
Abstract
Objective:
Reconstituted high-density lipoproteins (rHDL) improve wound healing in diabetes. We aimed to determine if rHDL elicit anti-inflammatory effects in diabetic wounds, as a mechanism to explain their wound healing benefits.
Approach:
Diabetes was induced using streptozotocin in C57Bl6/J mice. Two full-thickness wounds were placed on the subflanks of diabetic and nondiabetic (ND) mice. Phosphate-buffered saline (PBS) or rHDL (50 µg/wound/day) were applied topically. Wound closure was assessed daily. Inflammatory gene transcripts were measured by qPCR and proteins by Western blotting and enzyme-linked immunosorbent assay in wounds collected at baseline, 24 h, and 3 days postwounding. Wound macrophages were assessed by flow cytometry 7 days postwounding. The fate of fluorescent 3,3-dioctadecyloxacarbocyanine, perchlorate (DiO)-labeled rHDL was tracked by flow cytometry, fluorescent imaging, and microscopy.
Results:
In diabetic mice, rHDL increased wound closure rates at days 6 (+288%, p < 0.01) and 7 (+639%, p < 0.0001) postwounding, compared with PBS controls. After 3 days, rHDL-treated diabetic wounds had lower Rela (−65%) and C–C motif chemokine ligand 2 (Ccl2) (−59%) mRNA levels and CCL2 protein (29%) than PBS controls, p < 0.05 for all. Wound macrophage content was higher in diabetic than ND wounds, but rHDL did not change macrophage content or polarity. DiO-rHDL were taken up by key wound cells including fibroblasts, macrophages, keratinocytes and endothelial cells, and retained in wounds for at least 48 h.
Innovation:
rHDL exerts anti-inflammatory effects in diabetic wounds early postwounding, which may contribute to its wound healing properties.
Conclusion:
The anti-inflammatory properties of rHDL in diabetic wounds present topical rHDL as a novel treatment option for improving healing in patients with diabetic foot ulcers.
Christina A. Bursill
This is a visual representation of the abstract.
INTRODUCTION
Approximately 25% of people living with diabetes develop diabetic foot ulcers (DFU) during their lifetime and of these, 20% require amputation.1,2 Despite the significant clinical demand for an effective DFU treatment, there are none that actively improve healing. This is likely due to the complexities of the wound healing process that consists of four continuous, yet overlapping, phases including hemostasis, inflammation, proliferation, and tissue maturation and vascular remodeling. 3 An agent with pleiotropic effects may therefore promote wound repair in diabetes more effectively.
We have previously shown in a murine wound healing model that topical reconstituted high-density lipoproteins (rHDL) rescue diabetes-impaired wound healing. 4 It was demonstrated that rHDL improved wound angiogenesis, critical to successful wound healing. 4 Whether these beneficial effects of rHDL on diabetic wound healing extend beyond its pro-angiogenic capabilities remains to be fully elucidated.
Inflammation plays an important role in the early stages of wound healing by preventing infection, however, prolonged, inappropriate inflammation has negative effects on the healing process, 5 which is exacerbated in diabetic patients. 6 Diabetes significantly prolongs wound inflammation and halts the normal sequence of the wound repair process; this includes disruption to wound macrophage regulation. The increased infiltration of inflammatory cells including neutrophils and macrophages into the wound area results in significantly higher levels of pro-inflammatory chemokines, such as C–C motif chemokine ligand 2 (CCL2), prolonging the inflammatory phase and delaying the healing process. 7 Nuclear factor κB (NF-κB), is the master regulator of a host of inflammatory mediators, including CCL2. Research in diabetic mouse models demonstrated that hyperglycemia causes the release of a large number of inflammatory cytokines by upregulating the NF-κB pathway, which delays diabetic wound healing.8,9
A major disruption in diabetic wound healing is a delay or failure of the phenotypic switch from pro-inflammatory macrophages to anti-inflammatory macrophages. Shortly after injury and hemostasis, monocyte-derived macrophages enter the wound site and differentiate into pro-inflammatory M1-like macrophages which release inflammatory cytokines including interleukin-6 (IL-6).10,11 This is followed in the subsequent healing stages by polarization into anti-inflammatory M2-like reparative macrophages, which release anti-inflammatory cytokines such as transforming growth factor-β1 (TGF-β1) and support tissue remodeling and fibrosis.10,11 In diabetes, the transition from the M1 to the M2 phenotype is delayed, resulting in persistent wound inflammation that promotes chronic nonhealing wound development.5,11 Macrophages isolated from chronic wounds of patients with type 2 diabetes mellitus exhibit a pro-inflammatory M1-like phenotype. 12 Furthermore, wound macrophages obtained from diabetic mice persisted as pro-inflammatory M1-like macrophages and failed to transition to the reparative M2-like phenotype. 12
HDL exhibits potent anti-inflammatory properties.13,14 In vitro, incubation with rHDL inhibits cytokine-stimulated expression of CCL2 in endothelial cells, 13 smooth muscle cells, 15 and monocytes/macrophages.13,16,17 HDL also increases the transition of M1-like macrophages to M2-like macrophages. 18 Despite well-established anti-inflammatory properties, the effect of rHDL on diabetic wound inflammation is currently unknown. Furthermore, while rHDL have been applied topically to full-thickness murine wounds in several studies,4,19 its fate and association with wound cells has never been tracked. This information is important for understanding the required frequency of application and to support its future translation.
Accordingly, this study sought to assess the anti-inflammatory effect of rHDL in a diabetic murine model of wounding4,20 and track the fate of a fluorescently labeled rHDL. Our findings strengthen support for rHDL as a novel topical agent that actively improves diabetic wound healing.
