
Abstract
Human parvovirus B19 (B19), a human pathogen of the erythroparvovirus genus, is responsible for a variety of diseases. B19 cause less symptoms in healthy individuals, also cause acute and chronic anemia in immunodeficiency patients. Transient aplastic crisis and pure red cell aplasia are two kinds of anemic hemogram, respectively, in acute and chronic B19 infection phase, especially occurring in patients with a shortened red cell survival or with immunodeficiency. In addition, B19-infected pregnant women may cause hydrops fetalis or fetal loss. B19 possesses high affinity to bone marrow and fetal liver due to its extremely restricted cytotoxicity to erythroid progenitor cells (EPCs) mediated by viral proteins. The nonstructural protein NS1 is considered to be the major pathogenic factor, which has been shown to inhibit the differentiation and maturation of EPCs through inducing viral DNA damage responses and cell cycle arrest. The time phase property of NS1 activity during DNA replication and conformity to transient change of hemogram are suggestive of its role in regulating differentiation of hematopoietic cells, which is not completely understood. In this review, we summarized the bridge between B19 NS1 and Notch signaling pathway or transcriptional factors GATA, which play an important role in erythroid cell proliferation and differentiation, to provide a new insight of the potential mechanism of B19-induced differential inhibition of EPCs.
Background
Human parvovirus B19
B19 infection could cause erythema infectiosum or transient aplastic crisis (TAC) in acute infectious period, also the persistent existence of B19 in bone marrow lead to pure red cell aplasia (PRCA) in an immunocompromised host (41). Life would be threatened in patients with shortened red cell survival due to their lack of timely supplements in B19-induced severe anemia (65). Furthermore, pregnant women with B19 infection have higher risk of miscarriage or fetal complications (15). The rate of vertical transmission during maternal parvovirus B19 infection is estimated at 33%, with fetal complications occurring in 3% of infected women (84). B19 poses a potential hazard to the fetus as crossing the placental barrier and infecting erythroid progenitor cells (EPCs) in bone marrow and fetal liver to block fetal erythropoiesis causing profound anemia, fetal hydrops, and/or fetal death (4). Once the fetus/newborns develop hydrops, treatments like intrauterine red blood cell transfusion or intravenous immunoglobin and digitalis may fail to rescue, with a survival rate of only 60–70% overall (42,49).
B19 is a potent inhibitor of erythropoiesis, due to its highly restricted cytotoxicity to EPCs at the burst-forming unit and colony-forming unit erythroid stages (13,96). Severe anemia could appear in B19-infected patients with immunodeficiency or with shortened red cells and in fetus/newborns born to B19-infected women. B19 is an important antigen for Eugenics (4). However, pathogenesis of B19 is not completely clear on account of its difficulties in vitro culture, effective vaccines, or antivirus drugs of B19 being of urgent shortage.
Pathogenesis of Anemia in B19 Infection
Even though B19 infection is self-limited in healthy individuals, it is often associated with TAC in children with sickle cell disease (16). In patients with a short life span of red blood cells, such as sickle cell anemia, hereditary spherocytosis, thalassaemia, or chronic hemolytic disease, severe anemia exacerbated by B19 infection could be fatal due to the acute hemolysis and temporary arrest of erythropoiesis occurring on the basis of chronic hemolytic anemia and increased destruction of red blood cells if transfusions are not available or not administered urgently.
Even more seriously, chronic B19 infection could induce PRCA, which is exhibited in immunocompromised patients or transplant recipients on account of impaired ability of viral elimination (53). Also, PRCA in the intrauterine B19-infected fetus may show ultrasonographic signs of general edema, such as subcutaneous edema, pleural effusion, pericardial effusion, ascites, and placental edema. The main mechanism responsible for the nonimmune hydrops fetalis is probably cardiogenic heart failure secondary to severe anemia (58).
The B19 virus can be transmitted mainly by respiratory tracts, also through blood or pooled-blood products, from a pregnant mother to her fetus, and possibly even from tattooing (70). After overcoming the barrier of airway epithelium, vascular, or placenta, B19 arrives into the human body and initially interacts with P antigen (globoside), the primary low-affinity attachment sugar molecule abundantly expressed on the surface of EPCs, acting as an essential cellular receptor for B19 (5). However, P antigen is necessary for binding but not sufficient for parvovirus B19 entry into cells (6,95).
