
Abstract
Influenza virus infections remain a significant health burden worldwide, despite available vaccines. Factors that contribute to this include a lack of broad coverage by current vaccines and continual emergence of novel virus strains. Further complicating matters, when influenza viruses infect a host, severe infections can develop when bacterial pathogens invade. Secondary bacterial infections (SBIs) contribute to a significant proportion of influenza-related mortality, with Streptococcus pneumoniae, Staphylococcus aureus, Streptococcus pyogenes, and Haemophilus influenzae as major coinfecting pathogens. Vaccines against bacterial pathogens can reduce coinfection incidence and severity, but few vaccines are available and those that are, may have decreased efficacy in influenza virus-infected hosts. While some studies indicate a benefit of vaccine-induced immunity in providing protection against SBIs, a comprehensive understanding is lacking. In this review, we discuss the current knowledge of viral and bacterial vaccine availability, the generation of protective immunity from these vaccines, and the effectiveness in limiting influenza-associated bacterial infections.
Introduction
V
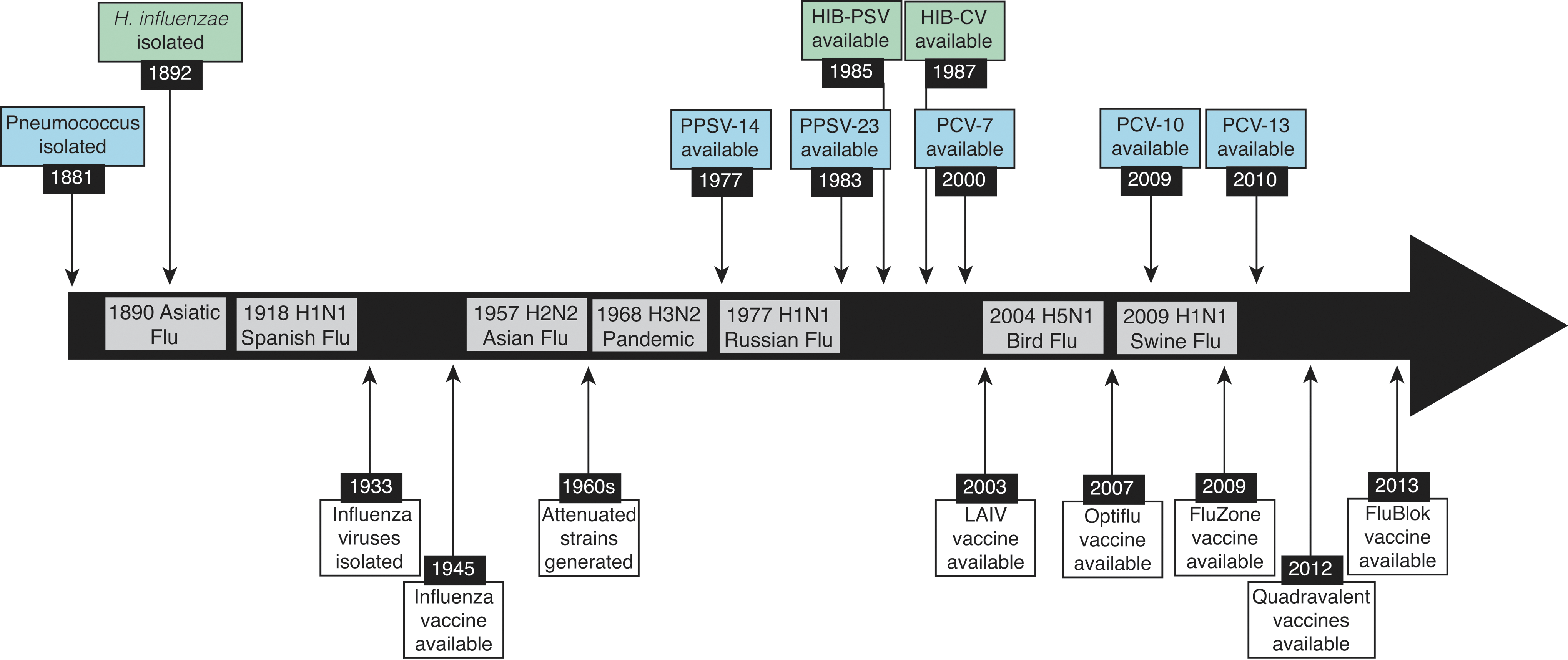
Timeline of available vaccines and influenza pandemics. Timeline of the major influenza virus pandemics from 1890 to 2017. Since the first isolation of an influenza virus in 1933, numerous vaccines have been developed and approved for use (white). Vaccines for pneumococcus (blue) and Hib (green), on the other hand, took nearly a century from the first isolation in 1881 and 1892, respectively.