CLINICAL PROBLEM ADDRESSED
With the escalating prevalence of diabetes, the number of patients with DFUs is also rising. This is coupled with ever-increasing social and economic burdens, due to the morbidity that DFUs cause and the multifaceted, labor-intensive approach required for DFU care. The topical application of rHDL presents as a novel wound healing agent with pleiotropic benefits for promoting diabetic wound healing including the promotion of wound angiogenesis and the potential to elicit anti-inflammatory effects. As an endogenous agent, rHDL have a very low risk of causing deleterious outcomes and is able to be readily scaled up for future clinical translation. rHDL have the potential to add benefit on top of, or work in parallel, with current treatment and management strategies that include biological agents, new skin and dressing technologies, and offloading regimens to help relieve the burden of DFUs.
MATERIALS AND METHODS
Electronic laboratory notebook was not used.
Discoidal rHDL
Apolipoprotein A-I (apoA-I), the main protein component of native HDL, was isolated from pooled donated plasma samples from healthy humans obtained from the Australian Red Cross (Supply Agreement 14-02NSW-04) by ultracentrifugation and anion-exchange chromatography, as described previously.4,13,15,17,19 Discoidal rHDL were prepared by complexing purified lipid-free apoA-I with the phospholipid 1-palmitoyl-2-linoleoyl-phosphatidycholine (PLPC) at an initial PLPC: apoA-I ratio of 100:1. The rHDL were filter sterilized prior to use in all cell culture experiments and the final apoA-I concentration was determined using the bicinchoninic acid (BCA) assay. The final PLPC: apoA-I molar ratio ∼80:1.
Animal studies
All experimental protocols were approved by the SAHMRI South Australian Health and Medical Research Institute animal ethics committee (SAM#301) and conformed to the Australian code for the care and use of animals for scientific purposes. Six to eight-week-old C57Bl/6J mice were rendered diabetic by a single high dose (165 mg/g) of streptozotocin (Sigma-Aldrich, St. Louis, MO, USA) injected intraperitoneally. This high dose STZ model is required to drive sufficient hyperglycemic conditions that will trigger diabetes-impaired wound healing, as described previously. 4 Hyperglycemia was defined as a blood glucose level of ≥15.0 mmol/L.
Murine model of wound healing
A murine model of wound healing was conducted as previously described at 2.5 weeks post-STZ administration.4,19–21 In brief, following isoflurane anesthesia (5% to 3–4% for maintenance), two full-thickness wounds (4 mm in diameter) were placed on the back flanks of mice and splinted with silicone splints (10 mm diameter circle with 6 mm inner circle cut). The placement of silicone splints around the wound area prevents healing by wound contraction that commonly occurs in rodents and drives wound healing by epithelialization, cellular proliferation and angiogenesis, which closely mirror the biological processes seen in humans. 20 For each mouse, one wound received rHDL (50 µg/wound/day) and the other received phosphate-buffered saline (PBS, vehicle control) by topical application. rHDL (25 μL) were applied at the time of surgery and then daily (every 24 h) thereafter until the study conclusion. A transparent occlusive dressing (Opsite) was applied. Wound tissues were excised at the time of wound surgery for baseline measurements (0 h). Wound diameter measurements along the x-, y-, and z-axes areas were measured daily with micro-callipers. A calculation for wound area was performed from the average of three daily diameter measurements along the x-, y-, and z-axes. Wound closure is expressed as a percentage of initial wound area at day 0. Two cohorts of mice were used to track the changes in wound inflammation at 24 h (early inflammation) and day 3 (mid inflammation) postwounding. An additional cohort of mice was used at day 7 postwounding for flow cytometry analysis of wound macrophages.
Gene expression
Total RNA from murine wound tissue was isolated using TRIzol reagent (Sigma-Aldrich) as per the manufacturer’s instructions. RNA concentration was quantified using NanoDrop 800 spectrometer (Thermo Fisher Scientific, Waltham, MA, USA). Total RNA (400 ng) was reverse transcribed using the iScript RT Supermix (Bio-Rad, Hercules, CA, USA). Quantitative real-time PCR was performed to detect expression of Rela (p65 subunit of NF-κB), Nfkb1 (p50 subunit of NF-κB), Ccl2, Ccl5, Vcam1, Icam1, Tgfb1, and Il6 with housekeeping gene 36B4 using previously published primers.13,21,22 Relative changes in gene expression were calculated using the ΔΔCt method, normalized to murine 36B4.
Protein expression
Wound tissues were snap frozen on dry ice after dissection and stored at −80°C. Tissues were lysed in radioimmunoprecipitation assay buffer on ice for 20 min, homogenized and clarified by centrifugation for 15 min at 4°C. Supernatants were collected and protein concentration was determined using Pierce BCA protein assay kit (Thermo Fisher Scientific). Overall, 15 μg of wound protein lysates were subjected to Western blot analysis and probed for IκBα (#4814, Cell Signaling Technology, Danvers, MA, USA), phosphorylated (S536) p65 NF-κB (ab86299, Abcam, Cambridge, United Kingdom) and total p65 NF-κB (ab32536, Abcam). Even loading was confirmed by α-tubulin (ab7291, Abcam). Protein lysates were also used in commercially available enzyme-linked immunosorbent assays for CCL2, TGF-β1, and IL-6 (R&D Systems, Minneapolis, MN, USA) as per the manufacturer’s instructions.