Unlike mature human red blood cells, primary human EPCs, the major target cells of B19, not only express high levels of P antigen, but specifically express α5β1 integrins, which permit β1 integrin-mediated entry of parvovirus B19 (94). Moreover, Ku80 was identified in vitro nonerythroid cells to function as a coreceptor for B19 infection together with P antigen and α5β1 integrins, which takes part in B19 binding and subsequent entry. Although originally known as a nuclear protein, Ku80 was found to have a high expression on the surface of EPCs expressing glycophorin A as well as on the surface of immune cells such as CD20+, CD3+, or CD14+ cells in bone marrow, which may explain the pathologic immunity in autoimmune diseases related to B19 infection (60).
Thus, B19 virion accomplishes its internalization inside EPCs through interaction with P antigen in the aid of α5β1 integrins and Ku80, and initiates its replication after entering the nucleus. Differential inhibition processes and apoptotic signals activated by massive replication of B19 eventually lead to cytolysis of EPCs and release of virions into the blood, which is consistent with the transient high-titer viremia in the acute phase. Destruction of a large amount of EPCs significantly influences the erythropoiesis and life span of red blood cells, which brings about acute hemolysis (Fig. 1).
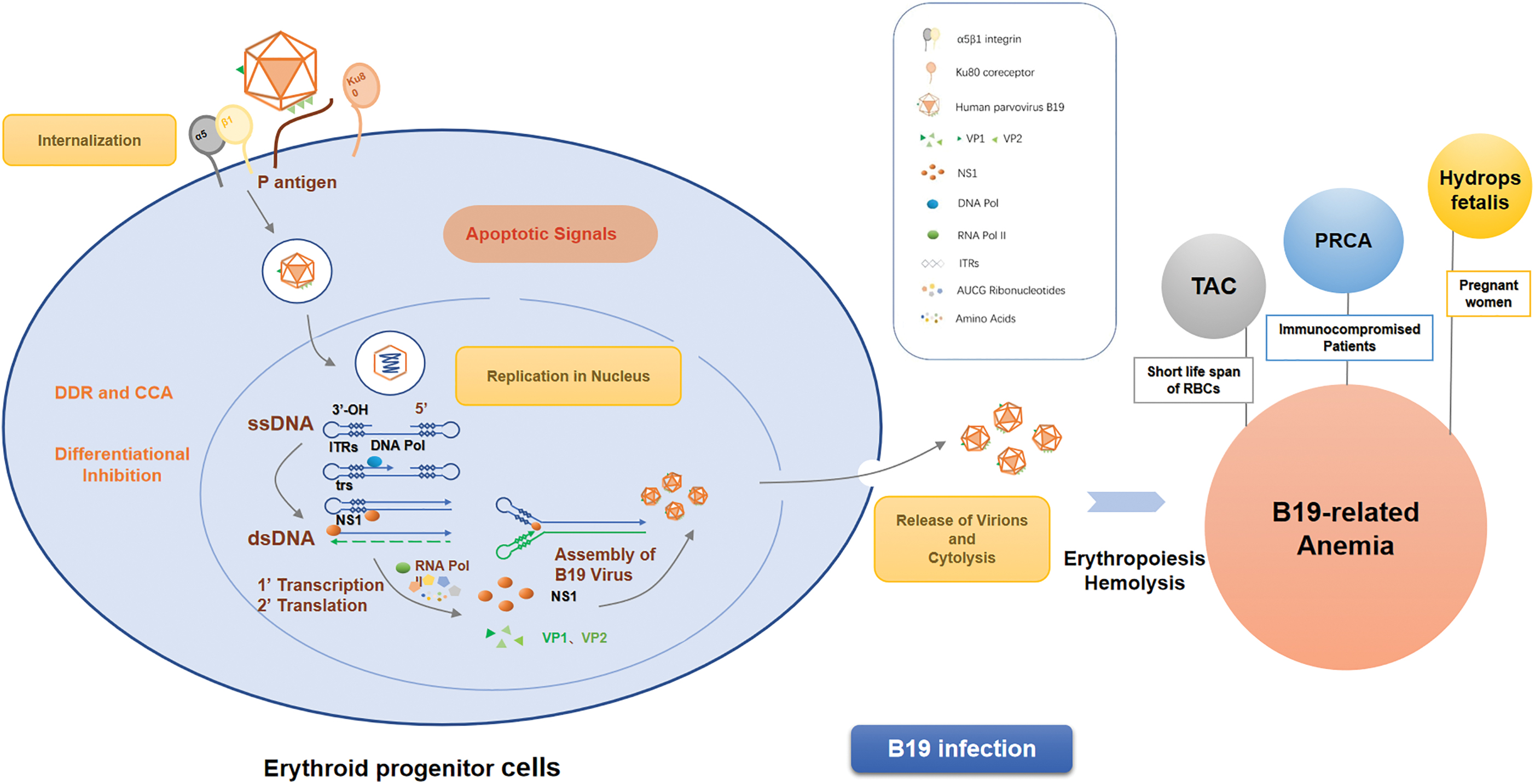
Mechanism of B19 causing erythropoiesis and hemolysis. B19 entry causes erythropoiesis by binding α5β1 integrins and coaction with P antigen. Replication of B19 leads to cytolysis of EPCs and influences the life span of erythropoietic cells, which brings about acute hemolysis. B19, human parvovirus B19; EPCs, erythroid progenitor cells. Color images are available online.
Nonstructural Protein NS1 Is the Major Pathogenic Factor in B19 Infection
B19 structure
