
Abstract
Henipaviruses are single-stranded RNA viruses that have recently emerged as zoonotic pathogens, capable of causing severe acute respiratory disease and encephalitis in humans. The prototypical henipaviruses, Hendra henipavirus and Nipah henipavirus, are a major health concern as they have high mortality rates and no currently approved human vaccine or drug therapy. Understanding the mechanisms of viral replication and pathogenicity is of critical importance for therapeutic developments. A novel target for such therapies is the Henipavirus Matrix (M) protein, a multifunctional protein that drives viral assembly and inhibits the innate immune response. These multifunctional attributes promote a complicated lifecycle: while viral replication occurs in the cytoplasm, M traffics to the nucleus, where it is ubiquitinated, for correct cellular targeting and virion packaging. In this study, we review the relationship between the structure and functions of M. In specific cases, the compatibility between structural accessibility and protein functionality is not always evident, and we highlight areas that require further investigation.
Introduction
Henipaviruses are nonsegmented, negative-sense RNA viruses that contain a host-derived membrane. The virions are variable in size, but on average can be described as pleomorphic to circular, and ∼300 nm in diameter (22,44,47). The Henipavirus genus belongs to the Paramyxoviridae family that contains many medically significant viruses, including measles virus, mumps virus, human parainfluenza viruses, and others, which greatly impact animal species such as Sendai virus and Newcastle disease virus (NDV).
Henipavirus genomes, however, differ by being significantly larger compared with most other paramyxoviruses (68). The genome encodes six structural proteins: the attachment (G) protein, matrix (M) protein, phospho (P) protein, fusion (F) protein, nucleo (N) protein, and polymerase (L). The P gene encodes P protein and three nonstructural proteins W, V, and C. W and V proteins are generated through an RNA editing site, and C is translated through different open reading frames (18,33).
Henipaviruses are recently emerged zoonotic pathogens that cause acute respiratory disease and lethal encephalitis in animals and humans (43), with an average mortality rate ranging from 40% to 75%, but as high as 100% in some outbreaks (70,73). The two prototypical viruses from this genus, Hendra henipavirus (HeV) and Nipah henipavirus (NiV), have been isolated from Pteropus bats (2,25). There is an absence of disease in Pteropus bats, which are a natural reservoir for HeV and NiV (19,24). The intermediate hosts involved differ between the viruses, as HeV infects horses and NiV was originally transmitted to humans from infected pigs (2,4). Other henipaviruses have been described and include Ghanaian bat henipavirus (GhV), Mojiang henipavirus (MojV), and Cedar henipavirus (CeV). NiV is the only henipavirus that is known to be transmitted directly from bats to humans, and from human to human (35).
Due to the high risk posed by HeV and NiV, they are classified as Biosafety Level 4 pathogens, requiring the highest level of containment for research involving live viral particles, which contributes to the slow progress in the development of effective drug treatments and vaccines against HeV and NiV. A greater understanding of the molecular mechanisms henipaviruses use to replicate their genomes, assemble infectious virions, and antagonize host defenses is urgently needed to inform the development of novel therapeutics against these pathogens.
Emergence, transmission, and symptoms of henipaviruses
The first outbreak of HeV occurred in the suburb of Hendra, Queensland, Australia, in September 1994 (45,60). The symptoms of HeV infection are acute respiratory disease and neurological manifestations, including encephalitis. Since the initial outbreak, there have been 85 confirmed and 20 unconfirmed equine cases of HeV, and by July 2020, the latest confirmed case of HeV was near Murwillumbah, NSW, Australia in June 2020 (15,46). To date, there have been seven human cases of HeV with four deaths and no case of bat to human transmission. Although Field et al. (19) demonstrated that HeV can be excreted in flying fox urine throughout the year in variable locations, the presence of virus alone does not determine spillover into horses and postulate that environmental and host factors play a role in infection.
The initial outbreak of NiV occurred in Malaysia and Singapore (NiV-MY strain) in 1998–1999, where 265 human cases of encephalitis were recorded, resulting in 105 deaths (4,9,10). The spillover of virus from flying foxes into farmed pigs facilitated the transmission to humans in pig farms, and as a result, over 1 million pigs were culled to contain the disease (34). A second strain of NiV emerged in Bangladesh and India in 2001 (NiV-BD) (26,27), infecting humans without the requirement of pigs to act as an intermediate host. NiV-BD is acquired through ingestion of contaminated date palm sap and >50% of cases resulted from person to person transmission (35). Since its emergence in Bangladesh, yearly outbreaks have occurred. The most recent outbreak of NiV saw six people infected, including four deaths in Bangladesh in January, 2020 (1). NiV infection presents as fever and severe respiratory disease followed by dissemination throughout the central nervous system, resulting in neurological symptoms and encephalitis (4).
Since the characterization of HeV and NiV, other zoonoses have emerged, which have revealed similar viral agents that have been classified in the Henipavirus genus. CeV was isolated from bats from Queensland, Australia, although it has been deemed nonpathogenic (30,39). MojV was discovered in 2012 by amplifying and sequencing viral RNA obtained from rats (Rattus flavipectus), after three cave miners in China died from viral pneumonia (71). GhV has been sequenced from straw-colored fruit bats (Eidolon helvum) in Africa, and there is serological evidence that it has spilled over into humans (41). While it is unknown if it causes disease, it has been suggested that GhV may be the etiological agent for undiagnosed encephalitis (52). Both MojV and GhV are yet to be isolated from humans to confirm pathogenicity (58). The Henipavirus genus is of great concern as further disease spillover events could result in widespread disease.
M protein
The M protein is a multifunctional protein that is common to Paramyxoviridae species. M is essential for viral assembly and budding, accumulating in inclusion bodies below the plasma membrane during late stages of infection (58), before incorporating F and G proteins into the membrane and associating with the ribonucleoprotein for egress of the virion at the apical cell surface (31). HeV-M has been shown to assemble with the N–P–L complex of NiV to coordinate budding in a chimeric viral analysis (74), demonstrating the high similarity of the M proteins and anticipated conserved structural features between the species. While virus-like particles (VLPs) can assemble in M-deficient expression studies, the infectivity and stability of these particles are reduced, suggesting a key role of M in the correct assembly of the virion (16). Johnston et al. examined the role of F and M in the assembly of VLPs and established that F is also a strong modulator of assembly, relying on cellular actin and vesicular trafficking proteins to facilitate VLP formation (28). In addition to the structural functions, M is involved in modulating the host's innate immune response (8). Post-translational addition of monoubiquitin occurs within the host cell nucleus to regulate its function through enabling interaction with several host proteins to coordinate pathogenesis (8,69).
