
Abstract
Hantaan virus infection may cause severe lethal hemorrhagic fever with renal syndrome (HFRS) in humans. The chemokine fractalkine (CX3CL1) acts as a proinflammatory cytokine, and it is elevated in several infectious diseases. However, little is known about the contributions of CX3CL1 to HFRS pathogenesis. Present study detected plasma CX3CL1 levels and expression of the receptor CX3CR1 in HFRS patients and discussed the possible effects of CX3CL1 on pathogenesis of HFRS. Plasma CX3CL1 in acute phase and Critical/Severe groups of HFRS patients were significantly increased compared to that in normal controls (p < 0.001 and p < 0.01, respectively). High plasma CX3CL1 was negatively correlated with platelet count (r = −0.5844, p < 0.0001) and positively correlated with blood urea nitrogen (r = 0.3668, p = 0.0039), creatinine (r = 0.42, p = 0.0008), and white blood cells (r = 0.2646, p = 0.0411). Expression of CX3CR1 on nonclassical and intermediate monocytes was also increased in the acute phase (p < 0.01 for both the cells) and Critical/Severe groups (p < 0.05 and p < 0.01, respectively) of HFRS patients compared to that in normal controls. Taken together, elevation of plasma CX3CL1 in HFRS patients and expression of CX3CR1 on nonclassical and intermediate monocyte subsets might provide new insights into the potential role of CX3CL1/CX3CR1 in pathogenesis of HFRS.
Introduction
Hantaan virus (HTNV) causes hemorrhagic fever with renal syndrome (HFRS), which is a major nephropathis epidemica in Asia and Europe, and 90% of cases are documented in China (21). HTNV belongs to Hantavirus genus within family of Bunyaviridae, and it may cause a severe lethal HFRS in humans with a mortality rate of up to 10% (1). Typical course of HFRS involves five sequential phases, febrile, hypotensive, oliguric, diuretic, and convalescent phases, with clinical manifestations of sudden fever, hemorrhage, thrombocytopenia, and acute renal failure (20,40). It is widely accepted that HFRS pathogenesis largely entails an immune-mediated response involving immune complexes, complement activation, T cell responses, B cell responses, and cytokine production.
Previous reports suggested that a cytokine storm in early phases was a potential mechanism of HFRS pathogenesis (41). Proinflammatory cytokines, such as tumor necrosis factor-α (TNF-α), interferon-α (IFN-α), interleukin-1β (IL-1β), IL-6, IL-8, IL-33, IL-34, chemokine (C-C motif) ligand 2 (CCL2), CCL5, and chemokine (C-X-C motif) ligand 10 (CXCL10), are elevated after HTNV infections and correlate with severity of the disease (6,22,25,32,39,45,47). These cytokines may enhance the host defence against HTNV infection via recruitment and activation of innate and adaptive immune cells and induce an increase in endothelial permeability, which manifests as vascular endothelial barrier dysfunction (15,31). However, explanation of the immunological pathogenesis of HFRS is not sufficient.
Chemokines are a family of low-molecular-weight proteins, which are involved in homeostatic migration and pathogenesis of numerous inflammatory diseases and other disorders by mediating chemotaxis and adhesion of immune cells (4). Chemokine (C-X-C motif) ligand 3 (CX3CL1), also known as fractalkine, is the only member of CX3C class of chemokines with well-documented roles as a chemoattractant and in assisting CX3CR1+ cell adherence and migration (30,38).
CX3CL1 is primarily produced by endothelium in two forms. Membrane CX3CL1 is an adhesion molecule, and soluble CX3CL1 is a chemo attractant for CX3CR1+ cells (20). The membrane form of CX3CL1 is cleaved by metalloproteases ADAM10 and ADAM17 to generate the soluble forms of CX3CL1 (9), which appear in many infectious diseases and exhibit a proinflammatory function (7,18,27,35). CX3CR1 is widely expressed in immune and nonimmune cells in a highly cell-type-specific manner depending on the organ and tissue. For example, CX3CR1 in the brain is mostly expressed on microglia, and CX3CR1 expression in blood is restricted to monocytes, T cells, and some subsets of lymphocytes (2,11).
Human primary monocytes comprise a heterogeneous population that can be classified into three subsets based on the expression of CD14 and CD16: classical CD14++CD16− monocytes, intermediate CD14++CD16+ monocytes, and nonclassical CD14+CD16++ monocytes (33). In addition to the function of phagocytosis by classical monocytes, the intermediate monocytes are predominately involved in neovascularization, immune regulation, and tissue remodeling (12). Major roles of nonclassical monocytes, such as the patrolling behavior in lumen of blood vessels and retention as accessory cells of the inflamed endothelium to recruit other inflammatory cells, are largely dependent on the CX3CL1/CX3CR1 axis, and most monocytes interacting with the microvascular endothelium perform patrolling and surveillance functions in basal conditions (19,24,37).
Notably, CX3CL1 was the unique chemokine with its mRNA level upregulated in endothelial cells infected by pathogenic Hantaviruses (HTNV and New York-1 virus), but not by nonpathogenic Hantaviruses (Prospect Hill virus) (10). However, the roles of CX3CL1 and CX3CR1 in HFRS have not been fully elucidated. Present study detected the dynamic changes in plasma CX3CL1, analyzed its relationship with the parameters associated with the severity of the disease, and examined the expression of CX3CR1 on peripheral blood mononuclear cells (PBMCs) from HFRS patients to provide a better understanding of HFRS pathogenesis.