The B19 virion has a linear ssDNA genome of 5– 6 kb, and a nonenveloped, icosahedral protein shell of ∼280 Å in diameter, known to be the smallest DNA virus so far (33,98). The 5,596 nucleotide-long genome is made up of an internal coding sequence flanked on both sides by identical inverted terminal repeats (ITRs) (70). These palindromes can acquire a hairpin configuration and serve as primers for complementary strand synthesis, while the central region genome encodes the five kinds of proteins of B19 virion (68). The two structural proteins, VP1 and VP2, account for 4% and 96% capsid proteins to form the icosahedral protein shell, respectively (28). The region of them shows a greater sequence variation in contrast to the large nonstructural protein (NS1) region that is highly conserved, which implies their function in host antivirus responses (68). Expressions of the other two small nonstructural proteins of 11 and 7.5 kDa were also documented (46,83), the former one was suggested as a potent inducer of apoptosis through enhancing viral DNA replication and virion release (8,20), whereas the function of 7.5 kDa protein is inconclusive.
Roles of nonstructural protein 1 in B19 infection
The large nonstructural protein NS1, located predominantly in the nucleus, is the major pathogenic factor in B19 infection (66). NS1 gene (616–2,631 bp) encodes its protein of 672 amino acids with a molecular mass of ∼78 kDa, which is of critical importance in both early virus DNA replication and transcription in B19-infected human EPCs (70). NS1 has a N-terminal DNA-binding/nickase domain, a central domain displaying sequence motifs for helicase/ATPase, and a putative transactivation domain (TAD) at the C-terminus (21).
Researches implied that, with the help of the transcription factors Sp1/Sp3, NS1 N-terminal nuclease domain specifically binds to the origin of replication in the virus DNA, including the NS1-binding elements (NSBEs) and the overlapping P6 promoter DNA sequence, and the interaction between NS1 and virus DNA mediates the cleavage of DNA at the ITRs by melting the hairpinned ITRs to create a new 3′-OH and permit the following DNA synthesis (71,76,88). NS1 is not confined to its own viral promoter p6 to regulate gene transcription. It has also been identified in the transactivation of several host promoters, like tumor necrosis factor alpha (TNF-α), interleukin-6 (IL-6), and p21, which may explain the pathogenesis of B19-associated inflammation and apoptosis (19,54,62).