M localizes to the nucleus
Paramyxoviruses (like most RNA viruses) carry out replication in the cytoplasm, yet several species, including NDV (50), HeV, and NiV, target the viral M protein to the nucleus. McLinton et al. (42) examined and compared the nuclear-cytoplasmic trafficking of M proteins of HeV, NiV, CeV, GhV, and MojV, and another Paramyxovirus, Measles morbillivirus (MeV). While the green fluorescent protein (GFP) control produced a diffuse pattern throughout the cytoplasm and nucleus, GFP-tagged Henipavirus M proteins were localized in puncta within the nuclei and nucleoli and appeared to translocate to the plasma membrane 18–22 h post-transfection in both human and bat cell lines (with the exception of GhV-M, which did not translocate to the plasma membrane). In contrast, GhV and MeV M were mostly localized to the cytoplasm, while a small proportion could be detected within the nucleus.
Numerous studies have performed functional and interactome analyses of Henipavirus M in an attempt to elucidate key motifs and binding partners involved in the nuclear trafficking process (7,8,14,32,42,48,51,54,69). In an attempt to identify host cell regulatory factors of HeV and NiV M, Pentecost et al. (51) tagged M with 3X-Flag and expressed the protein the human cell line HEK 293, followed by purification and mass spectrometry to identify co-purified host proteins. Among the protein interactome of HeV-M were Nuclear Pore Complex proteins, Importin-α (IMPA) isoforms 3,4,5,6,7, and nuclear exporters chromosomal maintenance 1 (Exp1/CRM-1) and exportin 2 (Exp2). NiV-M interactome revealed IMPA isoforms 1,3,4,5,6,7 giving further evidence that Henipavirus M undergoes nuclear localization through the classical pathway (Fig. 1). Further co-immunoprecipitation and immunoblotting revealed interaction of NiV-M with IMPA isoforms, with the strongest interaction occurring with IMPA6 and IMPA7. Martinez-Gil et al. (40) also analyzed the NiV protein interactome using mammalian expression vectors, affinity purification, and mass spectrometry. Several nuclear proteins were identified as interaction partners of M, including ANP32B, ANP32E, HUWE1, RAD18, and TRBP7; however, no nuclear import receptor was identified (40).
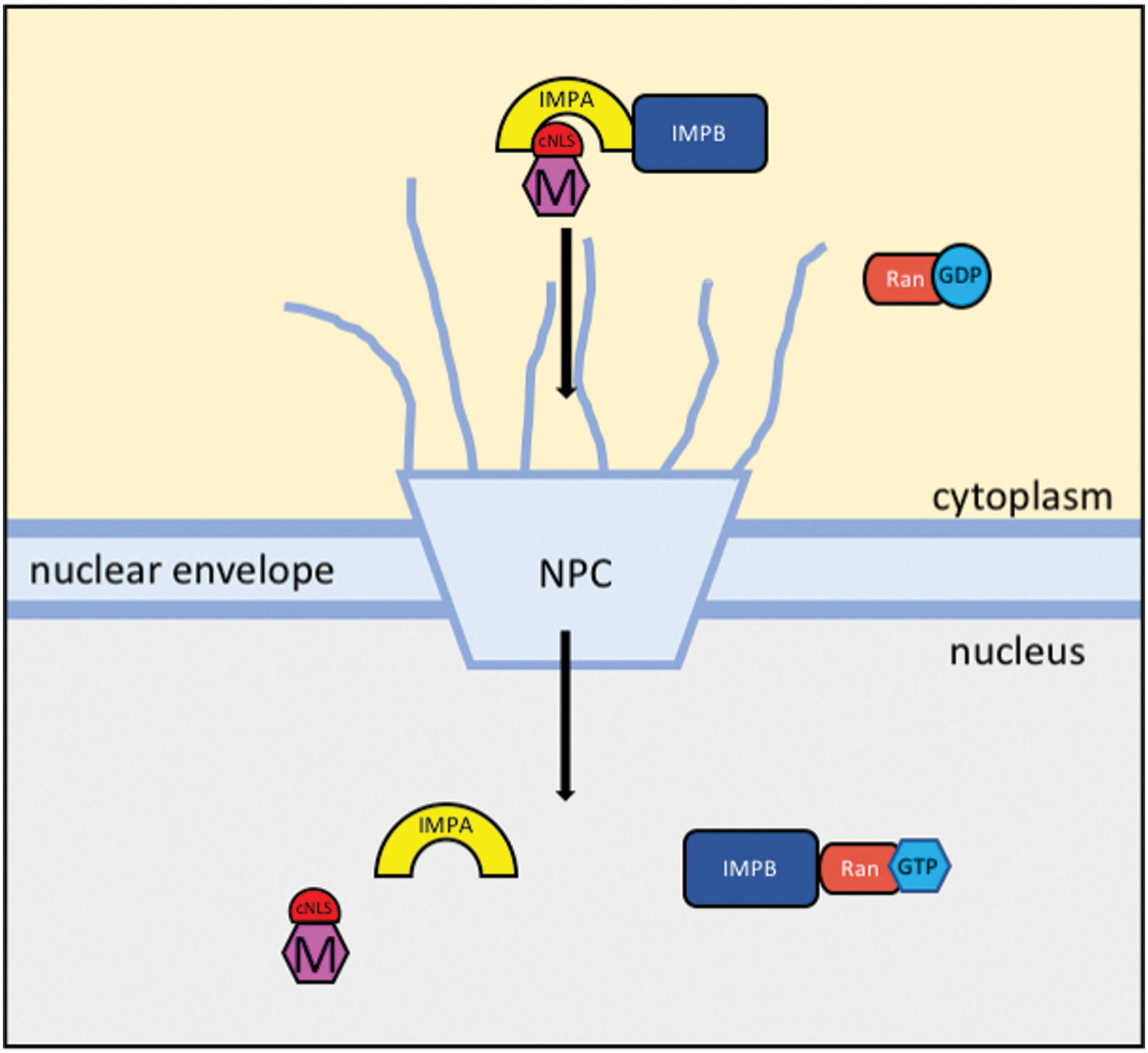
Schematic representation of the proposed nuclear import pathway of HeV and NiV M protein. Cargo proteins contain cNLS for binding to the adaptor protein IMPA for translocation through the NPC by IMPB. IMPB binds to Ran-GTP within the nucleus and dissociates IMPA and the cargo protein. NLS, nuclear localization signal; cNLS, classical nuclear localization signal; IMPA, Importin-α; NPC, nuclear pore complex; IMPB, Importin-β; HeV, Hendra henipavirus; NiV, Nipah henipavirus; M, matrix. Color images are available online.
The interactions of M with nucleolar proteins have also been explored. Immunoprecipitation and mass spectrometry identified the nucleolar protein Treacle as a major interaction partner of M (54). Treacle is a regulator of rRNA synthesis in the DNA damage response, and Rawlinson et al. (54) established that silencing Treacle with siRNA before HeV infection resulted in a significant increase in viral production. The authors interpret their findings as a novel mechanism by which the Henipavirus M can modulate the host cell's environment to support viral production, despite the DNA damage caused by infection (54).
Deffrasnes et al. (14) performed a genome-wide siRNA screen to identify host proteins that are required for Henipavirus infection and identified the nuclear and nucleolar protein fibrillarin as essential. Curiously, they demonstrated that fibrillarin co-localizes to the nucleolus with M early in Henipavirus infection. Although fibrillarin was not required for nuclear import of M, fibrillarin knockdown resulted in inhibition of HeV infection with reduced budding efficiency (16). The function of this association is not yet understood and further investigation into the nuclear functions of M and a potential link to fibrillarin are required.