Materials and Methods
Patients and sample preparation
Seventy-four HFRS inpatients were recruited from 2014 to 2016 at Tangdu Hospital. Twenty-six healthy age, sex, and ethnicity-matched subjects were also recruited as controls. The clinical diagnosis of HFRS was serologically confirmed via the detection of specific IgM and IgG antibodies to HTNV in the patients' serum specimens. The plasma samples were isolated from EDTA (anticoagulant)-treated blood samples using centrifugation and cryopreserved at −80℃ before analysis.
The diagnostic criteria for HFRS disease severity in China are as follows: mild or moderate patients, from mild kidney damage with no obvious oliguric period to obvious symptoms of effusion (bulbar conjunctiva), uremia, hemorrhage (skin and mucous membrane), and significant oliguric period. The subjects were clinically diagnosed as Severe or critical patients when the effusion, uremia, hemorrhage, and kidney damage with oliguria (urine output, 50–500 mL/day) for ≤5 days or anuria (urine output, <50 mL/day) for ≤2 days; or visceral hemorrhage, refractory shock, heart failure, pulmonary edema, brain edema, severe secondary infection, and severe kidney damage with oliguria (urine output, 50–500 mL/day) for >5 days, anuria (urine output, <50 mL/day) for >2 days, or a blood urea nitrogen (BUN) level of >42.84 mmol/L were observed (20).
The course of HFRS was divided into two sequential phases according to clinical observations: acute phase (febrile, hypotensive, and oliguric) and convalescent phase (diuretic and convalescent). Patients with other kidney diseases, hematological diseases, diabetes, cardiovascular diseases, autoimmune diseases, viral hepatitis, or other liver diseases were excluded from this study. The Institutional Review Board of the Tangdu Hospital and the Fourth Military Medical University approved this study, and all of the enrolled patients and donors signed an informed consent form before blood sample collection.
Clinical information of the study subjects
A total of 116 plasma samples from 74 patients with HFRS were examined. Table 1 summarizes the information of all enrolled subjects.
Patient Characteristics of Enrolled Subjects in this Study
Enzyme-linked immunosorbent assay for the detection of CX3CL1 levels in plasma
Plasma CX3CL1 levels were measured using an enzyme-linked immunosorbent assay kit (R&D Systems, Minneapolis, MN) according to the manufacturer's protocol. Plasma samples (100 μL) were used for each test. Optical densities were determined at 450 nm with a 570-nm wavelength correction.
Flow cytometry
Blood samples (10 mL) were collected from each of the healthy controls and HFRS patients. PBMCs were isolated using density-gradient centrifugation. PBMCs (106 cells/test) were stained with fluorescent antibodies for flow cytometry analysis. Antibodies used in the study were specific for CX3CR1 (anti-CX3CR1-PerCP/Cy5.5, clone 2A9-1, BioLegend), CD3 (anti-CD3-FITC, clone HIT3a, BioLegend), CD19 (anti-CD19-APC, clone HIB19, BioLegend), CD56 (anti-CD56-PE, clone 5.1H11, BioLegend), CD14 (anti-CD14-FITC, clone 61D3, eBioscience), and CD16 (anti-CD16-PE, clone 3G8, BioLegend). Matched isotype controls were used as negative controls. Monocytes were gated based on their forward and side scatter characteristics. The data were analyzed using FlowJo 7.6.1 software (Oregon).
Statistical analysis
All analyses were performed using GraphPad Prism6 software. The significance of differences in CX3CL1 and CX3CR1 between different groups was determined using the Mann–Whitney U test. The nonparametric Spearman correlation test was used for correlation analyses of CX3CL1 levels with clinical parameters. p-Values (two-tailed) below 0.05 were considered statistically significant.
Results
Plasma levels of CX3CL1 in patients with HFRS
Plasma CX3CL1 levels during the acute phase in HFRS patients were significantly increased compared to those during the convalescent phase [2.298 (0.497–4.993) ng/mL vs. 0.893 (0.160–4.225) ng/mL, p < 0.001] and normal controls [0.649 (0.154–1.762) ng/mL, p < 0.001], but there was no significant difference in plasma CX3CL1 levels between HFRS patients in the convalescent phase and normal controls (p > 0.05, Fig. 1A).
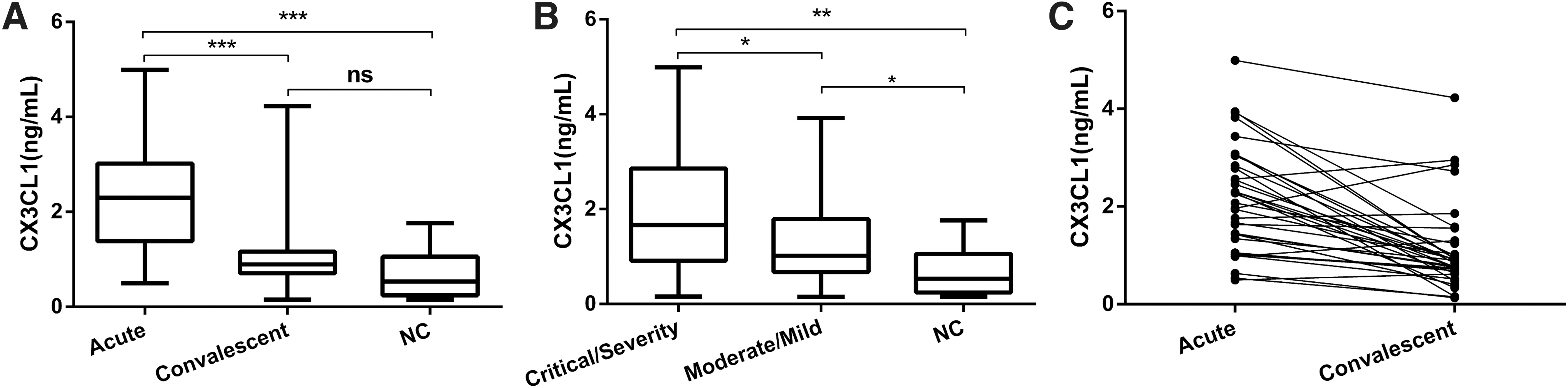
Dynamic changes of CX3CL1 levels in plasma of HFRS patients.