Studies on mechanism of B19-induced differential inhibition and apoptosis have made progress in recent years (8,30,55,96). B19 infection could induce a broad range of DNA damage responses (DDR) in ex vivo-expanded human EPCs, by triggering phosphorylation of all the upstream kinases of each of three repair pathways: ATM, ATR, and DNA-PKcs (48), which is of critical importance for virus replication. Also, the infected cells were reported to have a cell cycle arrest (CCA) at both late S phase and G2 phase, a point of the cell cycle at which cells contain 4N DNA (36,47,57). Recent advantages revealed that, NS1 transactivated cellular gene expression through the TAD2 domain, activating the ATR-CHK1-CDC25C-CDK1 pathway in the B19-induced G2 arrest (96), which is independent of p53 activation and DDR triggered by increased viral replication (36).
Interaction between NS1 and E2F family of transcription factors enhanced the nuclear import of these repressive E2Fs and induced stable G2 arrest (92). However, the putative metal coordination motif in the endonuclease domain of B19 NS1 is critical for NS1-induced S phase arrest and DNA damage (36). NS1 has also been reported to induce CCA at G1 phase in NS1-expressing UT7/Epo-S1 cells (57).
Replication of B19 virus promotes its NS1 covalently binding with host cellar DNA, causing DDR mediated by helicase or nickase in NS1 central region. Subsequently occurs the activating of the DNA nick repair pathway initiated by poly (ADPribose) polymerase and the DNA repair pathways initiated by ATM/ATR (69). In addition, the DNA repair processes activated by extensive DDR accompanied by a significant decrease in the ATP levels of the cell, act as the direct reason leading to apoptosis. Besides, NS1-induced DDR may play an indirect role in facilitating viral DNA replication by arresting cell cycle at G2 or S phase, during which host DNA replication factors are available (100).
Presumably, NS1 may also interfere with the expression of unidentified host transcriptional factors, resultantly perturbs cell cycle progression and inhibit the differentiation of EPCs. While S phase arrest enriches S phase factors that favor viral DNA replication as a compromised outcome of B19 genome replication, G2 arrest halts erythropoiesis of EPCs and eventually leads to apoptosis, in which 11 kDa protein may play a more efficient part (8,47,70).
In addition, the roles of NS1 in modulation of inflammatory signaling by activation of STAT3/PIAS3 and NLRP3, in inhibition of Na+/H+ exchanger activity, and in exacerbation of liver injury were also documented (22,32,37,89), which may partly explain the pathogenesis of multiple B19-associated diseases (Fig. 2).
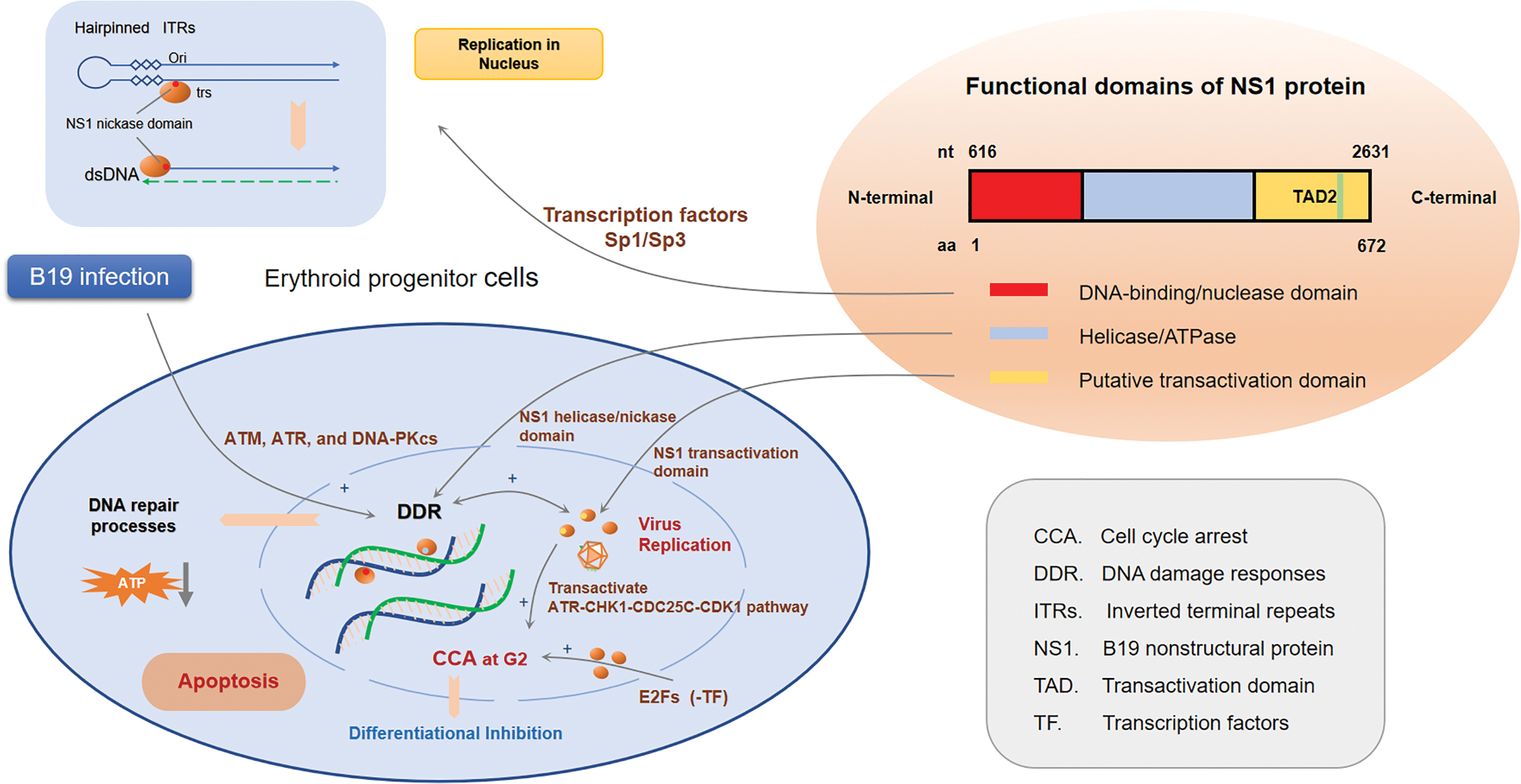