Nuclear export of M is also essential for viral assembly and budding, with L106/107A mutations (within the nuclear export signal) resulting in nuclear accumulation and failure of M to form inclusion bodies beneath the plasma membrane, a precursor to viral budding (56,69). Interaction and nuclear accumulation of HeV and NiV M with ANP32B, a CRM-1 nuclear export adaptor, were documented by Bauer et al. (7) and ANP32B has been further established by Gunther et al. (23) as a common nuclear target for paramyxoviral M proteins. The role of the nuclear protein interactions with M remains unclear; nuclear trafficking is crucial for regulating correct assembly and budding, and it is proposed that M performs nuclear functions to modify host immune pathways. Further structural investigations may reveal novel mechanisms of virus:host regulation that contribute to pathogenesis.
After nuclear transit, M recruits the cellular machinery of the endosomal trafficking system to target protein toward the plasma membrane for assembly and budding. Sun et al. (64) identified the trafficking protein AP-3 beta subunit (AP3B1) as a binding partner of HeV and NiV M through co-purification. The authors mapped the binding region to the serine-rich acidic Hinge domain of AP3B1; however, the binding region of M is yet to be described. Depletion of AP3B1 by siRNA reduced the VLP production threefold in cells transfected with NiV-M plasmid (64). Synthesis and co-expression of an AP3B1 hinge region peptide with NiV-M protein saw a 20-fold reduction in VLP production, with similar results for HeV-M. These data indicate that the hinge region peptide was able to bind M and interfere with cellular AP3B1 binding, abrogating cellular trafficking processes and reducing VLP budding capacity. Further investigation and structural analysis of the M binding domain could reveal a target for therapeutic peptides to target the cellular trafficking of Henipavirus proteins during infection.
Role of ubiquitination in regulating function
Wang et al. (69) reported that nuclear import, ubiquitination, and export of NiV-M is essential for the assembly and budding of the virion. Ubiquitin is a small protein modifier, which is covalently attached to lysine residues within eukaryotic cells as a signal for degradation at the proteasome, as a change in subcellular localization, or to alter a proteins' affinity for a binding partner (3). Wang et al. (69) transfected HeLa cells with GFP-NiV-M, treated with proteasome inhibitors MG132 and bortezomib to deplete free cellular ubiquitin, and then analyzed cellular localization using fluorescence microscopy. This resulted in NiV-M being retained in the nucleus and impeded budding, whereas overexpression of ubiquitin enhanced budding. Furthermore, immunoprecipitation with an antibody specific for ligated, monoubiquitin modifications was able to specifically co-precipitate NiV-M. A lysine residue at position 258 was identified as the site of monoubiquitination. This residue also resides within a putative nuclear localization signal (NLS) (discussed in later section “Nuclear localization signals”). Mutagenesis revealed that NiV-M K258R (lacking a lysine residue for ubiquitination, but retaining the positive charge) was retained in the nucleus, while K258A NiV-M mutant was excluded from the nucleus, indicating a positively charged residue at position 258 is required for nuclear localization. Both mutants were immunoprecipitated with significantly reduced levels of monoubiquitin than wild type, and mutants also failed to translocate to the plasma membrane for budding, indicating that ubiquitination of the key residue K258 is important for nuclear export, membrane association, and viral budding. This highlights the critical importance of nuclear trafficking of Henipavirus M in the role of assembly and budding of the virion. Whether these K258 mutations alter the interaction of M with host proteins such as ANP32B or fibrillarin remains to be investigated.
Type 1 interferon antagonism
M is the first Henipavirus structural protein to be described as having type 1 interferon (IFN-I) antagonism (8). Upon release into the host cell cytoplasm, and before de novo translation of viral transcripts, M protein blocks the host antiviral response. M can block the host antiviral response in the early stage of infection to allow for the virus to replicate and egress, evading detection. NiV is known to evade the host antiviral response through M antagonism of IFN-I production by interaction with (and degradation of) the host E3 ubiquitin ligase TRIM6 (8). This interaction reduces the production of K48-linked polyubiquitin chains that activate the IFN-inducing kinase inhibitor of κB kinase ɛ and so blocks the downstream components shared by several antiviral response pathways (53).
Bharaj et al. (8) determined that the NiV-M K258A mutant was excluded from the nucleus in HeLa cells and induction of antiviral antagonism was reduced. The authors postulated that this was due to loss of M protein K258 monoubiquitination (occurring in the nucleus) that was required for subsequent export from the nucleus and promotion of TRIM6 interaction. Considering these results [together with the work of Wang et al. (69)], it is likely that M requires nuclear localization to undergo ubiquitination for correct cellular sorting and viral assembly, and perhaps regulation of IFN-I antagonism. Understanding the nuclear translocation process of M is an important step for viral replication and assembly and is discussed in section “Nuclear localization signals.”
Structural Analysis of M
The structure of HeV-M was solved using x-ray crystallography (32), providing important insights into the conformation of the functional regions that mediate assembly, post-translational modifications, and subcellular localization. The structure also raises some uncertainty about the accessibility of ascribed regions for functionality. The structural features of M driving the viral:host interactions are yet to be confirmed using structural biological methods.
