2 glycoprotein of BVDV. Pigs were maintained in the Large Animal Research Center (LARC) BSL2 facility at Kansas State University. Animal care procedures were followed according to the Guide for the Care and use of Agricultural Animals in Research and Teaching by the Federation of Animal Science Society (FASS), as described previously (12). Use of alphavirus RPs and animals was approved by the Kansas State University Institutional Animal Care and Use Committee and Institutional Biosafety Committees.
The E2 glycoprotein gene from BVDV subtype 1b was cloned into a replicon vector plasmid, as previously described (17). A replicon-based expression system was created from the alphavirus VEE (24). RPs were produced in vero cells by coelectroporation of E2 replicon RNA and structural gene helper RNAs and the particles were harvested from vero cell culture (10). The pigs were acclimated before the onset of the trial. Animals were injected intramuscularly with 2 mL/animal of 2.5 × 108 infectious units/mL RPs (11). At days 21 and 42 postimmunization, animals were boosted with same doses of RPs. Whole blood was collected in tubes containing EDTA on days 0, 7, 14, 21, 28, 35, 42, and 49 after immunization and processed for serum isolation. Serum was separated by centrifugation and stored at −80°C until use. Oral fluids were collected from a group of pigs (pens) at 7-day intervals for up to 49 days postimmunization (dpi).
Recombinant BVDV E2 protein expression and purification