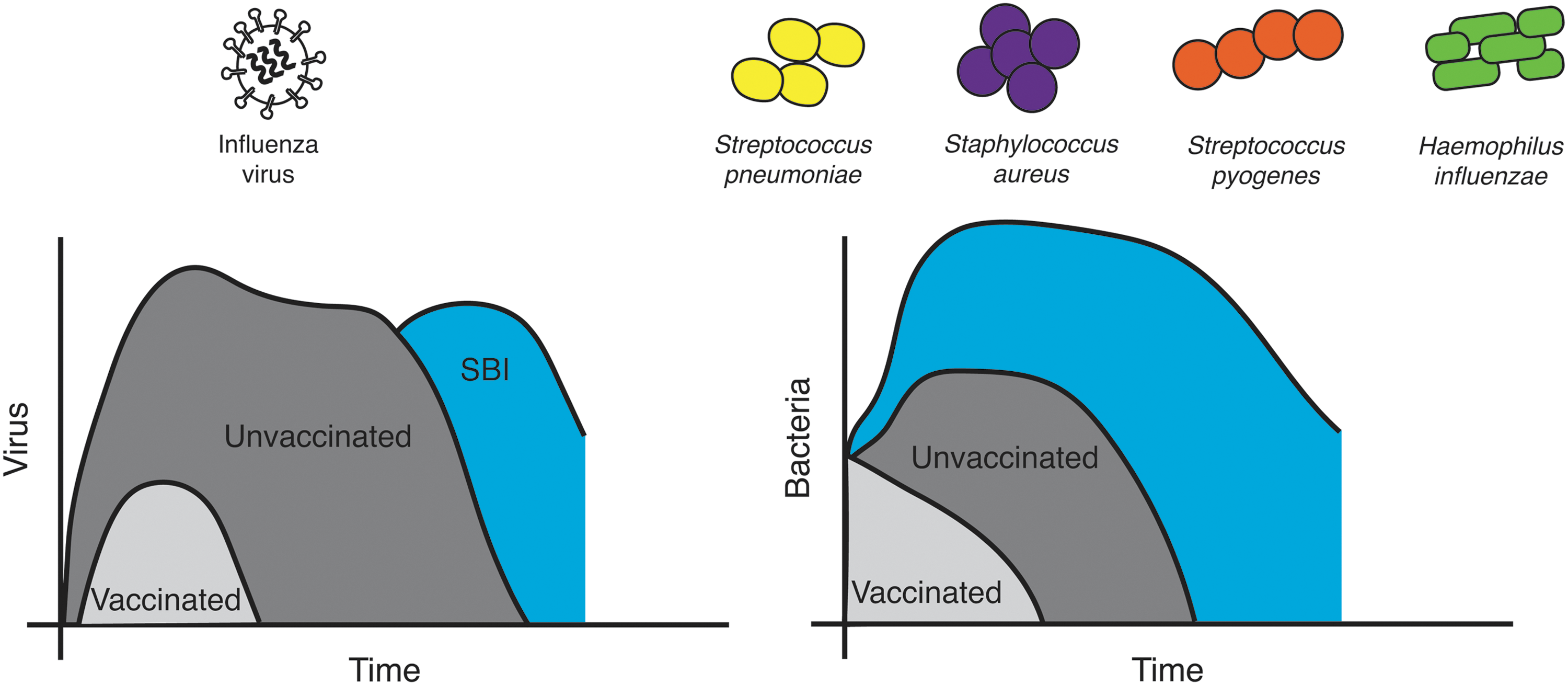
Time course of influenza-bacterial coinfections and major causative agents. Influenza virus infection in unvaccinated individuals (dark gray) results in rapid viral growth before the virus peaks and the infection resolves. Bacteria often invade late in the infection, which can result in a rebound of viral loads (blue). In vaccinated individuals (light gray), viral loads are reduced and the infection length is shortened considerably, leaving little opportunity for bacterial invasion. Similarly, primary bacterial infections with S. pneumoniae, S. aureus, or S. pyogenes in vaccinated individuals (light gray) result in more mild infection compared to those that are unvaccinated (dark gray). When these pathogens invade during influenza (blue), bacteria grow rapidly with little immune control.
Association between influenza virus and SBIs was first recorded by R.T.H. Laennec in his observations regarding excess mortality after an outbreak of la grippe in the 1700's (120). In current clinical settings, influenza-related SBIs can be difficult to diagnose because viruses other than influenza, including respiratory syncytial virus, parainfluenza virus, and adenoviruses are also known to initiate SBIs (62,94,95). Further complicating matters is the wide range of bacterial pathogens that are associated with community-acquired (34,94,95) and hospital-acquired (1,100) SBIs. The majority of those at risk for an SBI are the young (<1 year old) and the elderly (>65 years old) (202), and these populations notoriously have weak immune responses toward both infection and vaccination (138,155,189).