Flow cytometry for identifying macrophage populations
The mice were euthanized 7 days postwounding and the wound tissues were excised and digested with liberaseTM (50 µg/mL, 2 h, 37°C) (Roche Applied Science, Penzberg, Germany). Disaggregates were neutralized with Iscove’s Modified Dulbecco’s Medium (Sigma-Aldrich) supplemented with 10% fetal bovine serum (FBS; CellSera Australia, Rutherford, NSW, Australia). Single-cell suspensions were resuspended in aliquots of ≤106 cells in 100 µL BD stain buffer (BD Biosciences, Franklin Lakes, NJ, USA). After blocking for 15 min at 4°C, cells were incubated for 30 min with an antibody cocktail containing Pharmingen™ PE Rat anti-CD11b (#553311, BD Biosciences), FITC anti-mouse F4/80 (#123108, BioLegend, San Diego, California, USA), Horizon™ BV650 Rat anti-mouse CD86 (#564200, BD Biosciences), and Pharmingen™ Alexa Fluor® 647 Rat anti-mouse CD206 (#565250, BD Biosciences). All samples were then washed and fixed with BD fixation buffer (BD Biosciences). Samples were run on a BD LSRFortessa™ X-20 Analyser (BD Biosciences), and list mode data files were analyzed with FlowJo™ version 10.5.3 software. A sequential gating strategy was used to identify populations expressing CD11b+/F4/80+ total macrophages, CD11b+/F4/80+/CD86+M1-like macrophages and CD11b+/F4/80+/CD206+ M2-like macrophages. Percentage of total macrophages was calculated from the total cell population and percentage of M1-like and M2-like macrophages were calculated from the total macrophage population.
DiO-labeled reconstituted high-density lipoproteins
Discoidal rHDL were fluorescently labeled by complexing PLPC with the fluorescent membrane dye 3,3-dioctadecyloxacarbocyanine, perchlorate (DiO) powder (Life Technologies, Carlsbad, CA, USA) in a 100:1 ratio as previously described.4,13,15,17,19,23 Thereafter, the preparation was the same as for rHDL. Photo bleaching was prevented by covering with aluminum foil.
Tracking cellular association of DiO-rHDL in vitro
Immortalized murine bone marrow-derived macrophages (iBMDMs) were kindly provided by Dr. Ashley Mansell (Hudson Institute of Medical Research, Victoria, Australia). iBMDMs were maintained in low glucose Dulbecco’s modified Eagle’s medium (D5523, Sigma-Aldrich) supplemented with 10% FBS (Sigma-Aldrich) in a 37°C, 5% CO2 incubator. iBMDMs were incubated with DiO-rHDL for 2 h. Cells were then visualized and imaged using fluorescence microscopy at 484 nm, 20× magnification.
Flow cytometry for measuring DiO-rHDL cellular association in vitro
To track fate of DiO-rHDL, DiO fluorescence for measuring DiO-rHDL cellular association was detected at 484 nm. Gating was performed based on light scatter morphology to exclude cell debris and expression thresholds were set at fluorescence intensities of unstained and fluorescence minus one control.
In vivo imaging system imaging
Mice were anesthetized using 5% isofluorane in 100% oxygen (flow rate 1L/min) before being placed in the in vivo imaging system (IVIS) chamber and maintained under anesthesia using 2.5% isofluorane inside the IVIS chamber. DiO-rHDL or PBS was applied to the wound site, while the mice were under anesthesia. The fluorescent signal was acquired using the excitation/emission range of 484/501 nm. The images were captured immediately, 5, 10, 20, 40, 60, 75, and 90 min and 4, 24, and 48 h post-DiO-rHDL application. The average wound radiance (avg radiance [p/s/cm≤/sr]) was calculated using Live Image (version 4.4) software.
Wound section preparation for DiO-treated wounds
Wounds were excised and cut in half through the center of the wound. The wound was carefully placed into the Tissue-Tek cryomolds and fully submerged in optimal cutting temperature (OCT) compound. Overall, 5 μm sections were air dried for at least 30 min at room temperature and kept overnight at 4°C in the dark. The wound sections were covered with antifade mounting media containing 4′,6-diamidino-2-phenylindole (to visualize nuclei). The wound sections were then imaged by Axio Scan. Z1 (Zeiss, Oberkochen Germany).
Flow cytometry for tracking DiO-rHDL fate in wound cell types
To determine the cellular association of DiO-rHDL with different wound cell types, flow cytometry was performed on wound tissue single-cell digests incubated with an antibody cocktail containing Pharmingen™ APC-Cy™7 Rat anti-CD11b (#557657, BD Biosciences), Brilliant Violet 785™ anti-mouse F4/80 (#123141, BioLegend), PE/Cyanine7 anti-mouse CD144 (VE-cadherin) (#138015, BioLegend), Pharmingen™ PerCP-Cy™5.5 Rat anti-mouse CD31 (#553014, BD Biosciences), Pharmingen™ PE Rat anti-mouse CD90.2 (#553014, BD Biosciences) and Horizon™ BV421 Rat anti-mouse CD34 (#562608, BD Biosciences). DiO fluorescence was detected at 484 nm. A sequential gating strategy was used to identify wound cell populations including CD11b+F4/80+ macrophages, CD144+CD31+ endothelial cells, CD90.2+ fibroblasts, and CD34+ keratinocytes. Percentage of DiO-rHDL+ macrophages, endothelial cells, fibroblasts, and keratinocytes were calculated from the total DiO-rHDL+ cell population.
Statistical analysis
Data were analyzed on GraphPad Prism software (v7.0, Software MacKiev, California, USA). All results are expressed as mean ± standard error of the mean. All data were compared using either an paired student t-test, one-way or two-way analysis of variance (ANOVA), followed by post hoc analysis using Tukey’s multiple comparison tests with p < 0.05 considered statistically significant.