For the antigen targets in FMIA, eight recombinant antigen fragments were generated from E2 glycoprotein of BVDV subtype 1b. Whole E2 (1–375 aa) was fragmented into seven small pieces (aa 1–183; aa 91–274; aa 184–375; aa 1–90; aa 91–183; aa 184–274; and aa 275–375) (Figs. 1 and 2). Eight E2 DNA fragments, including full-length DNA, were cloned into expression vector pHUE (4), the recombinant proteins expressed in BL-21 (DE3) Escherichia coli, and purified by methods as described elsewhere (12).
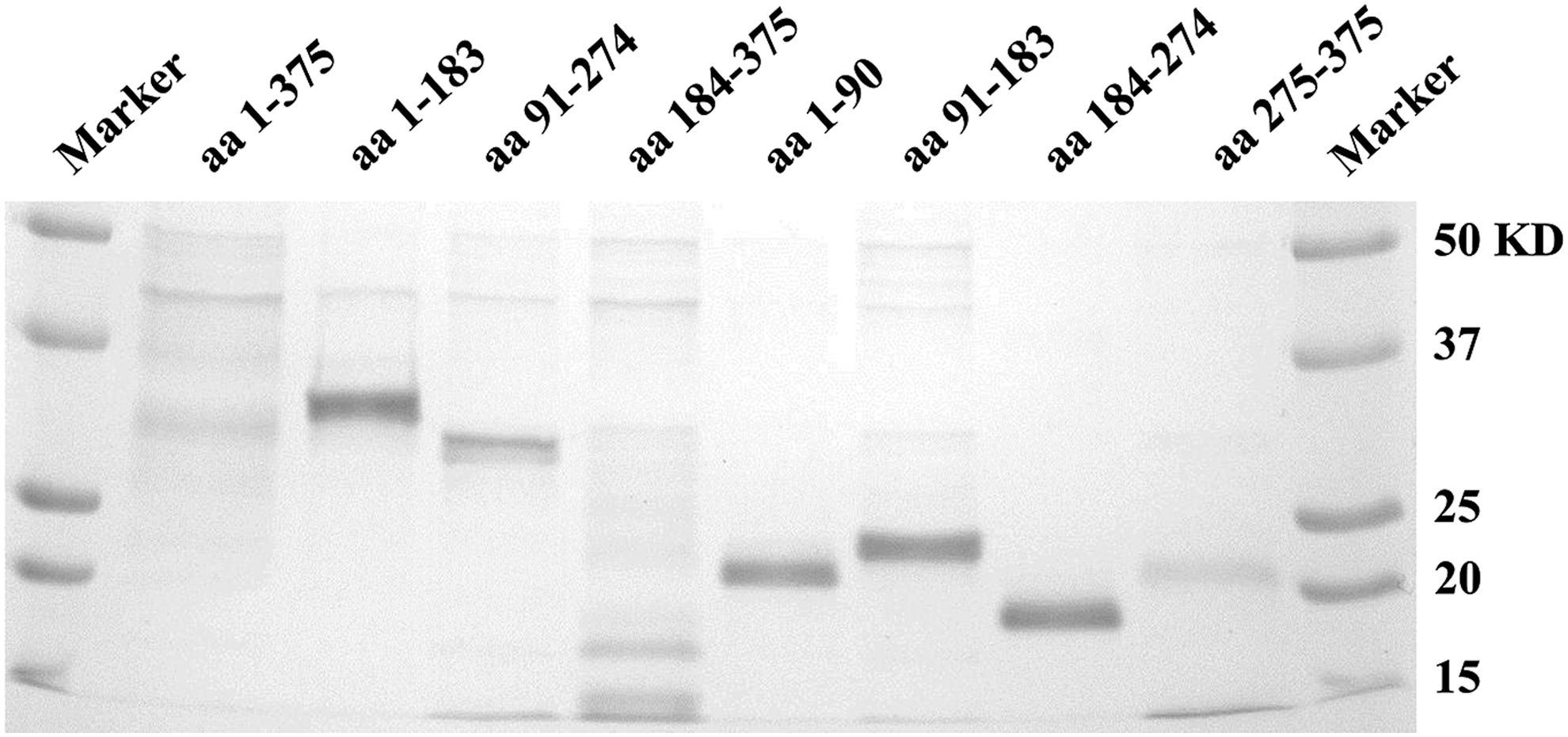
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis of recombinant bovine viral diarrhea virus (BVDV) E2 proteins after Coomassie blue staining. The right lane shows the molecular mass standard; the remaining lanes represent BVDV E2 recombinant protein fragments.
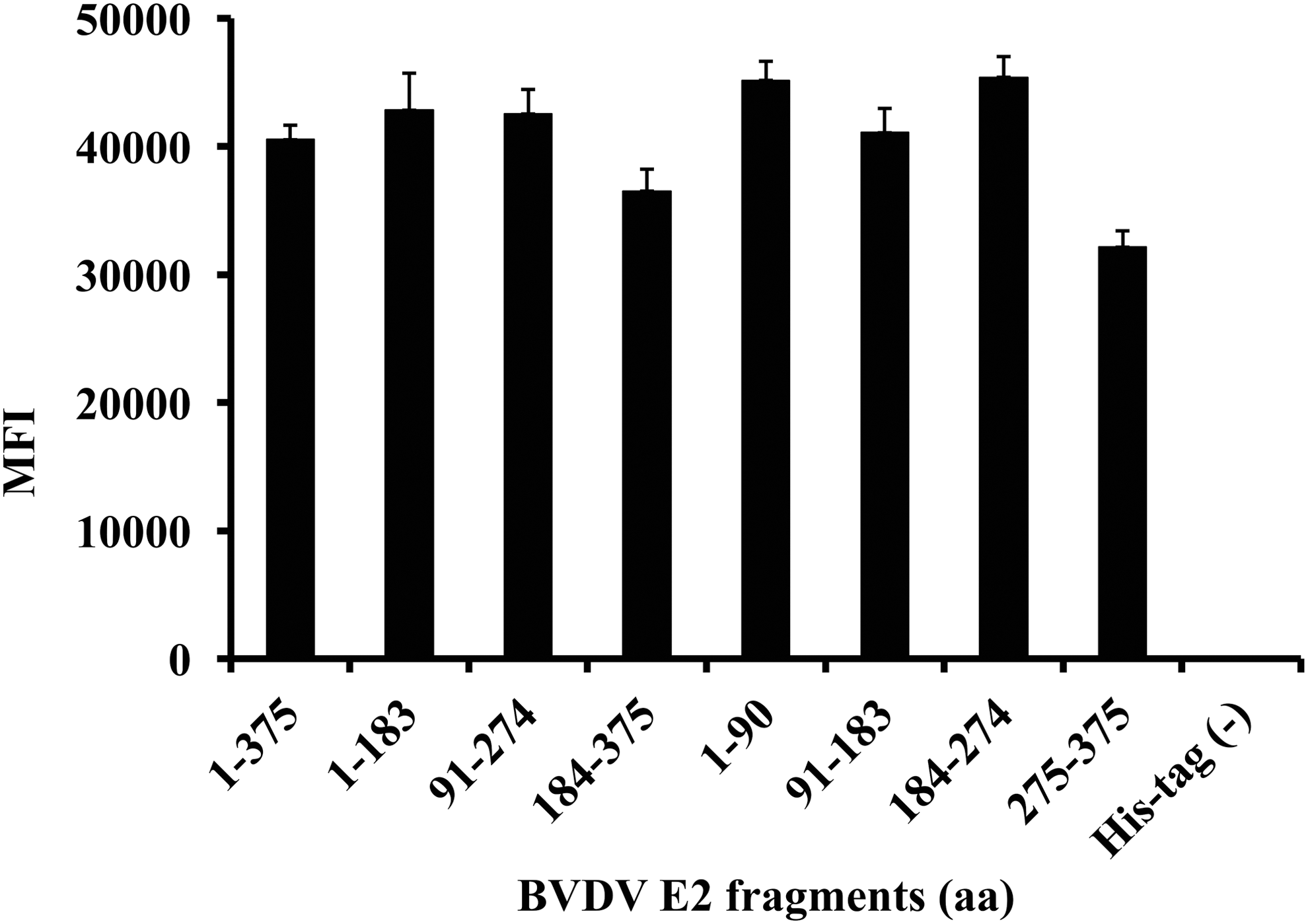
Binding activity of anti-5xhistidine antibody against recombinant BVDV E2 proteins. Anti-His™ mouse monoclonal antibody was diluted 1:100 in sample buffer and fluorescent microsphere immunoassay (FMIA) was performed in the presence of BVDV E2 protein-conjugated microsphere beads. Results are reported as mean fluorescence intensity (MFI) and standard errors.
Protein expression and purification
Expression and purification methods were followed as described previously (12). In brief, for the expression of recombinant proteins, bacteria were grown in LB medium with ampicillin at 37°C. When the OD600 reached 0.4–0.6, the expression of the gene was induced by adding isopropyl β-D-1-thiogalacto-pyranoside (IPTG) to a final concentration of 1.0 μg/mL with an additional 4 h of incubation in the culture with shaking. The culture