Roles of nonstructural protein 1 in B19 infection. Replication of B19 virus promotes its NS1 covalently binding with host cellar DNA, induces CCA at G1 phase in NS1-expressing UT7/Epo-S1 cells and causing DDR mediated by helicase and nickase in NS1 central region, resultantly perturbs cell cycle progression and inhibits the differentiation of EPCs. CCA, cell cycle arrest; DDR, DNA damage response. Color images are available online.
As the abundant expression of B19 NS1 is predominantly located in the nucleus of EPCs (92), the protein only takes on its activity during the replication of virus DNA, while not participating in the processes of assembling and release, and loses its function as the elimination of B19 virions, which is in accordance with the transient erythropoietic arrest in aplastic crisis. The time phase property of NS1 activity and conformity to transient change of hemogram implied that NS1 activity is closely related to differentiation and apoptosis of hematopoietic cells. Despite the enormous progress made in understanding the roles of B19 NS1-induced apoptosis, further studies are needed to explore the regulatory mechanism in NS1-induced inhibition of EPC differentiation, which is essential for therapeutic treatments of B19-related anemia. Besides B19, other virus such as Epstein/Barr virus, could also inhibit erythroid lineage cells differentiation by interfering certain signaling pathways involving in hematopoietic dysfunction. Among these pathways, B19 replication and transcription play an important role in Notch-Hes (hairy/enhancer-of-split)-GATA signaling regulation, which inhibit hematopoietic cell differentiation.
Transcriptional Factors Related to Hematopoiesis
The GATA family
GATA-binding proteins, known to be the erythroid-specific transcription factor family, have a evolutionary significance in understanding the development of precursors from hematopoietic stem and progenitor cells, the generation of red blood cells from progenitors, and the regulation of hemoglobin synthesis (32). All members of the GATA family have highly conserved DNA-binding proteins that recognize the motif WGATAR through two zinc fingers to regulate the transcription of downstream target genes (22,37,89). Millions of copies of the specific sequences of DNA in genome present upstream of gene transcriptional origin, such as promoters, enhancers, and locus control regions of β-globin and other genes, including itself as an autoregulatory mechanism (12,32). The C terminal zinc finger specifically binds to the GATA consensus sites, whereas the N terminal zinc finger stabilizes the interaction between GATA and specific DNA sequences as well as regulates the transcriptional activity of GATA factors through recruiting other cofactors of zinc finger protein, such as FOG1, CPB/p300, Pax5, and Pu.1 (7,27,63,86,90).