RESULTS
Topical rHDL rescue diabetes-impaired wound closure in a murine model of wound healing
We first sought to determine the effect of rHDL on the rate of wound closure in a type 1 diabetic murine model of wound healing (Fig. 1). The body weights of STZ-rendered diabetic mice were significantly lower compared to their nondiabetic (ND) controls (diabetic: 22.0 ± 2.1 g versus ND: 26.5 ± 1.3 g, p < 0.01). PBS-treated wounds in diabetic mice had significantly impaired wound closure at day 5, day 6, and day 7, when compared to ND PBS-treated control wounds (day 5: 94%, p < 0.001; day 6: 69%, p < 0.05; day 7: 80%, p < 0.001). Daily topical application of rHDL of diabetic wounds significantly increased wound closure at day 6 and day 7 (day 6: +288%, p < 0.01; day 7: +639%, p < 0.0001) when compared with diabetic PBS-treated wounds. Consistent with our previous study, 4 topical rHDL treatment rescued diabetes-related impairment in wound closure back to the level of ND control wounds. rHDL treatment in ND mice also significantly increased the rate of wound closure at day 7, when compared with PBS-treated ND wounds (30%, p < 0.0001).
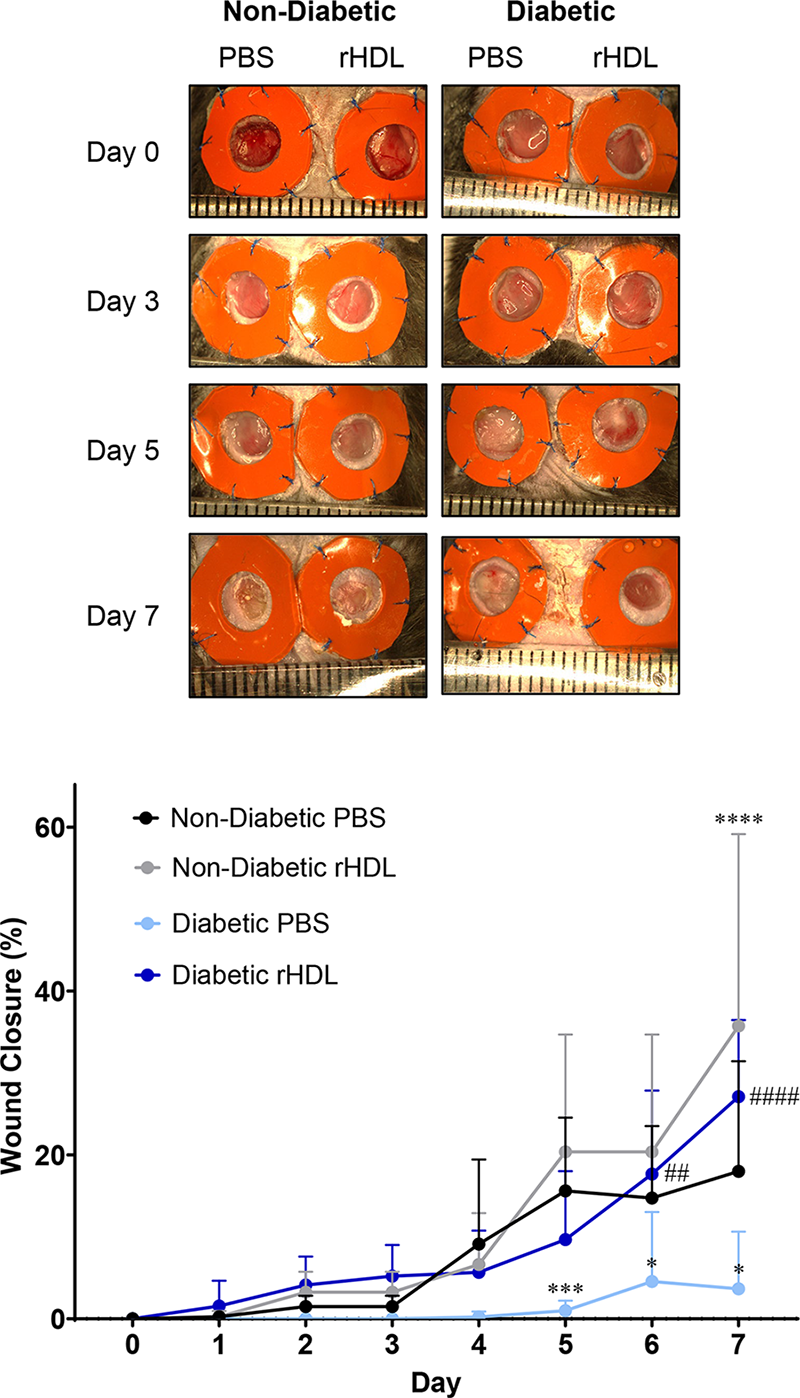
rHDL rescue diabetes-impaired wound closure in a murine model of wound healing. Two full-thickness wounds were created on the back flanks of ND (n = 9) and diabetic (n = 10) mice. Mice received daily topical applications of rHDL (50 μg/wound) or PBS (vehicle) for 7 days. Representative images of wounds at days 0, 3, 5, and 7 from ND and diabetic mice. Wound diameter measurements along the x-, y-, and z-axes areas were measured daily with micro-callipers. A calculation for wound area was performed from the average of three daily diameter measurements along the x-, y-, and z-axes. Wound closure is expressed as a percentage of initial wound area at day 0. Black: ND PBS wounds; Light blue: ND rHDL wounds; Gray: diabetic PBS wounds; Navy: Diabetic rHDL wounds. All data are expressed as mean ± SD. Individual data points are noted in Supplementary Table S1. *p < 0.05, ***p < 0.001, ****p < 0.0001 versus ND PBS; ##p < 0.01, ####p < 0.0001 versus diabetic PBS by two-way ANOVA test with Tukey’s multiple comparison post hoc. ANOVA, analysis of variance; ND, nondiabetic; PBS, phosphate-buffered saline; rHDL, reconstituted high-density lipoproteins; SD, standard deviation.