was centrifuged at 4,000 g for 10 min at 4°C, and the bacterial pellet was subjected to three freeze–thaw cycles and resuspended in 5 mL of 1× LEW buffer (50 mM sodium phosphate monobasic, 300 mM sodium chloride, pH 7.0). Bacteria were lysed with the addition of 1 μL/mL of EDTA-free protease inhibitor cocktail (Thermo Scientific, Worcester, MA) and 1 mg/mL of lysozyme. After incubation on ice for 30 min with shaking, sonication was performed using the Model 3000 Ultrasonic Homogenizer (Biologics, Inc.) for 30 sec on ice. The bacterial lysate was then centrifuged at 20,000 g for 30 min at 4°C. The supernatant was filtered through a 0.45-μm PVDF filter (Fisher Scientific, Pittsburgh, PA) to eliminate any debris. An affinity chromatography was performed with the addition of the filtered lysate to a PrepEase His-tagged protein purification high-yield kit (Affymetrix, USB, Santa Clara, CA). The His-tagged protein-bound column was washed three times with 1× LEW buffer, and the protein was eluted three times with 1× elution buffer (50 mM sodium dihydrogen phosphate, 300 mM sodium chloride, and 250 mM imidazole, pH 8.0). BIO-RAD protein assay reagent (BioRad, Hercules, CA) was used for measuring the concentration of protein using a precision microplate reader (Molecular Devices, Sunnyvale, CA) and bovine serum albumin (Sigma-Aldrich, St. Louis, MO) was used as the protein standard. The purity of protein was analyzed by sodium dodecyl sulfate (SDS)–polyacrylamide gel (10%) electrophoresis (PAGE) as described previously (18) and gels were stained with Coomassie blue.