Among the six members (GATA1–6) of the GATA family, GATA1 is the founding member and ushered in the cloning of the related proteins GATA2–6, and both GATA1, 2, and GATA3 are expressed in specific hematopoietic cell types of all stages. Involved in distinct and overlapping aspects of hematopoiesis, the three members play an essential role in the development and maintenance of diverse blood cell lineages, and are indispensable for regulating the development and maturation of red blood cell (11,32).
GATA1 is a prototypical transcriptional factor required for the erythroid, eosinophilic, and megakaryocytic commitment during hematopoiesis, taking part in the terminal differentiation. GATA2, predominantly expressed in hematopoietic stem and progenitor cells, is essential for maintenance of the pool of hematopoietic stem cells by regulating the proliferation and survival of early hematopoietic cells, and is also one of the most critical transcriptional factors required for direct induction of the hemogenic endothelium with pan-myeloid potential from human pluripotent stem cells (18,74). Both GATA1 and GATA2 are involved in lineage-specific transcriptional regulation, especially the dynamic and strictly controlled GATA factor switching from GATA2 to GATA1 during erythropoiesis plays a crucial role in orchestrating erythroid lineage differentiation (56,85), whereas GATA3 is of vital importance for multiorgan development and regulates tissue-specific differentiation; it plays an essential role in T lymphoid cell development and immune regulation as well (22,23,78).
Therefore, alteration of GATA factors' expression is closely associated with hematologic disorders and related diseases (12,80). No or less GATA1 expression notably influences the differentiation and maturation of erythroid cells, and massive apoptosis of proerythroblasts leads to anemia (12). Besides, GATA1-deficient mice develop thrombocytopenia and hyperproliferation of megakaryocytes due to dysmaturity of megakaryocytes and a failure of platelet production (91). GATA2 mutation is associated with immunodeficiency, lymphedema, and myelodysplastic syndrome (29,77,82). Inherited GATA3 variants are related to Ph-like childhood acute lymphoblastic leukemia and risk of relapse (67).
Researches have revealed some correlations between GATA family and the Notch signaling pathway, which is also involved in the hematopoietic system (14,31,50,75,87). It was found that Notch signals could inhibit the differentiation and maturation of erythroid/megakaryocytic cells by suppressing GATA-1 activity through the induction of Hes1 expression (31).
The Notch-Hes pathway
Notch proteins are a family of evolutionarily highly conserved single-pass transmembrane receptors, which are involved in the regulation of cell fate acquisition and differentiation in diverse systems. The notch signaling pathways not only play an essential part in the development of a wide range of tissues, such as hemopoiesis, vasculogenesis, myogenesis, neurogenesis, and osteogenesis, but also take part in the homeostasis maintenance of a broad variety of tissues (1,81). The family comprises of four Notch receptors (Notch1–4), five structurally related, single-pass membrane Notch ligands (Delta-like1, 3, and 4 and Jagged1 and 2), and specific factors, including the DNA-binding protein RBP-Jκ [recombinant-binding protein suppressor of hairless; also known as CSL/CBF1 in mammals, Su(H) in flies, and Lag-1 in worms], and the Mastermind-like family (MAML) (38,93).
In the absence of Notch activation, the Notch intracellular domain (NICD) is unavailable and the downstream effector protein RBP-Jκ associates with several different corepressors containing Mint/Sharp/SPEN, NCoR/SMRT, and KyoT2 to form a transcriptional corepressor complex which is bound to the chromatin and inhibits gene expression. Activation of the canonical Notch signaling is achieved by the generation of NICD, which is mediated by the interaction between receptors and ligands and subsequently a sequence of proteolytic events, as well as its eventual translocation to the nucleus, where the RBP-Jκ association module (RAM) domain of NICD initially binds the RBP-Jκ. This leads to the displacement of the corepressor complex and the recruitment of the transcriptional coactivators like MAML to form a transcriptional activator complex (NICD-RBP-Jκ-MAML), which triggers the downstream expression gene expression by recruiting transcriptional factors like p300 histone acetyl-transferase (1,2,38,43,81,93).