rHDL elicit changes in the NF-κ B pathway 72 h postwounding
We first examined the effect of rHDL on key mediators of the NF-κB signaling pathway at two time points: 24 and 72 h postwounding (Fig. 2). Rela is the gene that transcribes the p65 active subunit of NF-κB, a pivotal transcription factor that activates a host of inflammatory genes.8,9 Rela mRNA levels were significantly higher in the 72-h diabetic PBS wounds, when compared to both the 24-h diabetic PBS wounds (143%, p < 0.0001) and 72-h ND PBS wounds (153%, p < 0.001, Fig. 2A). However, topical administration of rHDL for the 72-h period prevented this induction in Rela mRNA. Seventy-two-hour diabetic rHDL wounds had significantly lower Rela mRNA levels (65%, p < 0.001). Gene expression of Nfkb1, which transcribes the p50 subunit of NF-κB, was threefold higher in diabetic PBS wounds at 72 h when compared with ND wounds (p < 0.05, Fig. 2B). This induction in Nfkb1 was attenuated in the rHDL-treated diabetic wounds, not reaching statistical significance compared to the ND wounds. We then explored the effects of rHDL on the IκBα protein, which binds both the p65 and p50 subunits of NF-κB, sequestering them in the cytosol and preventing them from reaching the nucleus. In the 24-h wounds, IκBα inhibitor protein was significantly elevated in both PBS and rHDL-treated diabetic wounds (PBS: threefold, p < 0.01, and rHDL: fivefold, p < 0.05, Fig. 2C), compared with the ND PBS wounds. In wounds from the 72-h time point, IκBα protein was threefold higher in the diabetic rHDL wounds compared to the ND PBS wounds (p < 0.05). There was, however, no significant difference between the PBS ND and the PBS diabetic wounds, suggesting rHDL supports the elevation of the IκBα inhibitor. Diabetic PBS wounds had significantly higher phosphorylated:total p65 protein (fourfold, p < 0.01, Fig. 2D) levels compared with ND PBS wounds at 24 h, with levels decreasing at 72 h. There were, however, no differences in phosphorylated p65 protein between PBS and rHDL-treated diabetic wounds.
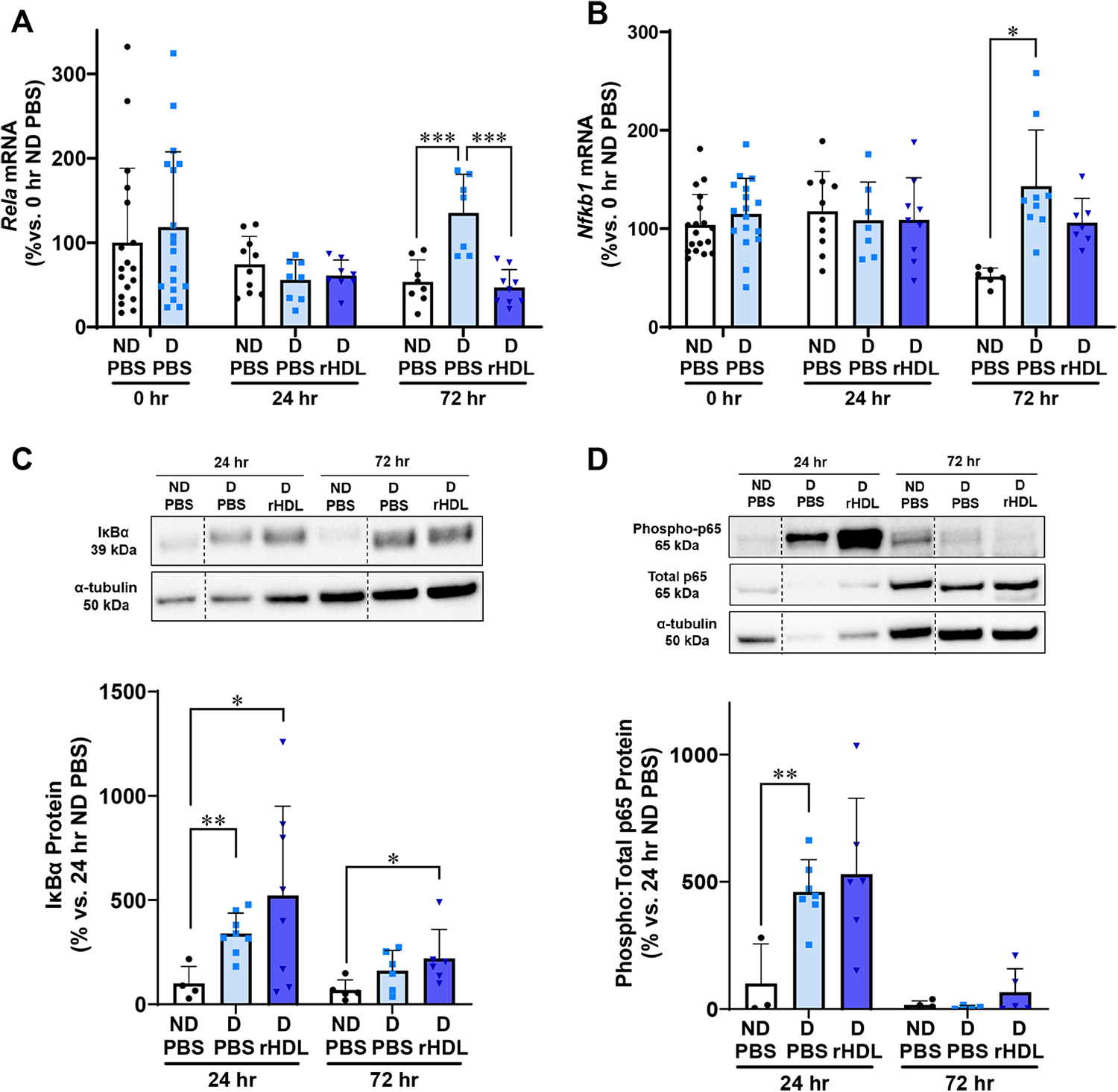
rHDL regulate changes in the NF-κB signaling pathway 72 h postwounding. Two full thickness wounds were created on the back flanks of ND and diabetic (D) mice. Mice received daily topical applications of rHDL (50 µg/wound) or PBS (vehicle) for up to 3 days. RNA was isolated from wound tissues then subjected to qPCR analyses for the measurement of:
rHDL elicit anti-inflammatory effects on Ccl2/CCL2, Ccl5, and Icam1 in diabetic wounds 72 h postwounding
We next measured inflammatory mediators including chemokines (CCL2 and CCL5) and cell adhesion molecules (VCAM1 and ICAM1). The inflammatory chemokines Ccl2 and Ccl5, play a key role in multiple aspects of wound healing. 24 There was a significant elevation in wound Ccl2 mRNA levels in the 24-h diabetic PBS (28-fold) and rHDL (27-fold) wounds when compared with the 0-h diabetic PBS wounds (p < 0.001, Fig. 3A). This elevation then declined after 72-h. Importantly, within the 72-h cohort, there was a significant reduction in Ccl2 mRNA levels in diabetic rHDL wounds, when compared with the diabetic PBS wounds (63%, p < 0.05).