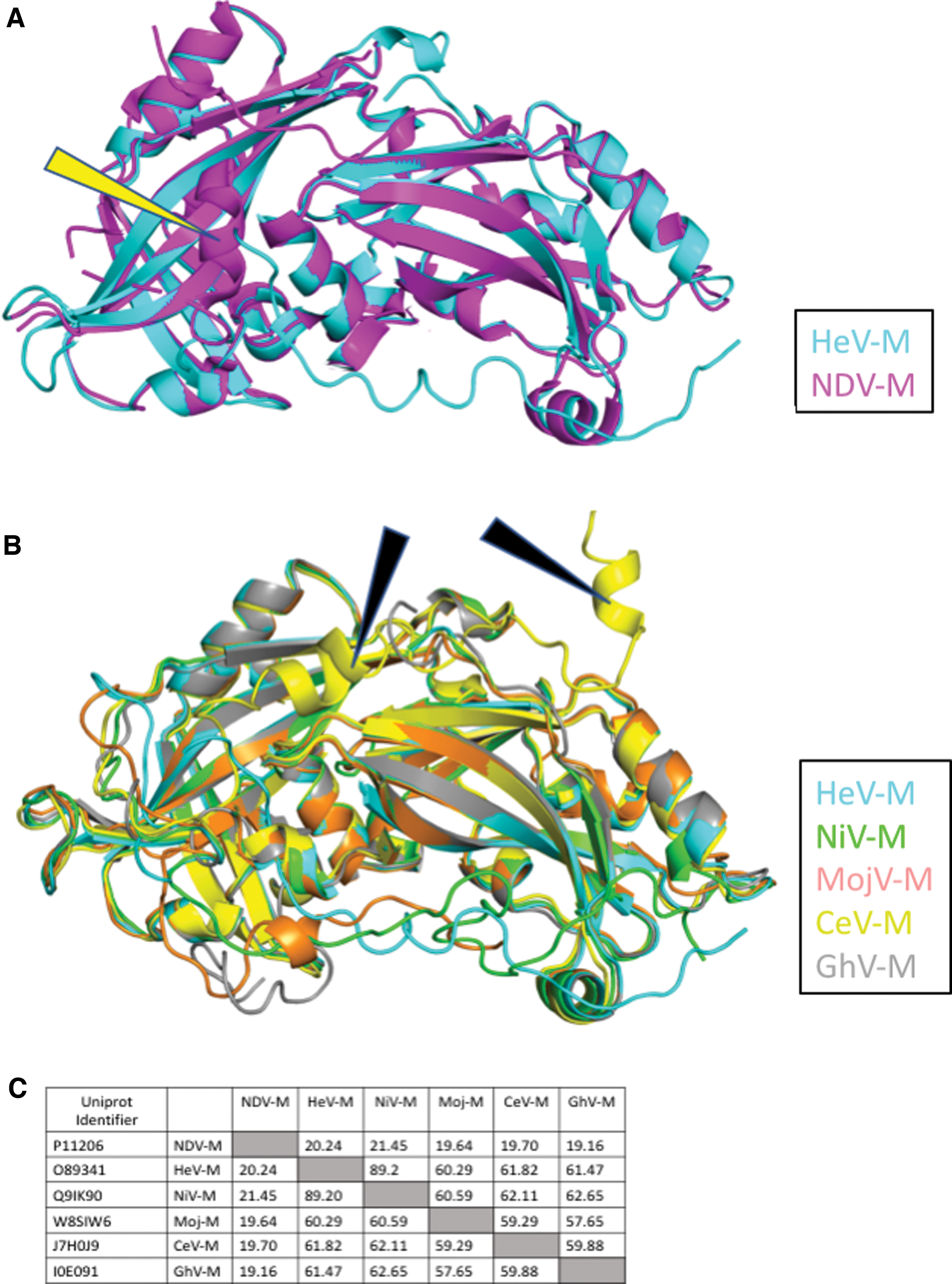
HeV-M protein consists of 352 amino acids in a single chain and comprised two similar β-sandwich core domains surrounded by α-helices (Fig. 2). The M protein forms a homodimer through a hydrophobic core, polar contacts, and two salt bridges. Sequence alignment of HeV-M with other henipaviruses (36) reveals many conserved sites (Fig. 3), and modelling of these structures illustrates an anticipated similar tertiary structure (29). Despite 20% sequence identity, HeV M is strikingly similar in tertiary and quaternary structure to NDV M, another Paramyxovirus (root mean square deviation of 1.7 Å over 255 residues) (Fig. 4). M protein from paramyxoviruses oligomerizes and self-assembles into VLPs when expressed alone (note that these VLPs are distinct from those mentioned in section “M protein” and consist solely of an M protein shell at the internal face of a secreted, enveloped particle). These VLPs are generated through homodimerization and dimer-dimer interactions of M (6,32).
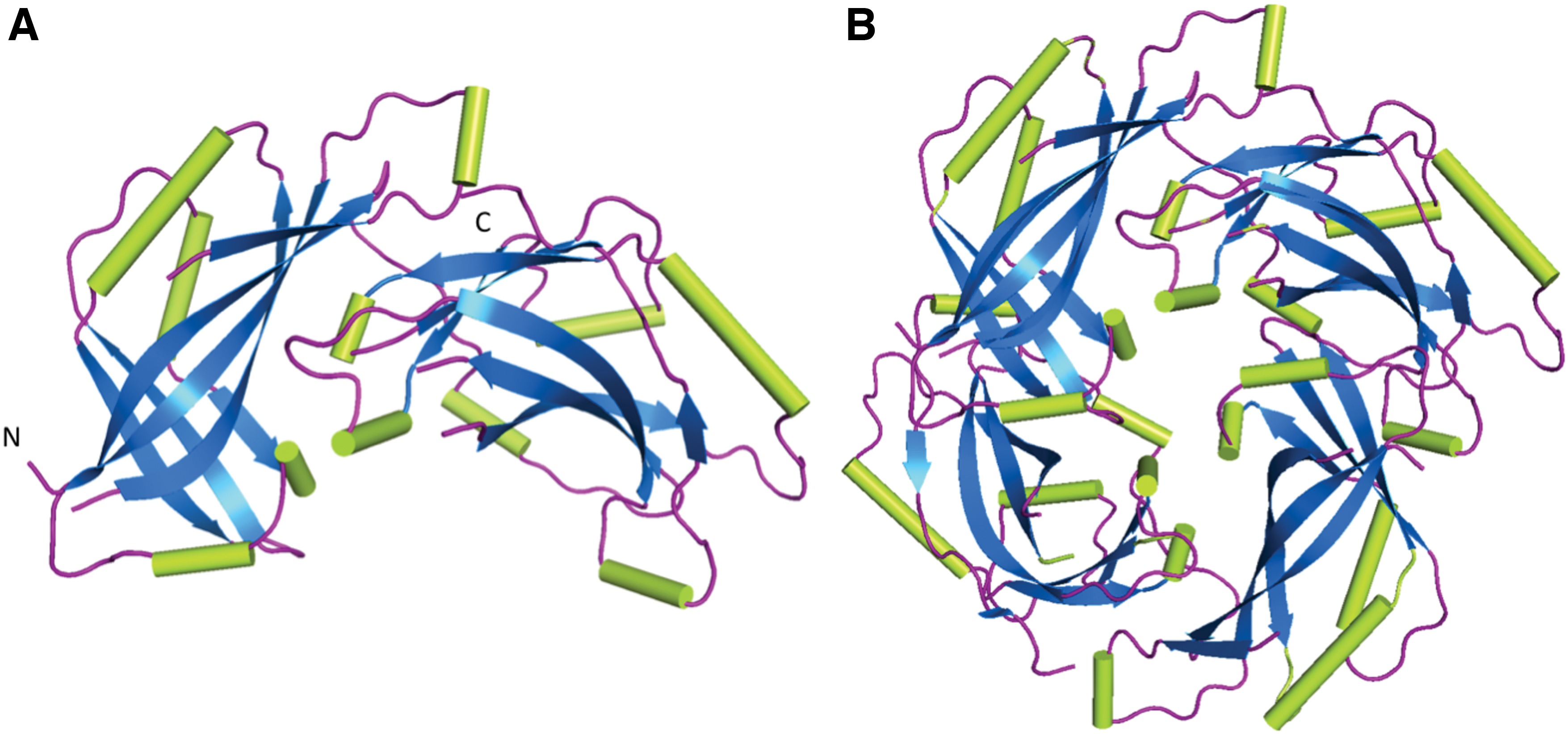
The structure of HeV-M PDB 6BK6 (32) in cartoon representation.