The various target genes of Notch, including Hes and the Hes-related (HESR/HEY) family of basic helix-loop-helix transcription repressors, are essential regulators of hematopoietic stem cell development, and subsequently modulate the proliferation and differentiation through regulating expression of other genes like GATA family (25,39). In addition, GATA2 was also identified as a direct target of Notch1 signaling, which revealed a crucial role of Notch activation for the onset of definitive hematopoiesis in the embryo (72).
Studies have identified Notch signaling as a key regulator of hematopoietic stem cell development (2,17,24,34,43,72,97). Among the Notch and Hes family members, Notch1, 2 and Hes1, 5 are widely expressed in all lineages of hemopoietic stem/progenitor cells, and participate in regulating their proliferation and differentiation to generate various hemocytes complying with extremely strict principle of spatiotemporal sequence mediated by the fine expression of GATA factors (2,25,31,39,43,72). Activation of Notch1 signaling could inhibit the differentiation and maturation of EPCs and exhibit a peripheral hemogram of increased immature red blood cells and distinctly decreased counts of mature red blood cells (9,35,39), which might be a clue of building a bridge between B19 infection and anemia.
The Pathway of B19 NS1 in Regulation of EPCs Differentiation
Even though limited research is available to clarify NS1 inhibition in the process of proliferation and differentiation through manipulating Notch signaling in B19-infected EPCs, evidences of the connection to Notch have been found in other virus-infected cells (26). Epstein/Barr virus nuclear antigen 2 (EBNA2) exerts its transactivating function through interaction with CBF1/RBP-Jκ, which is the coactivator of Notch signaling (99). Notch signaling is involved in the establishment of EBV latency in B cells possibly due to competitive binding of EBNA2 to CBF1/RBP-Jκ and the suppression of Notch/RBP-J signaling pathway, which promotes B cell proliferative responses (26,40). In addition, RTA, the lytic cycle regulator of Kaposi's sarcoma-associated herpesvirus (KSHV), also interacts with RBP-Jκ to activate gene expression (45). Furthermore, Notch pathway interactions have also been mentioned for adenovirus SV40 and human papilloma virus (3,61).
It has been identified that B19 could inhibit the differentiation of erythroid lineage cells both in vivo and vitro mediated by its major pathogenic factor NS1 (70,96). The Notch signaling pathway may play an essential part in hemopoiesis through regulating its downstream genes like Hes1/5 and GATA factors (2,25). Our tentative exploration showed that expression of Notch1, Hes1/5, and GATA2 is upregulated, whereas GATA1 is downregulated in the B19 NS1-transfected K562 cells, which implies that NS1 could perturb the differentiation of erythroid lineage cells through manipulating Notch signaling, leading to alteration of expressional patterns of target gene Hes and GATA factors (unpublished data). The crosslink provides a new insight of the potential mechanism of B19-induced differential inhibition of EPCs. Further studies are needed to explore the expressional alteration of related target genes and the concrete regulatory pathway of Notch signaling to have a clearer understanding of the pathogenesis of B19-related anemia (Fig. 3).