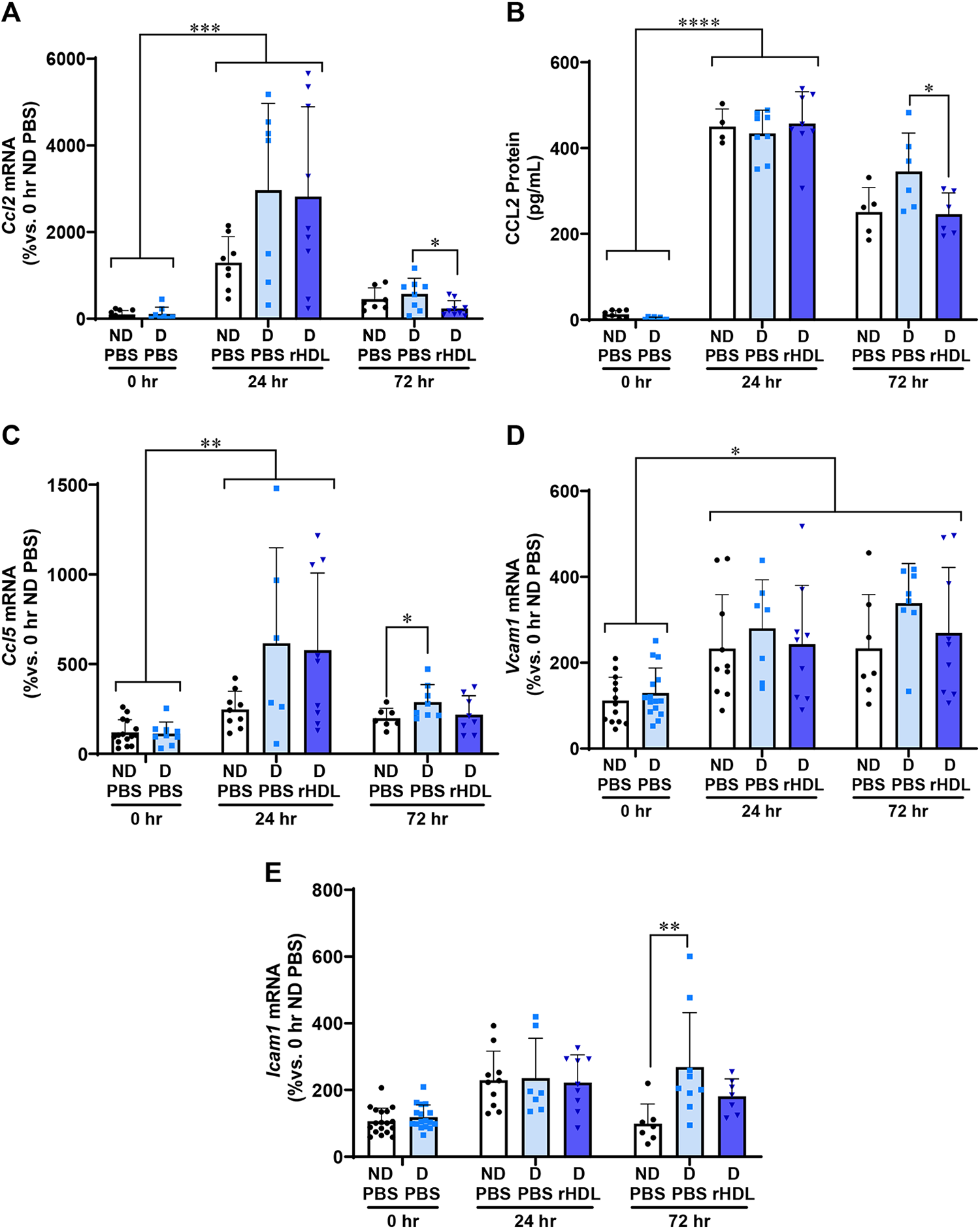
rHDL decrease inflammatory mediators Ccl2/CCL2, Ccl5, and Icam1 in diabetic wounds 72 h postwounding. Two full thickness wounds were created on the back flanks of ND and diabetic
We observed a similar pattern of expression for wound CCL2 protein (Fig. 3B). Consistent with the mRNA findings, there was a significant 20-fold increase in CCL2 protein 24 h postwounding in diabetic PBS and rHDL wounds, when compared with 0-h diabetic wounds (p < 0.0001). At the 72-h time point, diabetic rHDL wounds had lower CCL2 protein levels (29%, p < 0.05), when compared with diabetic PBS wounds.