Despite the high mortality rates associated with SBIs, influenza-infected individuals may be more likely to recover without complication than succumb to an SBI (20), an observation that indicates the potential for identifying mechanisms that can overcome these deadly infections. Recent laboratory-based studies into the cause of SBIs have advanced our understanding of the viral, bacterial, and host contributions to these synergistic interactions [reviewed in (22,146, 154,201,204,215,224), among others]. Defining these host-pathogen interactions should give insight into how the host responds to an SBI. However, the extent to which vaccines and therapeutics can be used to reduce SBI severity remains under investigation, especially with respect to translating laboratory findings into the clinic. Recent work using laboratory-based models of SBI to evaluate vaccines that are either FDA approved or in development has helped identify the impact each has toward limiting these deadly diseases. Currently, the true impact of vaccines on the outcome of polymicrobial infections is difficult to evaluate, in part, because there is little systematic surveillance of bacterial coinfections during seasonal influenza (43,111). In addition, because the immune status of the human population varies tremendously and is difficult to model in the laboratory, well-designed and controlled clinical trials will be needed to determine the effects of vaccines on SBI severity. In this review, we review the use of vaccines to abrogate influenza-associated bacterial infections and discuss advances in this area, including both surprising findings and current gaps in our knowledge.
Vaccines That Target Influenza Viruses
The influenza virus is a likely target for circumventing a secondary bacterial invasion. There are a number of FDA-approved vaccines against influenza viruses that range from traditional inactivated influenza virus (IIV) preparations, live attenuated influenza viruses (LAIV), and recombinant proteins expressed by baculoviruses (77). Both IIV and LAIV vaccines are prepared by infecting either chicken eggs (71,114) or cells in culture (137,165) with master donor viruses, while recombinant protein vaccines are created without the use of influenza viruses (38,92). The main target of vaccine-induced immunity remains the viral hemagglutinin (HA) protein (256), and the hemagglutination inhibition titer is the gold standard for correlating vaccine-induced immunity with protection (15,17,81,152,176). In population studies, influenza vaccines often show variable effectiveness (180), with recent reports indicating ranges from 23% to 62% effectiveness against acute respiratory illness (28,65,66) depending, in part, on how well the vaccine matches the circulating strains (3,66,67). Continuous change in circulating influenza viruses creates a situation where vaccine formulations must be considered annually (3) and we are often unprepared to effectively mitigate the initial emergence of a pandemic strain (19,75). This gap in vaccine preparedness can subsequently affect the incidence of both primary influenza virus infections and SBIs (5,8,160,214,244), and lead to devastating consequences during both epidemic and pandemic influenza virus outbreaks.
Because the HA protein is constantly changing through antigenic drift (227), there have been efforts toward developing universal influenza vaccines that target conserved viral proteins (231). Three targets, including the ion channel matrix 2 extracellular (M2e) protein (47,172), the conserved stalk region of the HA protein (50,233,251), and the neuraminidase [NA; (36,107,109,131,210)], are of particular interest in these efforts. Each of these targets has the potential to induce broad immunity within a specific subtype and may even allow for heterosubtypic immunity (117,131, 136,206). To date, the only FDA-approved vaccine that induces immunity against a single influenza virus protein is the recombinant HA vaccine that is produced using baculovirus (38,92), and vaccines that target conserved epitopes may be able to build upon the success of this vaccine (55). One potential drawback to focusing on a single viral component for vaccination may be the absence of immunity against other viral proteins that may help limit SBIs, including the NA protein (88,147). In addition, because vaccine-induced immunity is not always sterilizing (134,257), even when the vaccine closely matches the circulating strain, the effect that a mild or moderate infection can have on the host must be considered. Indeed, vaccines that target conserved influenza virus proteins often generate antibodies that limit virus spread, rather than prevent infection (2,50,58,127, 152,245). This can yield a mild infection that may leave the vaccinated host susceptible to an SBI, further justifying studies that determine the impact of influenza vaccines on SBI severity, both in the laboratory and the clinic (14,30,126, 156,178,243).
Effect of Influenza Vaccines on SBIs
To date, only a few clinical studies have reported on the influence of vaccine-induced immunity against influenza virus on subsequent SBI severity. One of these studies used military recruits, a group known to be susceptible to S. pyogenes infections (126). This study showed that vaccination against influenza viruses limited illness associated with S. pyogenes in this population, which was a promising finding in the area of vaccine-induced prevention of SBI severity. As further evidence of influenza virus vaccine-induced prevention of SBIs in humans, reduced incidence of otitis media was observed in pediatric populations during clinical trials that preceded approval of the LAIV vaccine (14). Together, these studies encourage future meta-analyses that may help determine the impact of influenza vaccines on SBIs. However, due to the complex nature of SBIs, direct evaluation of the benefit that influenza virus vaccines has on SBIs in humans remains limited to defined patient populations where conditions can be controlled. Expanding these studies into larger populations is difficult due to an inability to control prior immunity against influenza viruses and bacterial species, including immunity induced by vaccination (156).