Preparation of MARC-145 cell lysates as a source of His-Tag (−) antigen
Antigens from virus-infected cells were prepared as described previously (14). Bovine turbinate (BT) cells were maintained in minimum essential medium with 8% fetal bovine serum and 1× penicillin/streptomycin and fungizone (Invitrogen, Grand Island, NY). BT cells were infected with BVDV-1a Singer strain virus, culture supernatants were concentrated, and then viral pellets were lysed in 1% Triton X-100. Viral lysates was used as His-tag (−) antigens in FMIA.
Conjugation of recombinant antigens to microsphere beads
Recombinant proteins were covalently coupled to Luminex MagPlex® polystyrene, carboxylated magnetic microsphere beads (Luminex Corporation, Austin, TX) using the method described previously (13). Briefly, 500 μL of beads at a concentration of 1.25 × 107 beads/mL was placed into a low protein binding microfuge tube (USA Scientific, Inc., Ocala, FL). For all subsequent procedures, the beads were protected from prolonged exposure to light. The beads were removed from suspension by attachment to a magnetic separator and washed in 100 μL of sterile water, followed by gentle vortexing and sonication for 20 sec to disperse any aggregated beads. The beads were magnetically separated and resuspended in 80 μL of activation buffer (0.1 M sodium dihydrogen phosphate pH 6.2). After vortexing and sonication, 10 μL of 50 mg/mL sulfo-NHS (N-hydroxysulfosuccinimide) and 10 μL of 50 mg/mL 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide hydrochloride, EDC (Thermo Scientific, Rockford, IL), were added. Beads were incubated for 20 min at room temperature. The activated beads were magnetically separated and washed twice with 250 μL of PBS and resuspended in 100 μL coupling buffer, followed by the addition of 25 μg of recombinant protein. The final volume was brought up to 500 μL in PBS and the reaction mixture gently rotated for 2 h at room temperature. The coupled beads were washed three times in PBS with 0.05% Tween-20 and 0.05% w/v sodium azide (PBST) and resuspended in 1 mL of PBST with 1% fish gelatin (Sigma-Aldrich) and stored at 4°C in the dark until use.