The plasma levels of CX3CL1 from HFRS patients in the Critical/Severe group were significantly higher than the Moderate/Mild group [1.669 (0.160–4.993) ng/mL vs. 1.01 (0.274–3.765) ng/mL, p < 0.05] and much higher than the normal controls [0.649 (0.154–1.762) ng/mL, p < 0.01, Fig. 1B]. The plasma CX3CL1 levels in the Moderate/Mild group were higher than the normal controls [1.01 (0.274–3.765) ng/mL vs. 0.649 (0.154–1.762) ng/mL, p < 0.05, Fig. 1B]. The acute and convalescent phase plasma CX3CL1 levels also showed the same tendency among 42 observed patients (Fig. 1C).
Plasma CX3CL1 levels correlated with the clinical parameters
To determine the relationship between plasma CX3CL1 levels and patients' clinical parameters, the clinical parameters and CX3CL1 levels were analyzed from samples drawn on the same day. Plasma CX3CL1 levels in HFRS patients negatively correlated with the platelet counts (PLT) (r = −0.5844, p < 0.0001, Fig. 2A), positively correlated with BUN (r = 0.3668, p = 0.0039, Fig. 2B), creatinine (Crea) (r = 0.42, p = 0.0008, Fig. 2C), and white blood cell (WBC) counts (r = 0.2646, p = 0.0411, Fig. 2D).
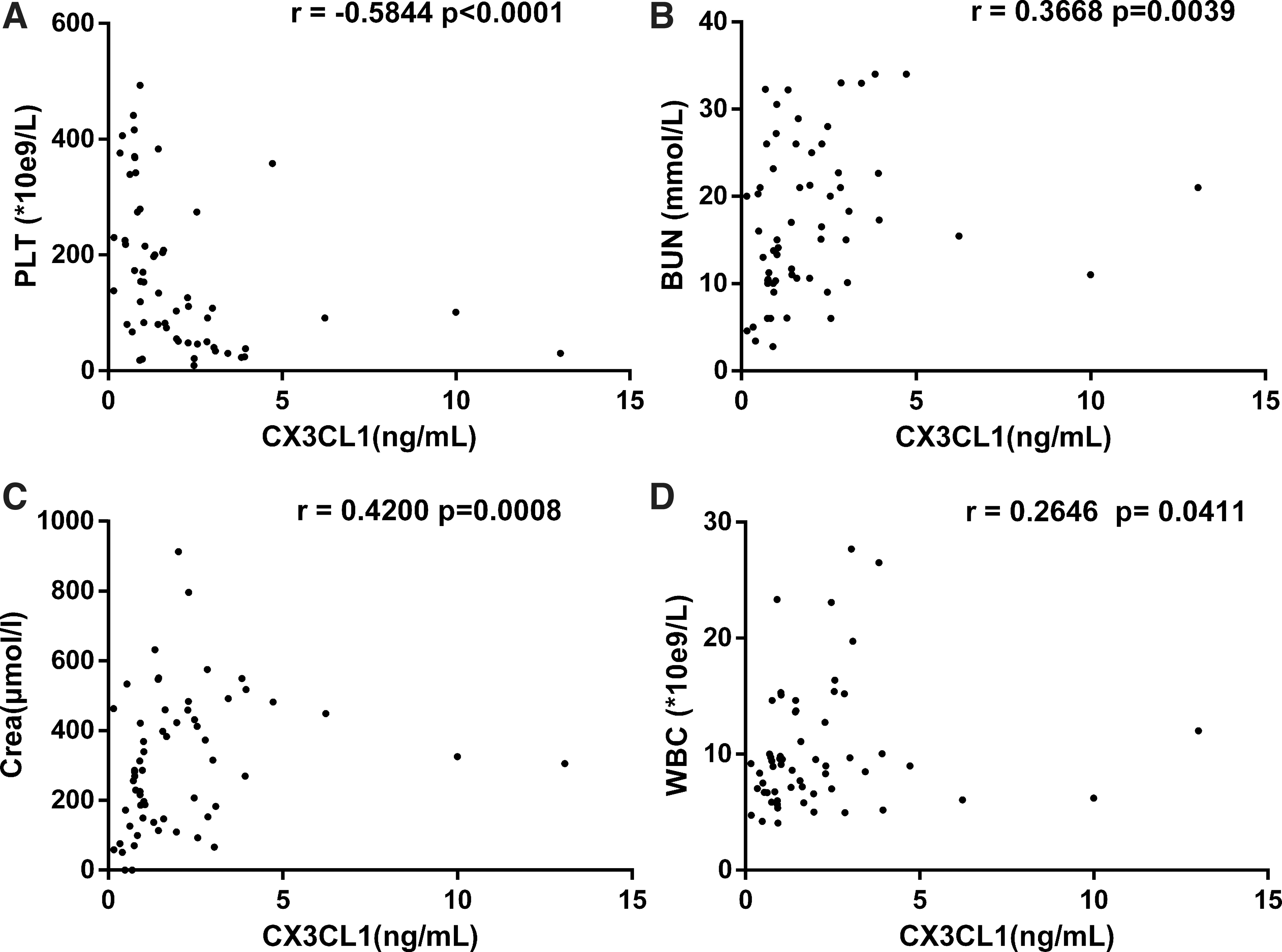
The correlations of plasma CX3CL1 levels with the observed clinical parameters of patients with HFRS. Negative correlations between the plasma CX3CL1 levels and platelet counts
Expression of CX3CR1 in monocytes from HFRS patients