Experiments that evaluate vaccine-induced immunity in the context of SBIs can be difficult to control clinically, but key concepts related to vaccine-induced protection can be modeled in the laboratory. To date, the majority of vaccine studies are designed to evaluate protection against the pathogen the host was vaccinated against, and there have only been a limited number of studies evaluating influenza virus vaccines in the context of an SBI. One such study used IIV preparations to determine whether immunity against the HA protein or the NA protein was more effective in limiting SBI severity (88). Results from this study showed that immunity against either protein could limit the severity of secondary infections when S. pneumoniae was used as the secondary invader. Because the NA expressed by an influenza virus is known to increase both bacterial attachment (147) and SBI severity in animal models (186,187), and NA is one of the potential vaccine targets mentioned above, these findings justify continued efforts to evaluate the contribution of anti-NA immunity toward SBI severity.
In separate studies in mice, IIV and LAIV vaccines were evaluated for their ability to limit secondary S. pyogenes infection (30,177). In these studies, vaccine-induced immunity against influenza virus improved survival after an SBI, but did not always yield complete protection. Interestingly, these studies show that S. pyogenes can establish a lung infection in mice that were challenged with influenza viruses that matched the vaccine strain (30). This is surprising because the vaccinated mice in these studies did not show signs of infection after virus challenge, observed using weight loss, yet bacterial persistence and death following inoculation with a sublethal dose of S. pyogenes were observed in some mice (30). Because influenza vaccines are not always able to completely protect against an SBI, the immune mechanisms associated with severe SBIs in the vaccinated host could be further elucidated as a step toward translating these findings into clinical practice. Similarly, future vaccine studies could focus on defining the impact of supplementing antiviral and antibiotic options in situations when vaccine and challenge strains do not match.
Despite the limitations described above, it is clear that vaccinating against influenza viruses limits SBI severity in the majority of vaccinated animals (30,88,157). However, the LAIV vaccine has an important caveat in laboratory studies, which shows that bacterial colonization and replication may be enhanced in the nasopharynx and middle ear after LAIV delivery (158,159). Changes in commensal microbiota within the nasal passage after LAIV administration (241) may contribute to this increased colonization and replication. This observation brings into question the value of evaluating the timing of anti-influenza and antibacterial vaccines in an effort to limit colonization, particularly in pediatric populations. Indeed, carriage of H. influenzae, but not pneumococcus, may be moderately increased in children within 1 week after LAIV administration (243). Similar to the animal studies, increased pneumococcal density in children was observed, but not until 28 days after vaccination (243). Thus, the impact of influenza vaccines on subsequent bacterial infections should be considered both in the days after vaccination and at the peak of influenza season.
Vaccines That Target Bacterial Pathogens
While influenza viruses initiate SBIs, bacteria often dictate illness severity (68,132,254). As such, the role of vaccines against bacterial pathogens in limiting coinfections must be discussed. Clinical (211) and laboratory (112,153) studies demonstrate bacterial vaccine effectiveness against SBIs, and use of bacterial vaccines in this capacity dates back to the 1918 pandemic (32,59,247). In 1918, vaccines were often crude preparations of bacterial membranes that were delivered without the benefit of strict clinical trial criteria, yet protective efficacy was conferred in some (32,59), but not all (26,125,143) of these clinical experiments. Similar to influenza vaccines, the efficacy of bacterial vaccines is limited to strains included in the vaccine, which can be important when the circulating serotype is not included in the vaccine (80). Further complicating matters, studies designed to evaluate bacterial vaccines in SBIs range in their reported benefit from complete protection against an SBI (112) to no protection observed (153,157). This efficacy range shows a benefit to vaccinating against bacteria even when influenza-induced illness is observed prior to bacterial inoculation. Given that few vaccines are available for coinfecting bacteria and identifying universal epitopes that can induce protective immunity within a bacterial species is difficult, the extent that antibacterial vaccines are efficacious for SBIs is not fully appreciated. These issues are discussed in more detail below.
The primary bacterial species associated with severe SBI includes S. pneumoniae, H. influenzae, S. aureus, and S. pyogenes (Fig. 2). Vaccines are available for S. pneumoniae and H. influenzae, but not for S. aureus or S. pyogenes (Fig. 1). Vaccines against S. pneumoniae, prepared using purified polysaccharide, have been available for decades as 23-valent preparations that are recommended for adults (221). Pneumococcal conjugate vaccines have been approved for use by the FDA since 2000, initially as 7-valent preparations and more recently in a 13-valent variety (190). Together, vaccines that protect against S. pneumoniae are regularly given to adults over 60 years of age and are included in the standard vaccination schedule for children (194). Despite these vaccination efforts, pneumococcus remains the most common bacterial pathogen associated with complications after influenza virus infection (94,95,173). This is likely due to the lack of comprehensive coverage of pneumococcal serotypes within a single vaccine preparation, which can become a bigger problem when nonvaccine serotypes circulate among vaccinated populations (4,191).