Bead-based microsphere immunoassay
For the multiplex FMIA, eight BVDV E2 recombinant antigen-conjugated bead sets and an unconjugated bead set as a background control were combined and mixed well. A penta-His monoclonal antibody was used for detection of relative amounts of recombinant protein bound to each bead set, as described previously (11). To normalize the number of beads in the reaction, beads were counted under a microscope using a hemocytometer. A fifty-microliter assay buffer (PBS with 4% goat serum) containing ∼1,250 antigen-coupled beads and 50 μL of twofold diluted penta-His™ mouse monoclonal IgG (Qiagen GmbH, Valencia, CA) or diluted serum samples were added in each well of a Corning™Costar™ polystyrene, white, round-bottom 96-well plate (Corning, Inc., Corning, NY). The plates were covered, placed on a shaker, and incubated for 30 min at room temperature. The plate was placed in a magnetic plate separator (Luminex Corp.) for 1 min and the supernatant was dumped by rapid forceful inversion two to three times and 190 μL assay buffer was added, and this step was repeated three times. Biotinylated anti-mouse IgG (H+L) (2 μg/mL) (Vector Laboratories, Inc., Burlingame, CA), biotin-SP-conjugated goat anti-swine IgG (2 μg/mL) (Jackson ImmunoResearch Laboratories, Inc., West Grove, PA), biotin-labeled goat anti-swine IgM (μ) (KPL, Gaithersburg, MD), and goat anti-pig IgA H&L (Biotin; Abcam, Inc., Cambridge, MA) were diluted in assay buffer. With the beads, fifty microliter of diluted biotinylated antibody was incubated for 30 min. Fifty microliter of streptavidin-conjugated phycoerythrin (SA-PE; Moss, Inc., Hadley, MA), diluted 1:500 in assay buffer, was added to the beads after three washes and incubated for 30 min at room temperature. The beads were resuspended in 100 μL of PBS-GS after washing three times, and the plate was read. Data were analyzed using xPONENT 4.2 software, and results were reported as mean fluorescence intensity (MFI). The MFI for 100 coupled microspheres for each individual target was calculated. The MFI was calculated from the median value for at least 100 beads. The results were reported as MFI of the test sample minus MFI of the background bead set (11). To determine the presence of antibody to BVDV E2 in swine, the sample value/positive value (S/P) ratios were calculated using the following equation S/P = (Mean MFI of test sample − Mean MFI of negative control)/(Mean MFI of positive control − Mean MFI of negative control).
Results
Expression of BVDV E2 recombinant proteins
To incorporate antigen targets in FMIA, recombinant antigens were initially expressed as His-tagged fusion proteins in E. coli. The purity of the recombinant protein was tested using SDS-PAGE. As shown in Figure 1, Coomassie blue staining of recombinant proteins in SDS-PAGE was identified as a major band of BVDV E2 fragments. All of the His-tagged recombinant proteins possessed the predicted size and migrated accordingly; the molecular masses of each band were recorded as 50 KD for full length (aa 1–375), 28 KD for aa 1–183, 28 KD for aa 91–274, 29.5 KD for aa 184–375, 18 KD for aa 1–90, 18 KD for aa 91–183, 18 KD for aa 184–274, and 19.5 KD for aa 274–375. In addition to protein purity, the identity of each bead set was confirmed by interaction of penta-His mAb and protein attached to each bead set. All of the bead sets were positive for binding the penta-His mAb (Fig. 2). The penta-His mAb showed similar binding activities with all of the eight recombinant proteins on the surface of the beads, even though full-length (aa 1–375) and aa 275–375 protein bands were weak, whereas BVDV lysates as a His-tag-negative control did not bind with penta-His mAb. Overall, the results show that anti-His tag antibody is a useful tool for the analysis of His-tag recombinant proteins in FMIA.
Multiplex detection of BVDV E2 IgG antibodies in alphavirus RP-immunized animals
To determine the IgG antibody response to BVDV E2 antigens, a multiplex FMIA was performed using BVDV E2 recombinant antigen-coupled beads in the presence of serum sample. Of the eight E2 antigen fragments, aa 1–183 showed the highest IgG response (Fig. 3). In this study, IgG responses to BVDV E2 (aa 1–183) in 10 animals immunized with alphavirus RP are represented in Figure 4. The FMIA was performed using (aa 1–183) antigen-coupled beads to investigate the kinetics of antibody responses in alphavirus RP-immunized animals. Serum samples used for this study were obtained from immunized animals on days 0, 7, 14, 21, 28, 35, 42, and 49. All of the serum samples were highly reactive to BVDV E2 (aa 1–183). The results showed that levels of IgG were undetectable by 21 dpi, peaked at 28 dpi after first boosting on day 21, and remained stable through 35 dpi. Further analysis showed that S/P ratios were decreased on 42 dpi and increased after the second boosting on day 42, suggesting that same doses of alphavirus RP after boosting at day 42 increased antibody responses. MFI values for the positive serum sample showed 100-fold increase on day 28 compared with the negative sample (Fig. 3).