Diabetes strikingly increased Ccl5 by at least sixfold in both diabetic PBS- and rHDL-treated wounds at 24 h (p < 0.01, Fig. 3C) then declined at 72 h. Direct comparison of the 72-h wounds showed that Ccl5 was significantly higher in diabetic PBS wounds (p < 0.05) compared to ND PBS wounds. However, this increase was not seen in diabetic rHDL wounds, suggesting an attenuation in the induction of Ccl5 by rHDL in diabetes.
Cell adhesion molecules including Vcam1 and Icam1 play a vital role in wound healing and tissue repair to enable cell migration, protein production, and proliferation. 25 Wound Vcam1 mRNA levels were significantly higher at both 24 and 72 h postwounding when compared with the 0-h time point across all groups (p < 0.05); however, no differences were observed within treatment groups (Fig. 3D). At the 72-h time point, there was a significant increase in Icam1 mRNA levels in the diabetic PBS wounds compared with ND PBS wounds (2.5-fold, p < 0.01, Fig. 3E). By contrast, there were no significant increases in Icam between ND PBS and diabetic rHDL-treated wounds, suggesting rHDL attenuated the induction of Icam in diabetic wounds.
rHDL have no effect on wound Tgfb1/TGF-β 1 or Il6/IL-6
The mRNA levels of cytokines Tgfb1 and Il6 were next tracked in diabetic and ND wounds with rHDL treatment (Fig. 4). No changes were found in wound Tgfb1 mRNA levels between diabetic or ND mice or with rHDL treatment over time (Fig. 4A). At the 0-h baseline, TGF-β1 protein levels were significantly lower in diabetic mice than ND mice (83%, p < 0.05, Fig. 4B). Topical rHDL treatment did not cause significant changes in TGF-β1 wound protein levels between groups.
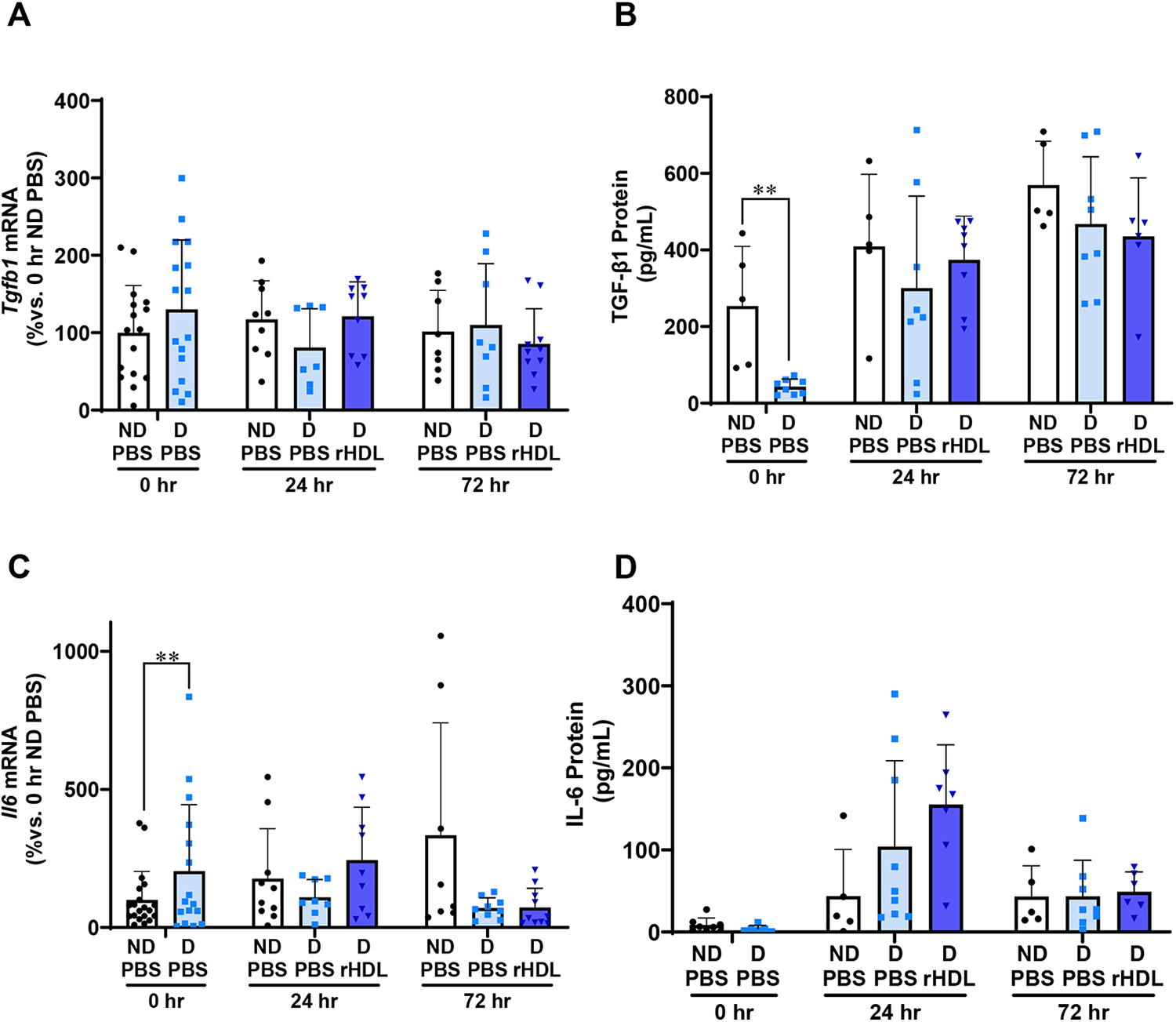
rHDL have no effect on wound Tgfb1/TGF-β1 or Il6/IL-6. Two full-thickness wounds were created on the back flanks of ND and Diabetic
Wound Il6 mRNA levels were significantly higher in diabetic mice compared to ND mice at 0-h baseline (twofold, p < 0.01, Fig. 4C). No other differences were found in wound Il6 mRNA levels between diabetic and ND mice or with rHDL treatment. Consistent with this, no changes were observed in the protein levels of wound IL-6 following topical rHDL treatment.