Vaccines that target conserved epitopes that are expressed by multiple S. pneumoniae isolates may help improve the breadth of protection, even if the target is not necessarily universal. In an effort to increase the breadth of serotypes affected by vaccination against S. pneumoniae, the PspA protein has been studied as a vaccine candidate since 1994 (149). A recent screen of 52 different pneumococcal proteins identified another seven potential targets (PhtB, PhtD, PhtE, PrtA, NanA, PavB, and Eng) with protective capabilities (7). However, to date, there have been no published studies reporting on the contribution of vaccine-induced immunity against any of these proteins in preventing severe SBIs. When considering conserved epitopes like PspA, it is important to determine whether antibodies would overcome influenza virus-induced defects in host immunity at the time an SBI is initiated (154). Vaccines that target these conserved epitopes would be of value, but would require additional testing in animals to validate their contributions toward protection against severe SBIs.
In contrast to S. pneumoniae, vaccines against H. influenzae b (Hib), which were introduced initially as polysaccharide vaccines in 1985 (185) and then as a conjugate vaccine for use in children (128) (Fig. 1), have largely reduced Hib incidence globally (16,64,209). The fact that there are fewer Hib serotypes compared to the >90 serotypes of S. pneumoniae (190), may have helped to improve Hib vaccine effectiveness. Overall, bacterial vaccines have demonstrated a benefit in preventing primary infection with these pathogens, which in turn helps reduce the incidence and severity of SBIs (Fig. 2).
Although vaccines against S. pneumoniae and Hib are available, vaccines against S. aureus remain in development (161). S. aureus is a particular concern as it was a frequent cause of SBIs during both the 1957 and 1968 influenza pandemics (148), and it now accounts for a large number of influenza-associated childhood fatalities (262). In fact, the emergence of methicillin-resistant S. aureus (MRSA) (99,182,184), which can cause a severe and deadly necrotizing pneumonia (49,198), has contributed to this increased severity of SBIs. Recurring infections complicate therapies designed to combat S. aureus infection, and vaccinating against S. aureus antigens has had limited success (23,46,113,162). Advancing vaccine efforts against S. aureus could consider identifying and developing vaccines that induce broad immunity against conserved epitopes, and possibly using antigens from other organisms with similar epitopes to provide cross-protective immunity (260). Until a preventative measure is available, antibiotics remain the primary therapeutic option. However, these drugs often have reduced efficacy once an invasive SBI has established (103,104,222).
Similar to S. aureus, vaccines against S. pyogenes are not available clinically (234). S. pyogenes, which is also known as group A streptococcus, is a bacterial species that is associated with asymptomatic carriage of the skin and respiratory tract (240), which can cause both noninvasive and invasive diseases. Noninvasive diseases caused by S. pyogenes include pharyngitis (140), scarlet fever (140), pyoderma (40), and cellulitis (122), while invasive diseases caused by these bacteria include pneumonia (169), erysipelas (27), streptococcal toxic shock (124), and necrotizing fasciitis (35,39,261). Even after S. pyogenes is cleared, host responses against the bacteria can cause acute rheumatic fever (248) and poststreptococcal glomerulonephritis (25). It is estimated that lower respiratory tract complications are associated with 11–12% of invasive S. pyogenes infections (179) and there is a 38% case fatality rate given this diagnosis (169). While antibiotic resistance has been observed, in particular against macrolide antibiotics (76), S. pyogenes remains susceptible to penicillin (203). However, as with other SBI cases, these antibiotics are often unable to clear SBIs in hospitalized patients (103,104,222). Without clinically available vaccines against MRSA and S. pyogenes, evaluating the impact of these vaccines on SBI severity is limited to the laboratory (112).
Vaccinating Against Bacteria to Reduce SBI Severity
The effect that antibacterial vaccines have on reducing the severity of an influenza-associated SBI has been assessed for its impact on survival in animal SBI models using either S. pneumoniae or S. pyogenes as the secondary pathogen (112,153). Both the pneumococcal conjugate vaccine, Prevnar (153), and a recombinant M protein vaccine that targets S. pyogenes have been evaluated (112). In the case of Prevnar, the vaccine was <40% protective against a secondary infection, while the vaccine targeting S. pyogenes showed complete protection against SBI. Both of these studies relied on challenge with bacterial isolates that matched those included in the vaccine, which partially limits the translation of this knowledge into clinical settings where bacterial species cannot be controlled. The observed survival in these SBI models is impressive considering that influenza-induced illness was observed before inoculation with bacteria (112). This observation implies that the known virus-induced deficits in host immunity that predispose to severe SBIs (154) can be at least partially overcome in the presence of vaccine-induced antibacterial immunity. In the case of S. pyogenes, antibodies against the M protein could induce antibody-dependent cellular phagocytosis (ADCP) by macrophages, which was observed in vitro using sera from vaccinated mice (112). This potential contribution of vaccine-induced antibodies toward clearance of a secondary bacterial invader may help limit SBI severity, but further work in this area is needed to define relevant host mechanisms of protection.