Multiple sequence alignment of M proteins of henipaviruses and NDV (61). Protein sequences obtained from UniProt (65) NDV-M (P11206), MojV-M (W8SIW6), CeV-M (J7H0J9), GhV-M (I0E091), HeV-M (O89341), and NiV-M (Q9IK90). Putative functional regions of NiV-M/HeV-M are highlighted along with corresponding regions of aligned proteins. There is high conservation across these regions within the Henipavirus genus with low sequence identity with NDV-M. The dimerization contacts (green) reveal a conserved R57 residue within the Henipavirus genus that is absent in NDV-M. Dimer-dimer interactions (blue), L domains (orange), and NES (purple). The mpNLS (pink) is conserved between HeV and NiV with reduced similarity in other species. The henipavirus bpNLS (yellow) is conserved at 244RR with positions 256–257 being strictly positively charged residues with the exception of GhV 256S. 258K is conserved across all species and lies within the nonclassical bpNLS of NDV-M (gray). bpNLS, bipartite nuclear localization signal; GhV, Ghanaian bat henipavirus; MojV, Mojiang henipavirus; CeV, Cedar henipavirus; NDV, Newcastle disease virus; mpNLS, monopartite nuclear localization signal; L, late; NES, nuclear export signal. Color images are available online.
The basic unit of HeV-M is the homodimer (also present in the crystal structure), which oligomerizes to form a grid-like array beneath the host plasma membrane, and there are several regions of the single protein chain that interact to establish a stable dimeric structure. The center of the interface between the two protein chains is held together by hydrophobic interactions between V154 and R158 located on the α5 helix of one monomer, and P307 and W308 located on the α12 helix, the adjacent monomer. While arginine is not generally considered a hydrophobic residue since it contains a positively charged guanidino group, the long aliphatic side chain can contribute to hydrophobicity if the charged group is extended toward the exterior of the protein (5), and this occurs at the dimer interface of the M protein.
Electrostatic forces drive the quaternary structure and comprised a salt bridge between the positively charged side chain of R322 and the negatively charged side chain of E166, and hydrogen bonds between polar side chains R118 and T120 from one M monomer and D340 and Q328 from the adjacent monomer. These residues at the dimer interface were shown to be important for M protein assembly. Liu et al. (32) analyzed the VLP assembly of wild-type and mutant HeV-M and demonstrated that changes in electrostatic interactions reduced dimer stability and VLP production (Table 1).
Summary of Mutagenesis Effects of Hendra henipavirus-Matrix Protein at the Dimeric Interface
Source: Liu et al. (32).
HeV, Hendra henipavirus; M, matrix; VLP, virus-like particle.
Liu et al. (32) performed structural analysis of HeV-M using the PDBePISA server to identify dimer-dimer contacts. In this study, it was found that the dimer-dimer interface is mediated by the α1 and α2 helices of adjacent dimers, at a 16° angle, which is consistent with the curvature of the membrane in which the M lattice is underlying. This also produces a positively charged surface that is required for interaction with the negatively charged heads of phospholipids on bilayer membranes. These interactions are facilitated by the interactions outlined in Table 2, with bonding occurring in reciprocal directions (Fig. 5). Site-directed mutagenesis of R57D and R57E resulted in abrogation of VLP formation, while charge reversal within the α2 helix was less impactful, with D105R producing negligible change and E108R reducing VLP production by ∼50%. The impact of these mutations on the interaction of M with host proteins and on cellular trafficking of M remains to be explored.
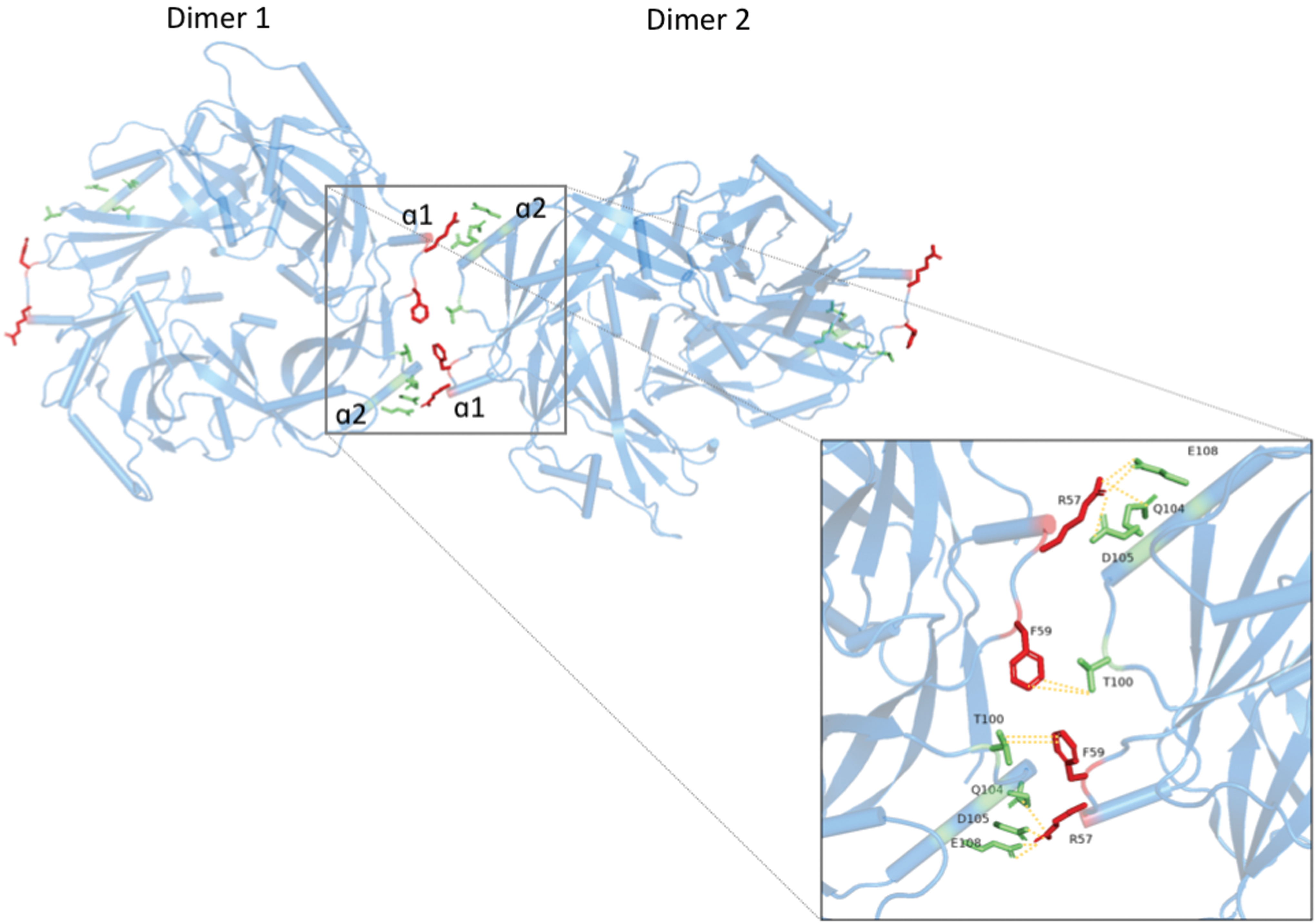
Dimer-dimer interactions of HeV-M (32), which are required for oligomerization and VLP formation. Dimers interact through the α1 helix (R57, F59) and the α2 helix (T100, Q104, D105, E108). Color images are available online.