To determine the distribution of receptor for CX3CL1, CX3CR1, the PBMCs from HFRS patients and healthy donors were analyzed by gating of CD3, CD19, CD56, and CD14-positive cells and CD3/CD56 double-positive cells. Results showed that CX3CR1 was expressed on CD14-positive monocytes, which was significantly increased during the acute phase in HFRS patients compared to that in the normal controls (p < 0.01) and was decreased during the convalescent phase (p < 0.05), but it remained higher than the level in normal controls (p < 0.05, Fig. 3). Although CX3CR1 was also expressed on CD3-positive T cells, CD19-positive B cells, and CD56-positive NK cells, rarely on CD3 and CD56 double-positive NKT cells, there were no significant differences in the expression of CX3CR1 on these cells between HFRS patients and normal controls (p > 0.05 for all comparisons).
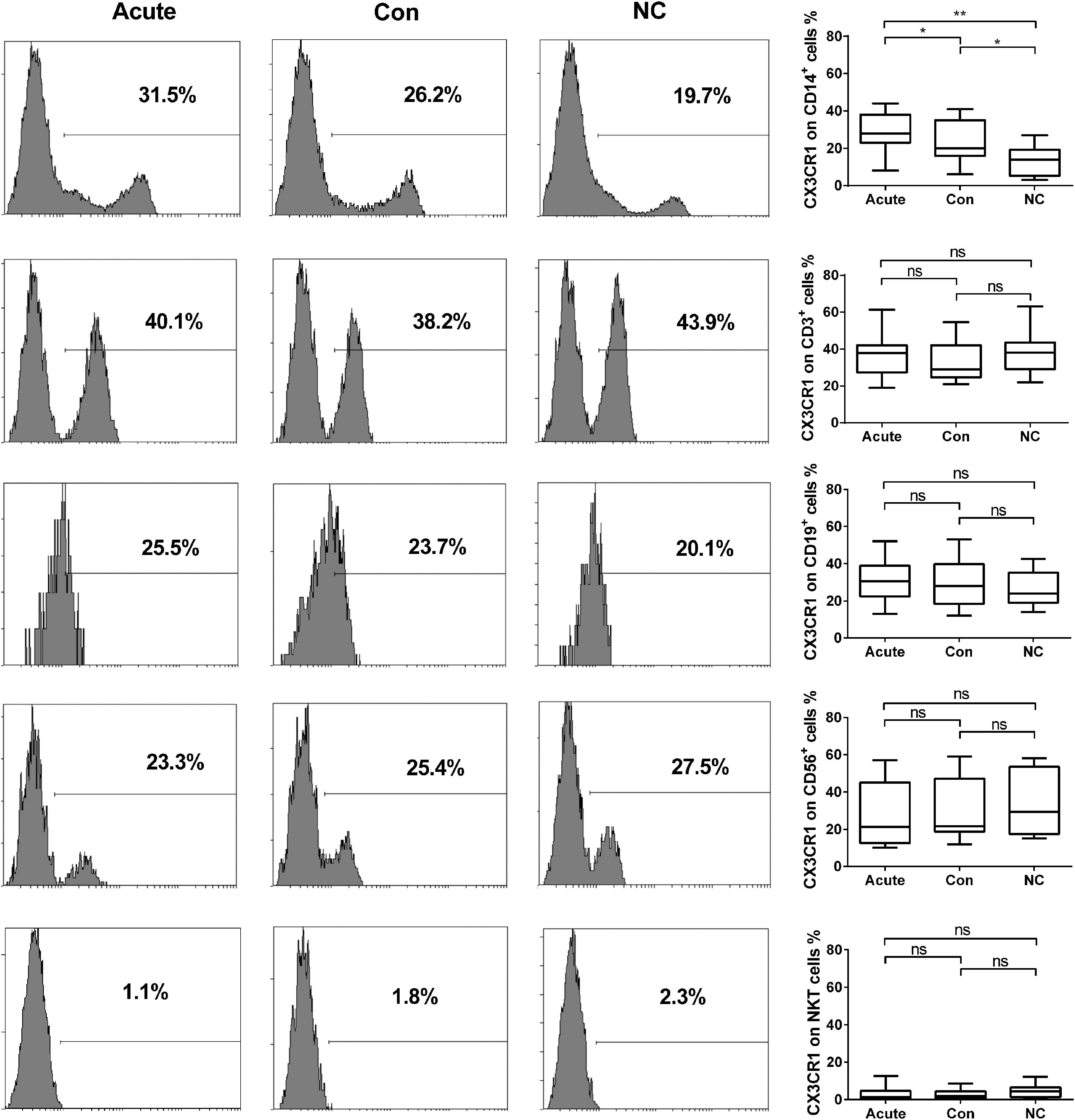
CX3CR1 expression on PBMC from HFRS patients of different phases. Flow cytometry figures on left side are from each representative case of CX3CR1 on PBMC in acute and convalescent phase of HFRS patients and normal controls. Comparison of CX3CR1 expression on PBMC in different phase of HFRS patients and normal control (right panel). The significance of the differences between different groups was determined with the Mann–Whitney U test, Black lines represent medians. p-Values below 0.05 were considered statistically significant. *p < 0.05; **p < 0.01; ns = no significant difference. PBMC, peripheral blood mononuclear cell.