Vaccine-Induced Immune Responses Affecting SBI Severity
When considering the effect of vaccine-induced immunity on SBI severity, one must understand how vaccination influences immune responses in the context of both the primary pathogen and the secondary pathogen of interest. However, to date, the majority of vaccine studies focus solely on vaccine-induced responses against the pathogen the vaccine was designed to protect against (156). Previous work with influenza vaccines in SBI models indicate that a severe SBI can develop even when the challenge strain matches the vaccine strain (30,177). Host factors that can influence SBI severity include changes in the microbiota, innate immune responses, and adaptive immune response. Recent work in the microbiome field shows that the unique group of microbes that comprise the microbiome changes during infection (29,89,195,232) and may influence SBI disease severity (192). Similarly, host responses from vaccination have the ability to directly alter the resident microbiota (175,205,241). While the role of the respiratory and gut microbiome in SBIs is of significant interest (48,142), we will focus the remainder of our discussion on contributions from innate and adaptive immunity.
Innate immune responses against influenza viruses can directly influence severity of influenza-associated SBIs. Numerous innate immune responses have been shown to play a role in SBI acquisition, viral and bacterial growth, and severity. Many of these are likely time dependent and may be correlated. Bacterial invasion and initial growth kinetics have been shown to be a consequence of viral-mediated alveolar macrophage (AM) depletion (74,223,225), while the subsequent neutrophil response is dysfunctional and these cells are unable to limit the later stages of bacterial growth (208,236). An impaired NK cell response, which may be upstream of the effects from AMs and neutrophils, has also been identified to have a role in the defective postinfluenza antibacterial defense (220). In addition, differences in the expression of cytokines, such as type I interferons (IFNs), can alter the function of these cells and enhance SBI severity (130,171,212,213). The contribution of innate immune responses is important to consider because they can be induced even when the infecting virus matches the vaccine strain, the vaccine-induced immunity is sterilizing, and the circulating virus has not mutated sufficiently to escape vaccine-induced immunity. Indeed, a live attenuated Bordetella pertussis vaccine demonstrated protection against lethal influenza virus challenges by controlling cytokine responses and reducing virus-mediated inflammation (129).
In addition to innate immunity, the dynamic interactions of an invading pathogen and the host adaptive immune response, including cell-mediated immunity (i.e., T cells and B cells) and antibodies (i.e., IgG, IgM, and IgA), have also been studied in the context of SBIs. In fact, recent work in mice shows that virus-specific antibody responses, B cell and T cell populations, and germinal centers are reduced during influenza-pneumococcal coinfection (18,255,258). Although animal models of influenza-bacterial coinfection are lethal, these immune alterations may indicate a reduction in the quality and quantity of protective immunity following coinfection in nonlethal situations. Treatment with influenza virus-specific immune serum can partially restore the response, improving survival (258), which may indicate a protective role for anti-influenza virus antibodies. Interestingly, when pneumococcal infection precedes influenza virus infection, the B cell and CD4+ T follicular helper cell responses are enhanced (255). However, the robust response is short lived with waning antibody neutralization within 30 days of infection (255). Further complicating matters, these innate and adaptive immune responses differ in unvaccinated and vaccinated individuals, which can influence the effectiveness of vaccines in SBIs.
As mentioned previously, laboratory and clinical studies have often focused on developing vaccines against single pathogens and evaluating each using challenge studies that incorporate the pathogen of interest. However, there are often off-target effects of vaccines that are underappreciated due to a lack of investigation within polymicrobial infection models (156). Because the majority of our knowledge of vaccine:challenge studies relies on outcomes using the single-pathogen approach, these responses can be considered when selecting the characteristics of vaccines that would most likely influence the severity of an SBI. This relies largely on our collective knowledge of humoral and cellular correlates of vaccine-induced protective immunity, and whether those aspects of the adaptive immune response may be specifically depleted or made ineffective during an SBI. Because postinfection innate immunity in vaccinated individuals and changes in the microbiome can be difficult to consider based on the available information, we will focus our discussion on known mechanisms of adaptive immunity, which may alter SBI severity, in particular those mediated by vaccine-induced antibodies.
Host Responses to Influenza Virus Vaccines
One main goal of vaccination is to generate neutralizing antibodies that prevent infection and, ideally, induce long-term sterilizing immunity (257). Achieving this level of protection is also useful for limiting, but not necessarily preventing, SBIs (30,177). Vaccines that stimulate neutralizing antibodies, including those against influenza viruses (249), are often most effective when these antibody titers achieve high levels. However, efficacy is often lost as antibody levels wane over time (207). In some instances, vaccine-induced immunity can wane within a single influenza season, which can affect vaccine effectiveness against viruses that circulate later in the season (63). In addition, both historical (82 –84) and recent evaluation of vaccine effectiveness have resulted in the antigenic distance hypothesis (226), which predicts the current season's vaccine effectiveness based on negative interference from the previous season's vaccine. This hypothesis continues to be tested to determine how repeated vaccination alters susceptibility to influenza virus infection, with some evidence that repeated annual influenza vaccination reduces vaccine effectiveness (12,217). This may be due to mutations of the influenza virus HA that arise while growing the viruses in cell culture (219) or in eggs (218), which are both used during vaccine production (71,137). Thus, challenges remain with respect to influenza vaccines and host responses to these vaccines that must be considered in the effort to improve effectiveness, especially as it relates to the impact of vaccination on SBIs.