Interactions of the Hendra henipavirus-Matrix Dimer-Dimer Interface as Described by Liu et al.
Source: Liu et al. (32).
Interactions are listed from dimer 1 to dimer 2; however, an additional set of identical interactions occurs from dimer 2 to dimer 1. Sequence alignment reveals that HeV-M R57, and the charged side chains within the α2 helix are conserved within the genus Henipavirus.
Similarly, NDV-M is dimeric in the crystal structure. Battisti et al. (6) performed cryoelectron tomography with three-dimensional reconstruction and x-ray crystallography to determine the structure of NDV-M and VLPs. This study suggested the dimer-dimer interaction is mediated by the α9 and α2 helices of adjacent dimers (Fig. 6), and therefore differs from the proposed arrangement of HeV-M. The R57 residue, conserved in the henipaviruses, is not present in NDV-M (Fig. 3). HeV-M lacks an α9 helix; however, the α8 helix is homologous with the NDV-M α9. Battisti et al. do not describe the residues that mediate this oligomerization; however, this arrangement also produces a positively charged surface of the M lattice for membrane interaction.
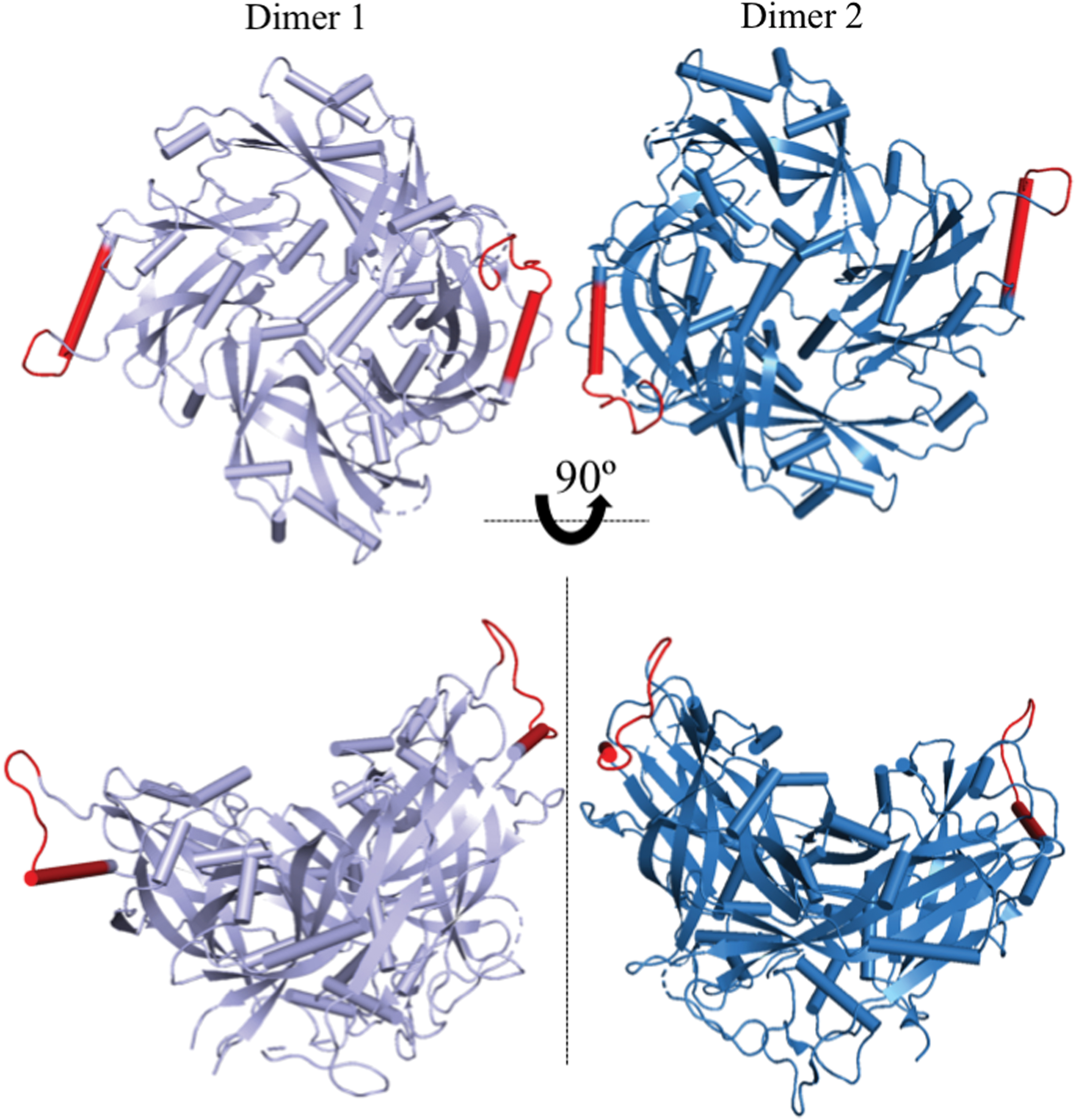
Oligomerization of NDV-M (PDB 4G1G) (6). NDV-M forms an oligomer through dimer-dimer interactions facilitated by the α9 and α2 helices. Interaction at the α9 helices (red) is shown in cartoon representation. Color images are available online.