Further studies showed that CX3CR1 levels on monocytes were also significantly increased in the Critical/Severe group compared to the normal controls (p < 0.01) and decreased in the moderate/mild group (p < 0.05), but it remained much higher than the control groups (p < 0.05, Fig. 4).
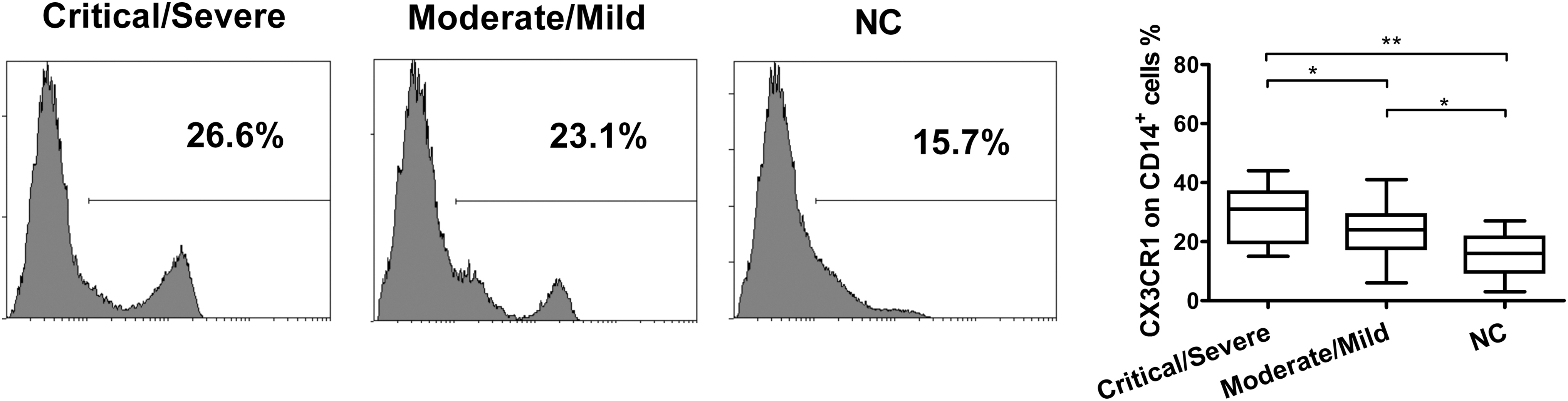
CX3CR1 expression on monocytes from HFRS patients of different severity. Representative expressions of CX3CR1 on monocytes from Critical/Severe and Moderate/Mild groups of HFRS patients and normal controls are shown (left panel). Comparison of CX3CR1 expression on monocytes in different severity groups of HFRS patients and normal control (right panel). The significance of the differences between different groups was determined with the Mann–Whitney U test. Black lines represent medians. p-Values below 0.05 were considered statistically significant. *p < 0.05; **p < 0.01.
To investigate the distribution of CX3CR1 in different monocyte subsets, the peripheral blood classical (CD14++CD16−), intermediate (CD14++CD16+), and nonclassical (CD14+CD16++) monocytes in HFRS patients and controls were characterized using flow cytometry. As shown in Figure 5, the expression of CX3CR1 on nonclassical and intermediate monocytes during the acute phase in HFRS patients was significantly increased compared to the healthy controls (p < 0.05 for both cells), while there were no differences in the expression of CX3CR1 on classical monocytes in different phases of HFRS patients compared with the normal controls (p > 0.05 for all comparisons).