From the host response perspective, antibodies may have the ability to interact with Fc receptors in addition to neutralizing virus. This can assist in virus clearance when sterilizing immunity is unavailable (45,50,86,87,168). Similarly, antibodies targeting conserved epitopes can rely on Fc receptor interactions to enhance clearance from an infected host (45,50,51,58,127,245). Thus, it appears that a balance of neutralizing and nonneutralizing antibodies encompasses an optimal response against influenza virus infection. At this time, the majority of immune assays focus on the neutralizing characteristics of the antibody, as defined by their interaction with an antigen through the Fab region (183,197,249), and only more recently has there been strong consideration for the contribution of the Fc region of the antibodies toward virus clearance (21,45,72,73,96,97,166,167). This change in focus to evaluate the whole antibody toward pathogen clearance has impacted our knowledge of the contribution of conserved targets toward clearing pathogens from an infected host (45,50,51,58,79,127,168,239,245,246), which we will discuss below.
The influenza vaccine field has focused recently on the development of universal vaccines that target more conserved epitopes of the influenza virus, including the M2 ion channel (47), the HA stalk region (110), and the NA protein (131). One issue facing these vaccines is the reduced ability of vaccine-induced antibodies to directly prevent infection in the conventional way that we envision neutralizing immunity. For example, because the M2 ion channel is expressed in higher abundance on infected host cells rather than the virion itself (123,263), the infected cell becomes the target of vaccine-induced immunity (58). Thus, vaccine efficacy relies on establishing at least a mild infection for vaccine-induced antibodies to eliminate the pathogen. As discussed above, this initial infection may have profound effects on the microbiota, innate immunity, adaptive immunity, and SBI severity.
Similarly, antibodies against the HA stalk region do not interfere with HA binding to sialic acid residues through the receptor-binding region of the HA (37,57). However, in contrast to antibodies against M2, antistalk antibodies appear to directly enhance ADCP (168) when viruses concomitantly bind host cells through sialic acid residues, while linking stalk-specific antibodies to the cellular Fc receptor (50,239). Host responses that develop during this interaction, including cytokine production by macrophages, have yet to be defined and may also affect severity of an SBI. Furthermore, the intracellular trafficking of these internalized viruses brings into question whether vaccine-induced antibodies enhance virus clearance through ADCP or if this interaction may contribute to the antibody-dependent enhancement of infection responses that have sometimes been observed in humans (237,238) and large animal influenza vaccine studies (69,70,105,196,229). There are clearly benefits of targeting conserved epitopes and developing vaccines that are broadly reactive against influenza viruses. However, additional research into the cellular and molecular immune responses that develop when these antibodies are actively clearing the virus from an infected host is needed.
Finally, antibodies against the NA have been of interest for decades (54,98,107,108,170,210). Their role in protection has been assigned to limiting virus spread rather than directly neutralizing virus particles (108,109). The NA is an intriguing target when discussing SBIs as it has an enzymatic activity known to enhance SBI severity (147). In addition, because the NA expressed by influenza A viruses with pandemic potential (e.g., H5 N1) can match the NA expressed by seasonal influenza viruses (e.g., H1 N1), NA immunity may provide some protection from these pandemic viruses (131,139,206). This potential impact of vaccine-induced anti-NA immunity on a future pandemic has led to a refinement of the NA inhibition assay with an emphasis on using this assay to define anti-NA antibodies as correlates of vaccine-induced protection (56,152,163). Furthermore, a contribution of anti-NA immunity toward limiting SBI severity has been demonstrated in a murine superinfection model (88). At this point, it is unknown whether anti-NA antibodies limit SBI severity by inhibiting NA activity itself or through another host immune mechanism, but the potential for this vaccine target to limit secondary infections has been demonstrated experimentally.
Host Responses to Bacterial Vaccines
In contrast to the more recent appreciation for Fc receptors toward influenza virus clearance, ADCP as a mechanism of pathogen clearance has been appreciated for decades when discussing the effectiveness of vaccines against bacteria (24,44,61,116,193). Specifically, antibodies induced in response to S. pneumoniae are often tested in opsonophagocytosis assays (24), and vaccines in development against S. pyogenes were designed to target opsonic epitopes of the M protein (41,42,85). Thus, it is not surprising that antibodies induced against S. pyogenes are able to increase bacterial uptake by macrophages, in a manner that may help reduce SBI severity (112). However, influenza virus infections have profound effects on the macrophage population, including defects in cellular function that contribute to SBI severity (141,154,235). Thus, it seems plausible that vaccinating to induce Fc receptor-interacting antibodies that also recognize bacteria through their Fab region may help overcome virus-induced macrophage defects. Future evaluation of vaccine-induced antibodies should include efforts that determine ways to optimize Fc receptor-mediated clearance of bacteria. This would determine whether mechanisms that are known to contribute to immune-mediated protection against primary bacterial infection can be effective during an SBI or if other immune mechanisms of protection should be considered.