Late domains
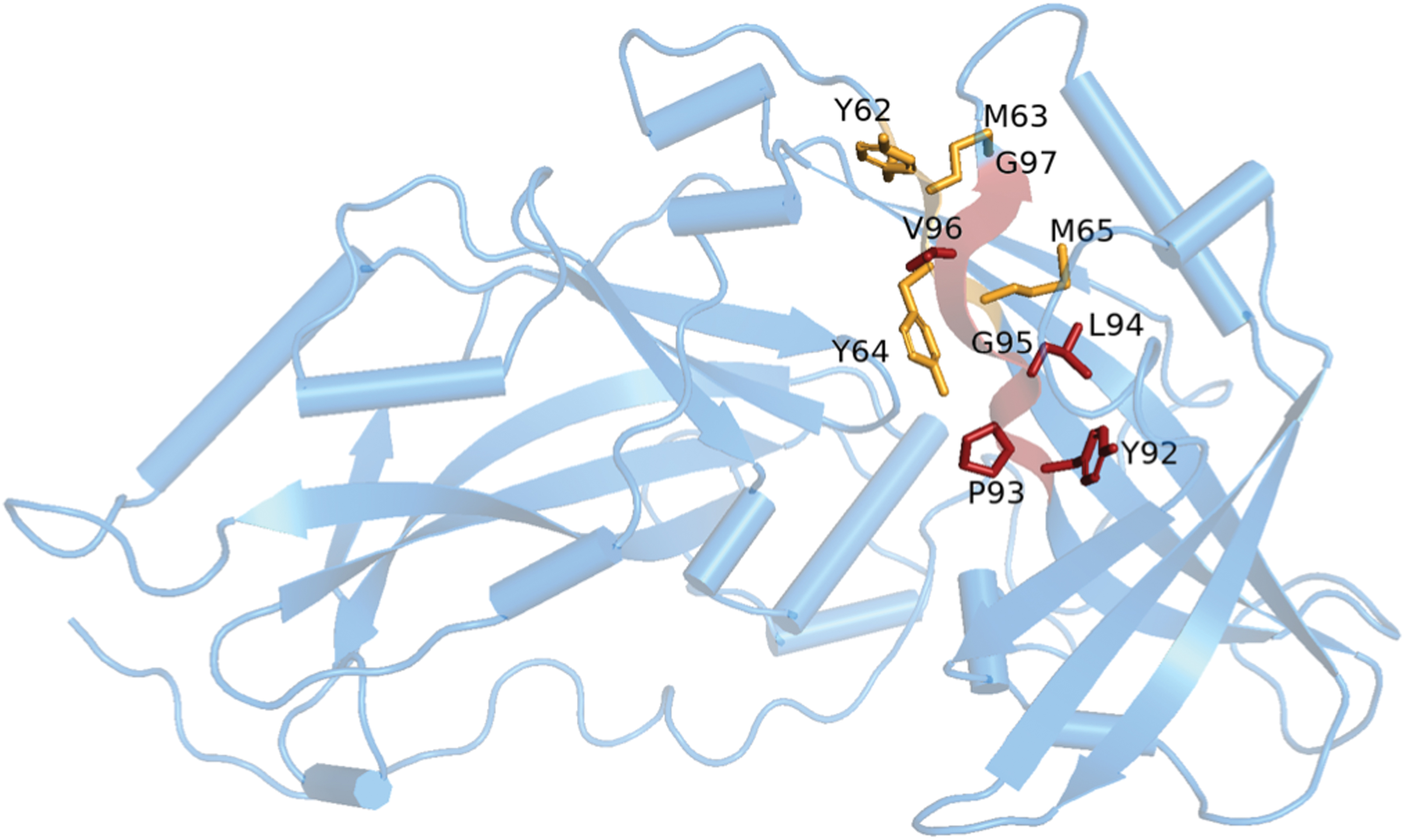
Late (L) domains encoded within viral proteins interact with host cell proteins to hijack the multivesicular body to drive viral budding. The amino acid sequence of NiV-M contains two motifs that have been described as L domains, 92YPLGVG97(48) and 62YMYL65 (11). The 90% sequence similarity of NiV-M and HeV-M allows confident prediction of the L-domains within the HeV-M structure, with the only variation being an L65M substitution. Deletion of the 92YPLGVG97 motif results in nuclear localization and abrogation of viral budding; however, replacement of the Ebola VP40 L-domain with the NiV-M YPLGVG sequence does not fully restore budding (48). Mutagenesis within the 62YMYL65 motif alters the cellular trafficking of M, with nuclear retention and abrogation of viral budding, despite preserved oligomerization (11). The YMYL sequence restores the budding ability of Ebola VP40 mutants; however, further investigation into host:L-domain interactions is required to provide further understanding of M trafficking. The L domains are contained within the N-terminal β-sandwich with the solvent-exposed residues being Y62, Y64, and V96 (Fig. 7). Due to the limited ability for these residues to participate in protein-protein interactions, improper functioning of M site or deletion mutants may be due to conformational changes or instability induced by disruption of the β-sheet. The HeV-M structure reveals hydrophobic interactions between the β-sheet core and the α2 helix, which contains a putative nuclear export signal (L106, L107, and L110) (51) and is important for oligomerization contacts (32). Changes to the M:host interactome and/or structural rearrangement due to these L domain mutations have yet to be experimentally determined.
Nuclear localization signals
Analysis of the amino acid sequence of HeV-M and NiV-M reveals two positively charged regions that could mediate nuclear import of the protein. These putative NLSs, one monopartite (mpNLS) and one bipartite (bpNLS), fit the consensus sequences described as classical NLSs (cNLS), suggesting that Henipavirus M binds to IMPA for recognition and translocation to the nucleus by the Importin α/β heterodimer. This is consistent with the experimental evidence described above. The putative NLS regions (42,51,54,69) are summarized in Table 3.
For bpNLS, the residues underlined are predicted to be interacting with the IMPA subunit, separated by a 10-residue linker.
IMPA, Importin-α; bpNLS, bipartite nuclear localization signal; mpNLS, monopartite nuclear localization signal; NiV, Nipah henipavirus.
NDV-M does not have a putative mpNLS within its sequence with the functioning bpNLS reported by numerous studies. Described by Coleman and Peeples (12), the M bpNLS motif of 247KKGKKVIFDKIEEKIRR263 (Fig. 8) is described as a nonclassical NLS that facilitates nuclear entry by interaction with IMP-β, without the adaptor subunit IMPA (17), similar to that of respiratory syncytial virus M protein (21). The bpNLS is located on α9 helix, which also forms a dimer-dimer contact within the M array (6). Despite similar tertiary structures of NDV-M and HeV-M, the proposed NLSs and oligomerization contacts differ, which may be linked to different functions of M proteins of henipaviruses.
Dimeric NDV M protein (PDB 4G1G) (6) indicating the bpNLS region located on the α9 helix. Color images are available online.
Several functional studies with mutagenesis have been undertaken to determine the functional NLS of henipavirus M. Both HeV-M and NiV-M localized to the nucleus in the early stages of infection before relocalization to the plasma membrane for budding. Mutations within the putative bpNLS changed the cellular localization of M (Table 4). Point mutation of the critical residue K258 (at the C-terminus of the predicted bpNLS) to a similarly charged arginine (R) resulted in increased diffuse nuclear localization and retention, with failure of M to locate to the plasma membrane for budding (42,69). In contrast, mutation of K258 to an uncharged alanine resulted in M being excluded from the nucleus (42,54,69) or nucleolus (51) with defective membrane association and budding. However, Rawlinson et al. (54) reported that HeV-M K258A mutant continued to localize to the nucleus, but not the nucleolus compartments, whereas no nuclear localization was apparent for NiV-M K258A. Interestingly, the K258A mutation also abrogates the interaction of M with Treacle (54), highlighting the multiple functions of this M domain. K258 clearly plays an integral role in the correct functioning of M within the host cell and structural analysis of these interactions must be pursued (8).
Mutagenesis Within Putative Nuclear Localization Signal Regions of Henipavirus Matrix Abrogate Cellular Localization and Virus-Like Particle Formation
VLPs here are defined as self-assembling and budding M protein shells surrounded by a host-derived bilayer membranous envelope.
No consistent, describable phenotype was observed; therefore, mutant was excluded from further investigation.
n/a, not investigated; GFP, green fluorescent protein.