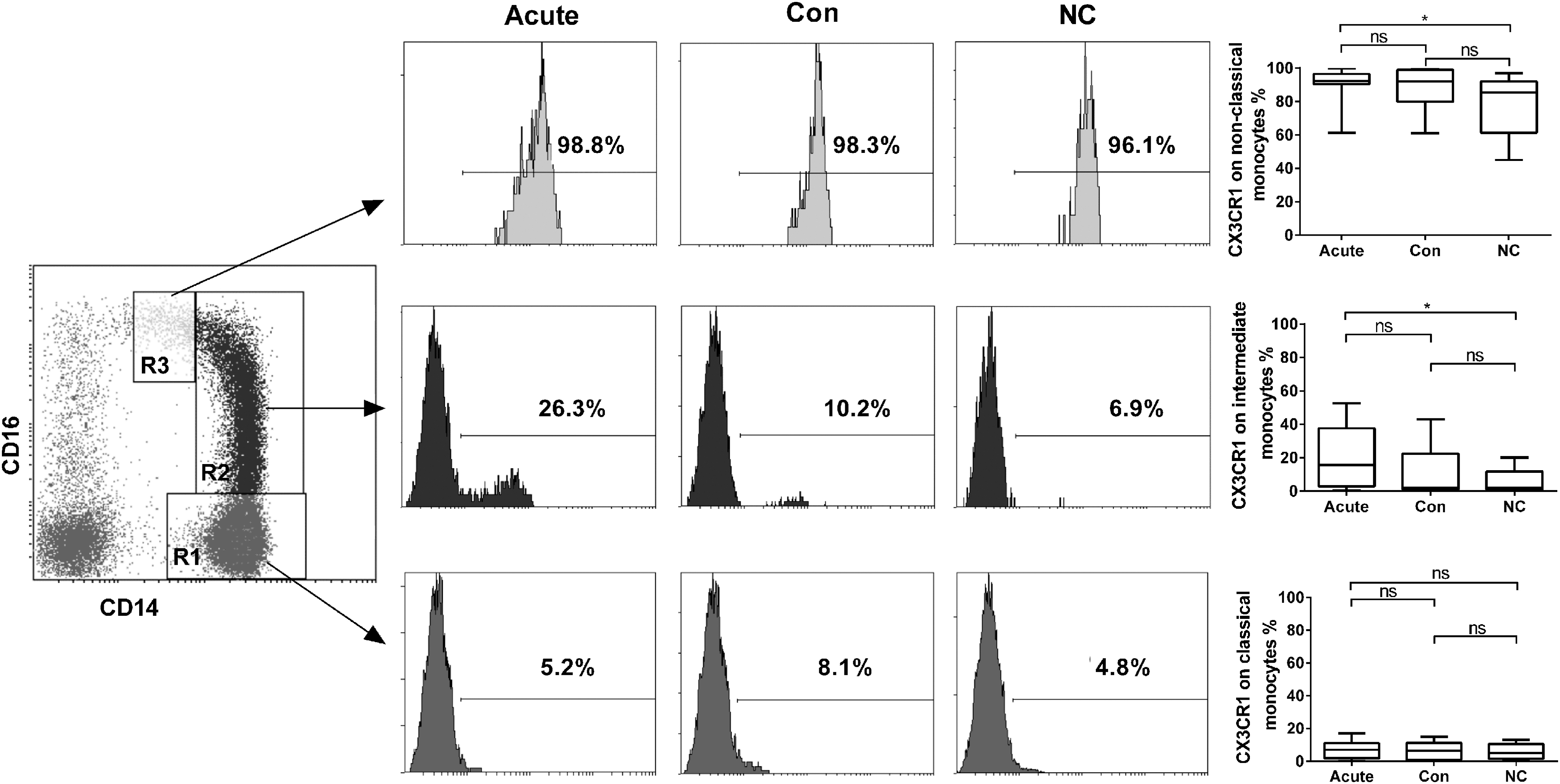
CX3CR1 expression on different monocyte subsets from HFRS patients of different phases. Monocytes, gated by their CD14 and CD16 staining levels, were divided into classical (CD14++CD16−) (R1), intermediate (CD14++CD16+) (R2), and nonclassical (CD14+CD16++) (R3) monocytes (left panel), and representative distributions of the monocyte subsets during the acute and convalescent phases of HFRS patients and in normal control are shown in the graph (middle panel). Comparison of CX3CR1 expression on different monocyte subsets in different phases of HFRS patients and normal control (right panel). The significance of the differences between different groups was determined by the Mann–Whitney U test. Black lines represent medians and p-values are plotted in each graph. *p < 0.05.
Furthermore, the expression of CX3CR1 on intermediate monocytes was greatly increased in the Critical/Severe group of HFRS patients compared to the Moderate/Mild group and normal controls (p < 0.05 and p < 0.01 respectively), the expression of CX3CR1 on nonclassical monocytes was increased only in the Critical/Severe group of HFRS patients compared to the normal controls (p < 0.05) and there were no differences of that on classical monocytes from HFRS patients of different severity (p > 0.05 for all comparisons, Fig. 6).
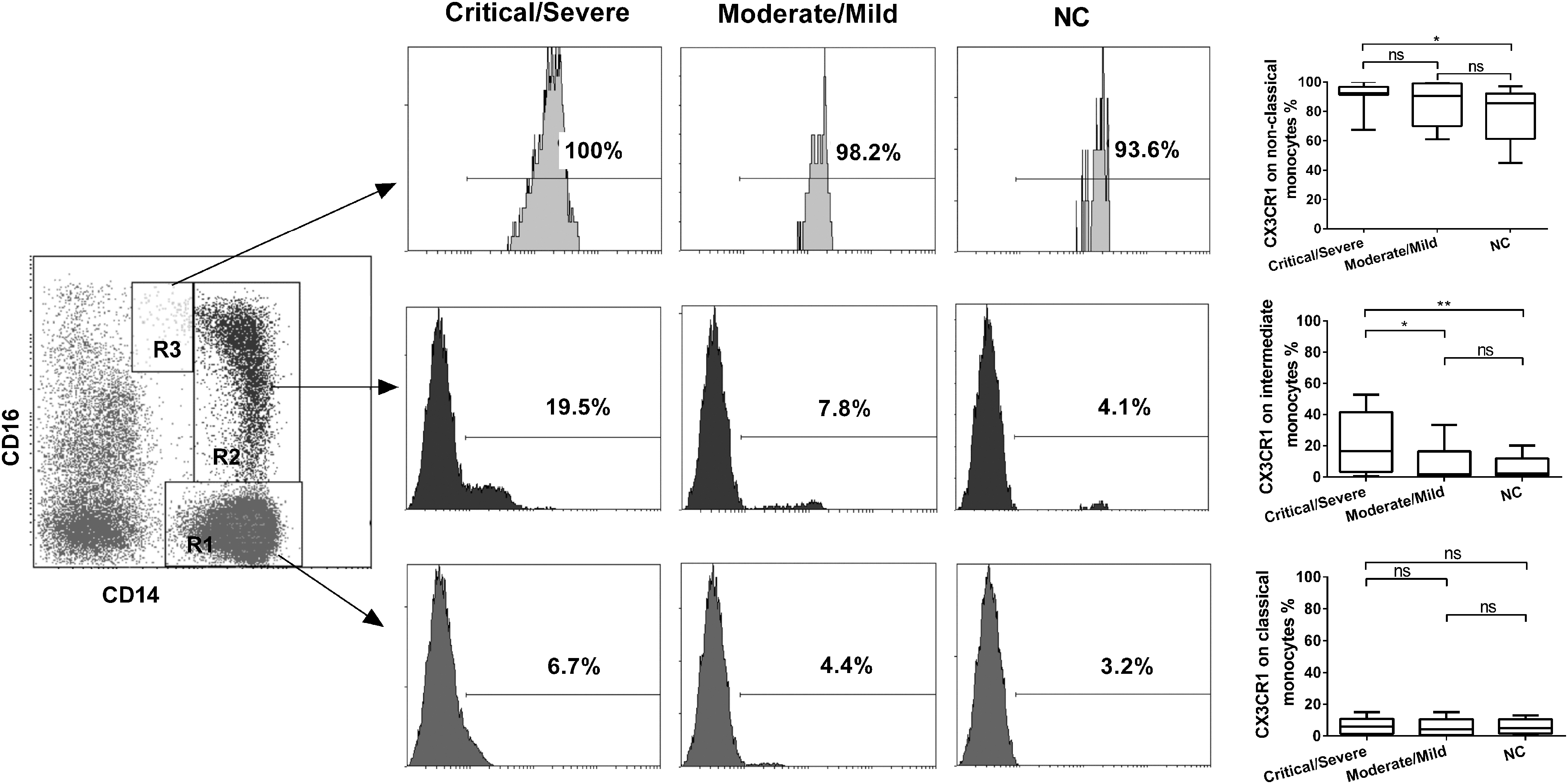
CX3CR1 expression on different monocyte subsets from HFRS patients of different severity. Representative results of CX3CR1 expression on different monocyte subsets of PBMC in Critical/Severe and Moderate/Mild groups of HFRS patients and normal controls (left and middle panel). Comparison of CX3CR1 expression on different monocyte subsets in different severity groups of HFRS patients and normal control (right panel). The significance of the differences between different groups was determined by the Mann–Whitney U test. Black lines represent medians and p-values are plotted in each graph. *p < 0.05; **p < 0.01.
Discussion