Alternative Interventions
In addition to vaccine-induced antibodies, the contribution of antiviral and antibiotic therapies toward limiting the severity of an SBI must be considered. From the antiviral side, neuraminidase inhibitors are effective against influenza A and B viruses (151) and can reduce the duration and severity of SBIs in animal models (144,145,222). On the other hand, antibiotics can differ in their efficacy in SBIs (103,104), a finding that is appreciated clinically (60) and experimentally with models showing that bacteriostatic antibiotics are more effective against SBIs than bactericidal antibiotics (103). Interestingly, computational models indicate a strong benefit if combination therapy were employed (222), but this scenario has not been tested in animal models or in the clinic. When considering expanding our antimicrobial repertoire, it is also important to understand how these drugs affect the microbiome, which is known to be affected by some antimicrobials (101). In addition, it is imperative that new therapies be evaluated using models of SBIs to ensure their use will be beneficial when vaccines are unavailable (e.g., during a rapidly developing pandemic) and to guide the appropriate selection and timing of therapeutic treatment.
Another potential approach that has been increasing in popularity and may represent a first-line treatment in the face of a pandemic, is immunotherapy using antibodies (52,91, 216,230). This is interesting because the use of passive antibody therapies revisits classical approaches with convalescent sera that were used to limit SBI severity during the 1918 pandemic (135). Today, therapeutic antibodies can be monoclonal (118,188,250,252) or polyclonal (52,106,200) in nature and can be used to target either the virus or the bacteria. As described above, optimal protection against influenza virus infection seems to benefit from a balance of neutralizing and nonneutralizing antibodies (6,121,239), a finding that hints that polyclonal antibody preparations may provide a more balanced approach toward therapeutic treatment. These polyclonal antibodies may be created using cocktails of monoclonal antibodies with demonstrated efficacy against their individual targets (78,90,216,228,230). Interestingly, off-target effects for these products are possible, including a global anti-inflammatory response that has been observed in studies testing human intravenous immunoglobulin (9,10,174). Studies on therapeutic antibody preparations would benefit from validation in SBI models, as both direct and indirect effects on SBI severity could be appreciated before use in the clinic.
Concluding Remarks
Several studies indicate a benefit of viral and bacterial vaccination on the potential deadly outcomes from SBIs, despite the fact that most of these vaccines were not preevaluated in SBI models. In particular, individual studies reporting decreases in otitis media and S. pyogenes infections in humans that were vaccinated against influenza (14,126) provide evidence that these vaccines can limit SBI severity. To date, how SBI severity changes in individuals vaccinated against bacterial pathogens, but not against influenza viruses is unknown. Laboratory models evaluating bacterial vaccines in the context of SBIs have shown mixed results (112,153). Thus, additional testing of these findings in the laboratory and clinic is warranted. Furthermore, as the number of bacterial vaccines increases and as more conserved targets for influenza virus vaccines are identified, there is an opportunity to determine optimal host responses that can be targeted to reduce SBI severity after vaccination.
From a clinical standpoint, work with both influenza and bacterial vaccines suggests that SBIs can be mitigated through robust vaccination. Because of this benefit, annual influenza vaccine efforts could be complemented with annual bacterial vaccines for maximum efficacy. To our knowledge, studies that evaluate SBIs in animals that have been vaccinated against both the challenge virus and the challenge bacteria have not been reported, which may be due to technical difficulties in modeling this clinically relevant scenario in the laboratory. Examining the potential of complementary vaccination may be best suited for computational investigation, which is more amenable to assessing complex phenomena and clinical situations. Currently, bacterial vaccines are indicated for the young and the elderly (115,190), with minimal use in healthy adults. If these vaccines were tested for their simultaneous effects on SBI severity, rapidly characterizing their impact could help preparedness for future epidemics and, importantly, pandemics where viral vaccines may not be available. Gaps in vaccine coverage for S. pneumoniae isolates and an absence of vaccines against S. aureus and S. pyogenes leave us with incomplete bacterial vaccine coverage. Future exploration in antivirals, antibiotics, and immunotherapies must continue to consider how vaccine-induced immunity affects host responses that contribute to severe SBIs as part of preclinical validation studies.
Footnotes
Acknowledgments
The authors received support from NIH Grants 1 R56 AI125324 01 (A.M.S.), 5 R44 AI117976 02 (VCH), and 5 P20 GM103443 16 (V.C.H.).
Author Disclosure Statement
No competing financial interests exist.