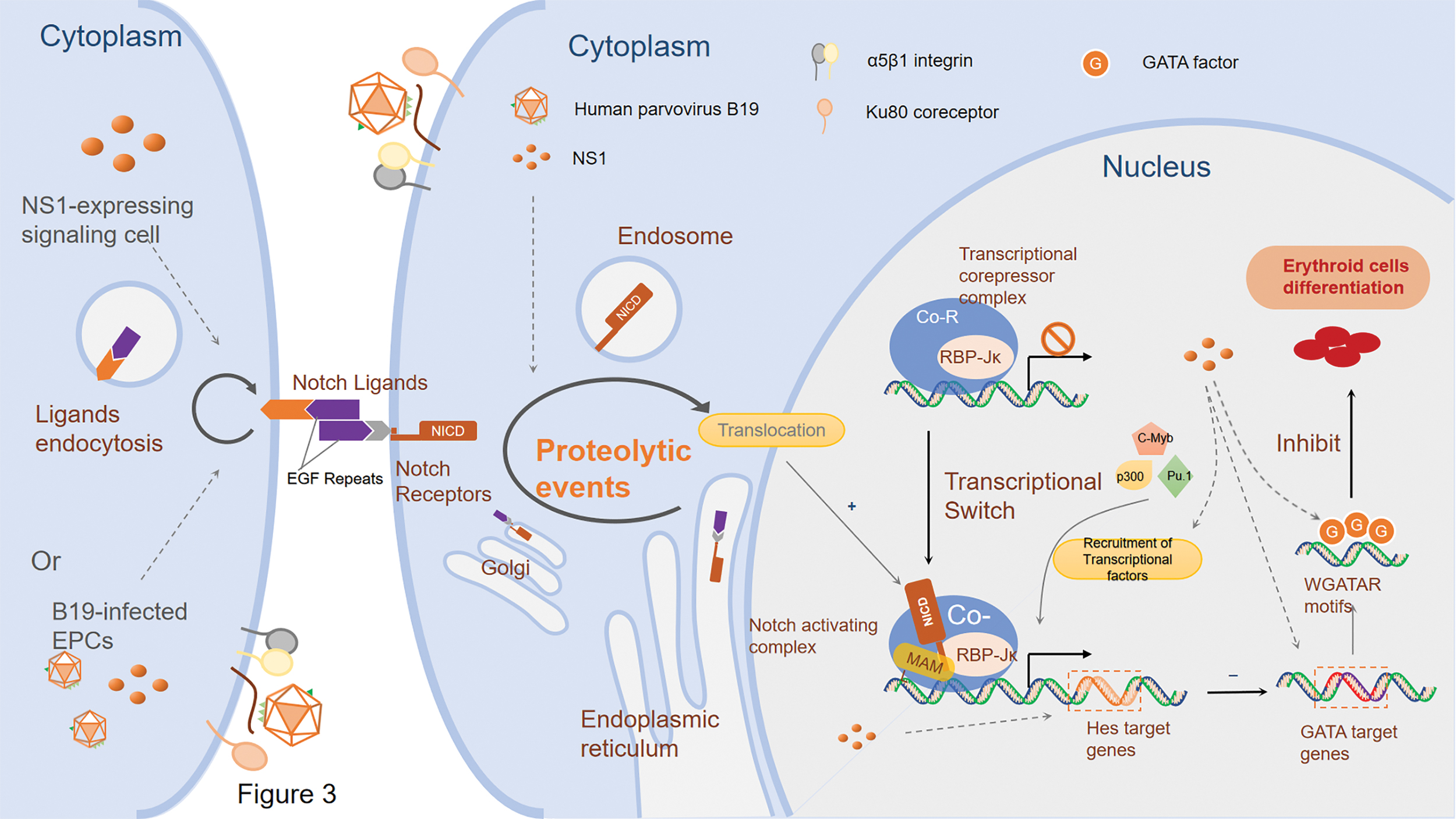
Putative pathways of nonstructural protein 1 in the regulation of EPCs differentiation. B19 NS1 upregulates the expression of Notch1, Hes1/5, and GATA2, whereas downregulates GATA1, which perturbs the differentiation of erythroid lineage cells through manipulating Notch signaling, leading to alteration of expressional patterns of Hes and GATA. Hes, hairy/enhancer-of-split. Color images are available online.
Conclusion
In summary, in this review, we provide a new insight of the bridge between B19 infection and Notch signaling pathway or transcriptional factors GATA: B19 NS1 could perturb the differentiation and proliferation processes of erythroid lineage cells through manipulating Notch signaling, leading to alteration of expressional patterns of target gene Hes and GATA factors. The crosslink provides a new potential mechanism of B19-induced differential inhibition of EPCs, which may also give a clue to prophylactic and therapeutic targets for B19-related severe anemia in high-risk groups, and develop effective vaccines or antivirus drugs of B19 infection.
Statement of Ethics
The research was conducted ethically in accordance with the World Medical Association Declaration of Helsinki.
Footnotes
Authors' Contributions
S.F. collecting references, drawing the figures, and writing this article; D.Z. and J.Z. participated in the writing and discussion; and D.Z. in writing and organizing this article.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This work was supported by a grant from the Chinese National Natural Science Fund 81170005 and 81670007.