HFRS is characterized by increased vascular permeability and hemorrhages. However, HTNV did not cause any noticeable cytopathic effects in human umbilical vein endothelial cells (HUVECs), which are highly susceptible to HTNV infection, in vitro. Notably, HUVECs produced many inflammatory cytokines after HTNV infection, which suggests that the response of endothelial cells to HTNV is central to the development of HFRS diseases (36,44). Various cytokines and chemokines, such as CCL2, CCL5, and CXCL10, in serum are significantly increased during the acute phase in HFRS patients (13,23,36). The elevation of plasma CX3CL1 is associated with various inflammatory diseases, such as sepsis and virus infection (8,17,28). A comparison of mRNAs from HTNV-infected endothelial cells and mock-infected endothelial cell controls showed that the mRNA level of CX3CL1 was upregulated 9.5-fold 4 days after HTNV-infection (10).
Our findings of elevated plasma CX3CL1 in the acute phase and critical and severe groups of HFRS patients support its role as a proinflammatory cytokine, which is consistent with these previous studies. We also observed that plasma CX3CL1 had a positive relationship with WBC counts, BUN, and creatinine and negatively correlated with platelet counts in HFRS patients, which indicates that plasma CX3CL1 correlated with the severity of HFRS and thrombocytopenia during HTNV infection. However, recent studies showed that CX3CL1 also reduced pathopoiesis in some renal diseases, which suggests that CX3CL1 levels are a double-edged sword. These results provide a novel perspective for the pathogenesis and treatment of renal diseases and disorders (49). The functional role of CX3CL1 in HFRS should be characterized in future studies.
Biological effects of CX3CL1 are the result of its interaction with CX3CR1. CX3CL1 and CX3CR1 are widely expressed in immune and nonimmune cells throughout organisms and exhibit cell-type-specific expression in different tissue. CX3CR1 is expressed on monocytes, macrophages, lymphocytes, mast cells, and dendritic cells in blood (3). Thus, interaction between CX3CL1 and CX3CR1 is one initial step in host defense for monocyte crawling or “patrolling” in the lumen of blood vessels, which indicate the unique immune regulation of CX3CL1 on monocyte (28).
CX3CR1 expressing immune cells play main roles in pro- and anti-inflammatory responses, depending on environmental condition. CX3CR1 expression in PBMCs promotes inflammation in granulomatosis with polyangitis (GPA) by promoting the migration of leukocytes into inflammatory lesions (5). Th1 cells expressing CX3CR1 are increased, which increases Th1-related cytokines IFN-γ and IL-12 and promotes inflammation.