In addition to the bpNLS, which is generally accepted to be the functional NLS for M protein, Wang et al. (69) produced a variety of mutants and tested the effect of uncharged alanine residues within the monopartite NLS, the bpNLS region 1, bpNLS region 2, and the entire bpNLS (Fig. 9). It was determined that R > A substitutions in region 1 of the bpNLS (244RR) lead to mild nuclear exclusion and moderately reduced budding. The impact of mutations within region 2 of the bpNLS (256RRK) was more severe with nuclear exclusion and defective budding, similar to K258A mutations. Interestingly, an mpNLS 81KRKK > AAAA mutation was reported to produce inconsistent localization patterns, with no discernible phenotype, and was excluded from further tests (69). The GFP-tagged mutant localizations in this study, however, showed reduced nuclear localization when compared to wild-type M. In light of these results, we examined the positioning of these NLS regions on the recently resolved crystal structure. The putative mpNLS is on a flexible loop, while the bpNLS with K258 is within an α-helix. Analysis of IMPA:NLS structures deposited to the protein data bank reveal that NLSs binding to IMPA do not form secondary structures such as α-helices when bound to IMPA (13,20,38,49,54,55,59,63) (Fig. 10). Moreover, it is interesting to note that the cNLS Mapper program, a prediction tool for scoring NLSs, scored the mpNLS from the prototypic henipaviruses as 7.5, while the putative bpNLS sequences were not detected as potential NLSs. Further experimentation may be required to resolve these ambiguities. Ascertaining the functional NLS of M protein has the potential to provide a novel target against Henipavirus infection and this approach has proven successful for other viral proteins; Ivermectin blocks import of HIV-1 integrase and dengue virus nonstructural protein NS5 (66,72), while 4-HPR blocks Zika virus NS5 (67). Prevention of the hijacking of nuclear import machinery by M could ultimately inhibit essential ubiquitination, innate immune antagonism, viral assembly, and budding, thereby potentially reducing the disease, social, and economic burden of these emergent viruses.
The HeV-M protein (PDB 6BK6) (32) in cartoon representation highlighting the putative cNLS regions (42,51,54,69).The bpNLS (lime) is positioned on an α-helix and not compatible with IMPA binding. The mpNLS (purple) was not resolved in crystallography due to its flexible nature and has been modeled for completeness. Color images are available online.
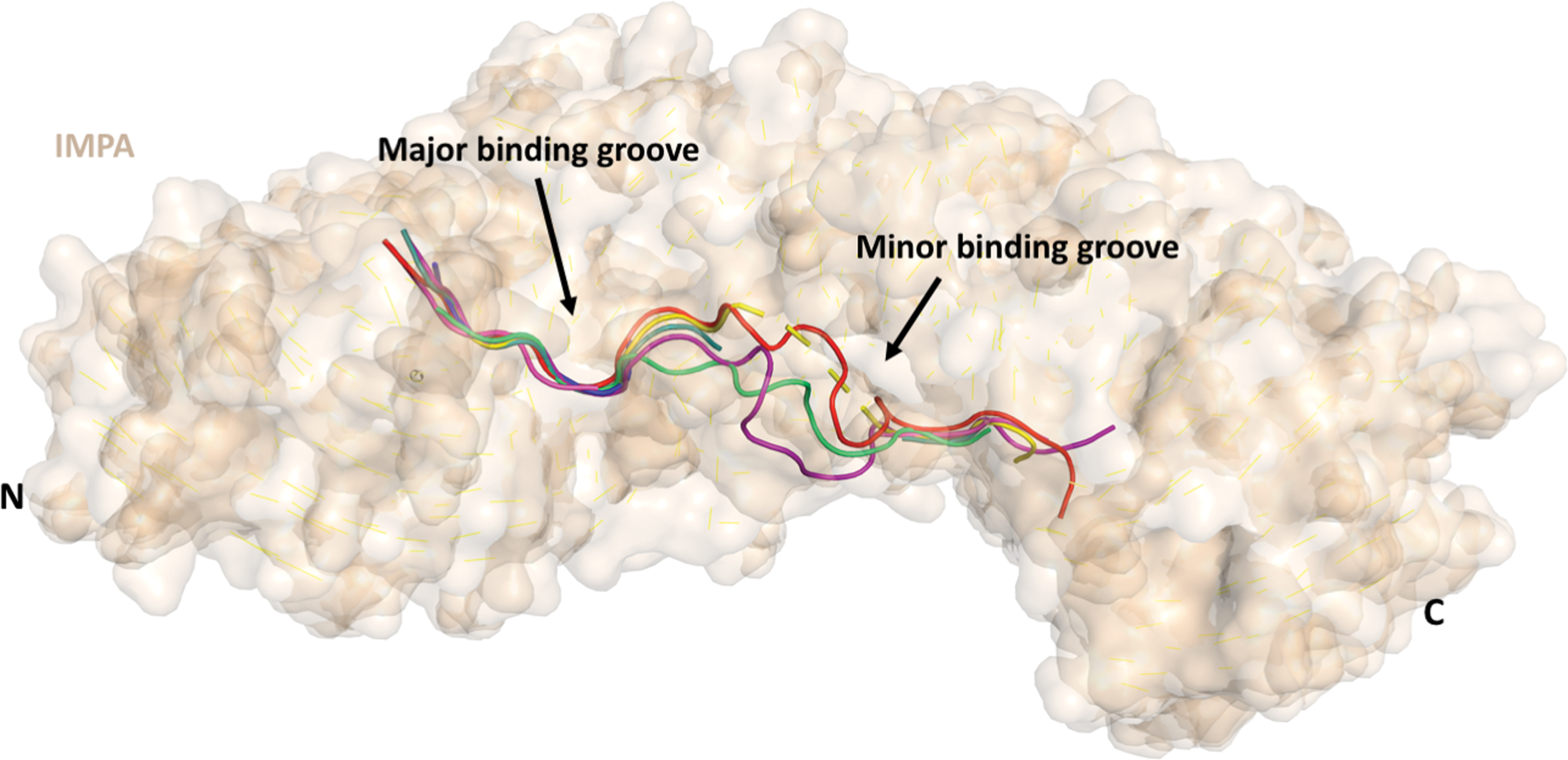
Analysis of IMPA:NLS structures deposited to the PDB reveal that NLSs binding to IMPA are unstructured flexible loops. PDB structures shown with IMPA in surface representation (wheaten) and NLSs in cartoon representation with 6BWB (red) (62), 4HTV (blue) (37), 3UKX (purple) (38), 5D5K (yellow) (55), 1EJY (green) (20), 4OIH (teal) (59), and 1BK6 (magenta) (13). Color images are available online.
Conclusion
The M proteins of henipaviruses undergo complicated, but necessary cellular trafficking and post-translational modifications to facilitate replication and the antagonism of host immune responses. The virus:host interaction interfaces that mediate these processes offer novel targets for the development of small molecule antivirals and/or attenuated vaccines. Understanding the mechanisms and biological interactions that allow Henipavirus M protein to direct viral assembly, while inhibiting innate immune responses, is critical for antiviral research efforts. Recent biochemical and structural endeavors have uncovered important regions within the protein that drive function, but further research is required to solidify some of the ambiguities between structure and function.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received.