Multiplex detection of BVDV E2 recombinant antigen-specific IgG responses in serum samples from animals immunized with alphavirus replicon particles (RPs). BVDV E2 recombinant proteins were conjugated to microsphere beads and FMIA was performed in the presence of 1:100 diluted serum samples of alphavirus RP-expressed BVDV E2-immunized animals. Data represent MFI and ± standard deviation of triplicate determinations.
Multiplex detection of BVDV E2 recombinant fragment-specific IgG responses in serum samples from animals immunized with alphavirus RPs. FMIA was performed in the presence of 1:100 diluted serum samples of immunized animals and BVDV E2 protein-conjugated beads. Data represent sample value/positive value (S/P) ratios and the S/P ratios were calculated from MFI values.
BVDV E2-specific IgA and IgM in alphavirus RP-immunized animal serum
To determine changes in BVDV E2-specific IgA during the course of the study, 10 animals were immunized with alphavirus RP, and serum samples were collected on days 0, 7, 14, 21, 28, 35, 42, and 49. IgA response against BVDV E2 (aa 1–183) was evaluated in serum samples by FMIA and the reactivity of antibody was recorded over time, as shown in Figure 5. BVDV E2 (aa 1–183) antigen-specific IgA was detected in 3 of 10 animals. Of the three, pig 11 showed a high level of IgA at 7 dpi, followed by undetectable level on days 14 through 21, and after first boosting, the level was elevated upto 50% of highest level (Fig. 5). Moreover, analysis showed that IgA level decreased to 0.5 S/P ratio on day 35, and after second boosting, it reached peak level on day 49. Similarly, IgM response to BVDV E2 (aa 1–183) antigen in 10 animals immunized with alphavirus RP was evaluated using FMIA. The results showed that 3 of 10 serum samples were positive to BVDV (aa 1–183) antigen in FMIA. All of the three pigs showed peak level of IgA on days 7 through 14, followed by 0.5 S/P ratio level on day 21, and after the first boosting, IgM level elevated upto 50% of the highest S/P ratio on day 28. Further results showed that IgM level decreased on day 35, and after second boosting, it reached a similar level as day 28 (Fig. 6).
Detection of IgA in RP-expressed BVDV E2-immunized pig. FMIA was performed in the presence of serum samples (1:100) of alphavirus RP-immunized animals and BVDV E2 antigen fragment (aa 1–183)-coupled microsphere beads. Results are reported as S/P ratios calculated from MFI.
Kinetics of BVDV E2-specific IgM in alphavirus RP-immunized pig. IgM antibody response was evaluated using FMIA in the presence of serum samples (1:100) of alphavirus RP-expressed BVDV E2 antigen-immunized animals and BVDV E2 (aa 1–183) protein-conjugated beads. Data represent S/P ratios.
BVDV E2-specific IgA, IgG, and IgM responses to alphavirus RP in oral fluid
IgA, IgG, and IgM responses to BVDV E2 (aa 1–183) over a time period were detected using multiplex FMIA in oral fluids and results are reported as S/P ratios (Fig. 7). BVDV E2-specific IgG was strongly reactive to E2 (aa 1–183) antigen compared with IgA and IgM. IgG was first detected 21 dpi, elevated after first boosting on day 21, slightly decreased on day 42, and then peaked at 49 dpi. IgM was first detected at 7 dpi and had decreased to near background on day 14. Again, IgM level was slightly increased on day 21 and observed to remained elevated at 28 dpi after first boosting on day 21 and then peaked on day 49 after second boosting on day 42. The data suggested that boosting enhanced antibody responses in oral fluids. IgM was first slightly detected on day 7 and then decreased to near background level.
Kinetics of BVDV E2 antibody isotypes (IgA, IgG, and IgM) in oral fluids. Reactivity of BVDV E2 (aa 1–183) antigen-coupled beads with oral fluids of alphavirus RP-expressed BVDV E2 antigen-immunized animals was tested in FMIA. Results are reported as S/P ratios.