As previously reported, CX3CR1 is predominantly expressed on monocyte in the peripheral blood and functions as an important component of innate and adaptive immunity (26). A deficiency in CX3CR1 signals reduces the binding of monocytes with the injured endothelium and results in decreased adherence (30). Other studies showed that CX3CR1 signals in monocyte enhance the expression of antiapoptotic factor B cell lymphoma 2 (Bcl2), which is required for monocyte homeostasis and arteriosclerosis, by promoting cell survival (14).
CX3CR1 was primarily expressed on CD14+ monocytes in HFRS patients in the present study, and the percentage of CX3CR1-positive monocytes showed a significant increase in the acute phase compared to the convalescent phase and normal controls, which suggests that the CX3CL1-CX3CR1 axis participates in the quick recruitment of monocytes in the early phase of HTNV infection. CX3CR1+ monocytes were also increased in the Critical/Severe group compared to the Moderate/Mild group and normal controls, which suggests that CX3CR1-positive monocytes correlate with the severity of HFRS.
We also found that CX3CR1 was expressed on CD3-positive T cells, CD19-positive B cells, and CD56-positive NK cells with no significant difference between the HFRS patients and the normal controls, which suggests that the elevated plasma CX3CL1 cooperates with CX3CR1 on monocytes in the recruitment of these phagocytes after HTNV infection.
Three major subpopulations of circulating monocytes are distinguished according to the CD14 and CD16 ratio: CD14++ CD16– (classical), CD14++ CD16+ (intermediate), and CD14+ CD16++ (nonclassical) (50). The proportion of different subsets changes during infections, such as sepsis and hepatitis B, which indicate the clinical significance of the monocyte subsets during inflammation (42,43).
The dynamic changes in monocyte subsets with different biomarkers were also associated with HFRS. For example, soluble CD14 in plasma was significantly higher in the acute phase in HFRS patients and these levels correlated with monocyte expansion and activation, which were characterized by increased blood monocyte counts, the proportion of CD14++ CD16+ intermediate monocytes, and elevated plasma TNF-α and soluble CD163 (sCD163) levels (40,46). Another study showed that HTNV infection induced significantly higher numbers of circulating CD14++ CD16+ CD163+ monocytes and CD14++ CD16+ CD206+ monocytes, which suggest the involvement of CD14++ CD16+ cells in HFRS pathogenesis (29).
CX3CR1 may be used for the identification of nonclassical monocytes with specific functional properties, for example, patrolling of the vascular endothelium under homeostatic and inflammatory conditions for the removal of damaged cells and debris from the vasculature, regulation of wound healing, and the resolution of inflammation in damaged tissues (44). CX3CR1 was predominantly expressed on the nonclassical monocytes in HFRS patients and normal controls in the present study. The expression of CX3CR1 on classical monocytes showed no difference in the different phases or severity groups of HFRS compared with the normal controls, but CX3CR1 on nonclassical and intermediate monocytes showed a significant increase in the acute phase and Critical/Severe groups of the disease. The expression of CX3CR1 on intermediate monocytes in the Critical/Severe group of HFRS patients was much higher than the Moderate/Mild group.
In our previous study, we found that the elevated CXCL10 Level in HFRS patients' sera was positively correlated with HFRS severity, and CXCR3 increased on all monocyte subsets which suggested that the unregulated CXCL10 in the acute phase of HFRS may recruit all monocyte subsets and the recruited monocyte may produce excessive cytokines to be involved in the “cytokine storm” (45). In the present study, we found that plasma level of CX3CL1 was also increased, but only intermediate and nonclassical monocyte subsets showed the upregulation of CX3CR1. These results suggested that the monocyte subsets recruited by CX3CL1 may enroll in immune regulation and the patrolling behavior in the lumen of blood vessels and retention as accessory cells of the inflamed endothelium to recruit other inflammatory cells, which indicated the uniqueness function of CX3CL1 and CX3CR1 on monocyte in HFRS.
Moreover, the mechanisms for upregulation of CX3CR1 on immune cells depends on CX3CL1 itself, TGF-β and some kinds of virus fragments (16,34,48). Since CX3CR1 expressed not only on monocytes but also on T cells, B cells, and NK cells in HFRS, there should be other factors except CX3CL1 itself induced the different expression of CX3CR1 on monocyte subpopulations, which need to be determined in the further studies. Because vascular endothelial damage is a hallmark of HFRS, the expression of CX3CR1 on nonclassical and intermediate monocytes in HFRS indicates the fast recruitment of these “patrolling” cells and “tissue-remodelling” cells in the vascular due to the CX3CL1 that is produced by HTNV-infected endothelial cells.
Taken together, our findings demonstrated that HFRS patients showed a marked increase in plasma CX3CL1 and the frequency of CX3CR1 on nonclassical and intermediate monocyte subsets. These changes seemed to depend on the state and severity of the disease. These results suggest that different subsets of monocytes recruit under infectious status as a result of the CX3CL1 and CX3CR1 axis and participate in the immunological pathogenesis of HFRS. However, the underlying mechanisms of the observed changes in monocyte subsets in HFRS patients should be explored further.
Conclusions
Elevated plasma CX3CL1 levels during the acute phase in HFRS were associated with disease severity. CX3CL1 may recruit CX3CR1-positive cells, especially intermediate and nonclassical monocyte subsets to participate in inflammation during the early phase of HTNV infection. These results provide new insights into the understanding of immunopathology of HFRS.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the National Natural Science Foundation of China (grant no: 81771705; 81501362; 81701555; 81871239; 81901600).