Discussion
The production of antibodies after immunization is crucial to induce immunity and protection against viral infections. The current study investigated whether alphavirus RP-expressing BVDV 1b E2 glycoprotein antigen in pig can induce immune response specific to BVDV antigen. The goal of this study was to detect serum antibody responses against BVDV antigen following immunization with an alphavirus RP-expressed BVDV E2 antigen in pig. To evaluate BVDV E2-specific IgA, IgG, and IgM antibody responses after immunization with alphavirus RP-expressed antigen, a multiplex FMIA was performed using pig serum samples. A total of eight BVDV E2 antigen fragments were evaluated in this study. BVDV E2 (aa 1–183) showed the highest antibody response (Fig. 3), and E2 (aa 1–183) was considered the best target for the analysis of serum and oral fluid in this study. The data in the current studies are supported by previous findings on detection of IgG response against CSFV E2 (aa 1–181) antigen using FMIA (11). In addition, it has been reported that an immunogenic domain of the E2 antigen is located in the region, about 1–120 residues (21).
The detection of neutralizing antibodies against BVDV E2 ELISA-based test has recently been reported (2). However, limited research work has been reported on the evaluation of IgM antibody responses against BVDV E2 antigen. In this study, IgM antibody response was detected 7 days after immunization with alphavirus RP-expressed BVDV E2 antigen in pig (Fig. 6). The results showed that seroconversion of BVDV E2-specific IgM is 2 weeks faster than IgG. This conclusion is supported by our previous FMIA-based detection of CSFV E2-specific IgM antibody response on day 7 in pigs immunized with CSFV E2 alphavirus RP (11). In other studies, IgG and IgM antibody responses were detected using indirect ELISA test after immunization; IgM appeared 1 week faster than IgG (20). Based on ELISA results, FMIA-based detection of IgM might potentially be a rapid and useful tool for detection of antibody response in pig after immunization.
IgA, IgG, and IgM were detected in oral fluid samples collected from a group of pigs (pens) immunized with BVDV E2 antigen. It has been reported that the largest proportion of locally produced antibodies are secretory IgA by plasma cells in the salivary glands and duct-associated lymphoid tissue (3). BVDV E2-specific antibody isotype (IgA, IgG, and IgM) responses in serum and oral fluid samples are illustrated in Figures 4 –7. The kinetics of BVDV E2-specific antibody isotype in oral fluids and serum after immunization with alphavirus RPs have been contrasted in this study. Our results showed that IgM in oral fluid is very low; however, the detection of IgM in oral fluids of BVDV E2 alphavirus RP-immunized animal might be species specific (Fig. 7). In this study, the highest S/P ratio of IgG was detected in oral fluid, therefore suggesting that IgG in oral fluid in serum is transported through the oral mucosa into gingival crevices of the oral cavity (5). Interestingly, the results suggested that sequential immunization with BVDV E2 alphavirus RP and boosting with the same doses of RP have potentially enhanced antibody response in both serum and oral fluids. Previous reports have also demonstrated a high level of antibody response using viral vectors encoding FMDV capsid proteins and boosting with inactivated virus antigens (26).
In conclusion, detection of antibody responses against BVDV E2 antigens in pigs immunized with an alphavirus RP-expressed BVDV E2 antigen is a new development in vaccine study. The current multiplex FMIA-based detection of BVDV antigen-specific IgA, IgG, and IgM in swine immunized with alphavirus-expressing E2 is a simple, rapid, sensitive, and novel confirmatory diagnostic approach. Further research is needed to determine the level of neutralizing antibody in alphavirus RP-expressed BVDV E2-immunized pigs.
Footnotes
Acknowledgment
This work was supported by a USDA NIFA Award 2013-68004-20362.
Author Disclosure Statement
No competing financial interests exist.