rHDL have no effect on wound macrophages
We next evaluated the effect of rHDL on total wound macrophages and changes in M1-like and M2-like macrophage populations at day 7 postwounding (Fig. 5). We found that wound macrophages were significantly higher in diabetic mice for both PBS-treated and rHDL-treated wounds (PBS: 78%, p < 0.05; rHDL: 102%, p < 0.05, Fig. 5A), when compared with their respective ND wounds. rHDL did not, however, significantly change the number of wound macrophages. When examining the CD86+ (Fig. 5B) and CD206+ (Fig. 5C) wound macrophage populations, no changes were found with the induction of diabetes or following the application of rHDL.
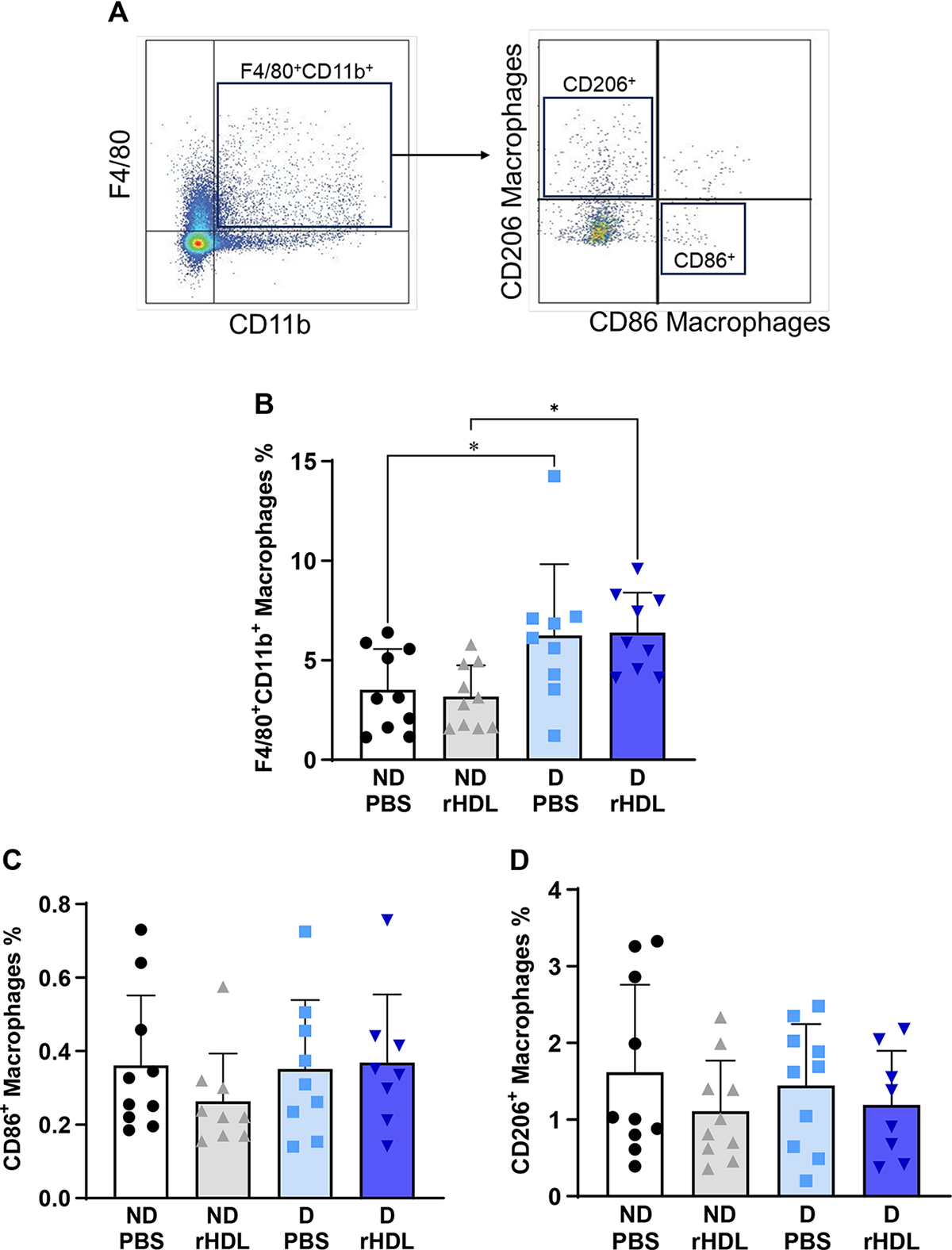
rHDL have no effect on wound macrophages. Two full-thickness wounds were created on the back flanks of ND and diabetic
Detection of fluorescently labeled DiO-rHDL in macrophages in vitro and in wounds in vivo
Fluorescently labeled DiO-rHDL were used to detect the cellular association and retention of topically applied rHDL in murine wounds (Fig. 6). We first confirmed the association of DiO-rHDL with iBMDMs in vitro as its detection by fluorescence was visualized using fluorescent microscopy (Fig. 6A) and confirmed further using flow cytometry (Fig. 6B).
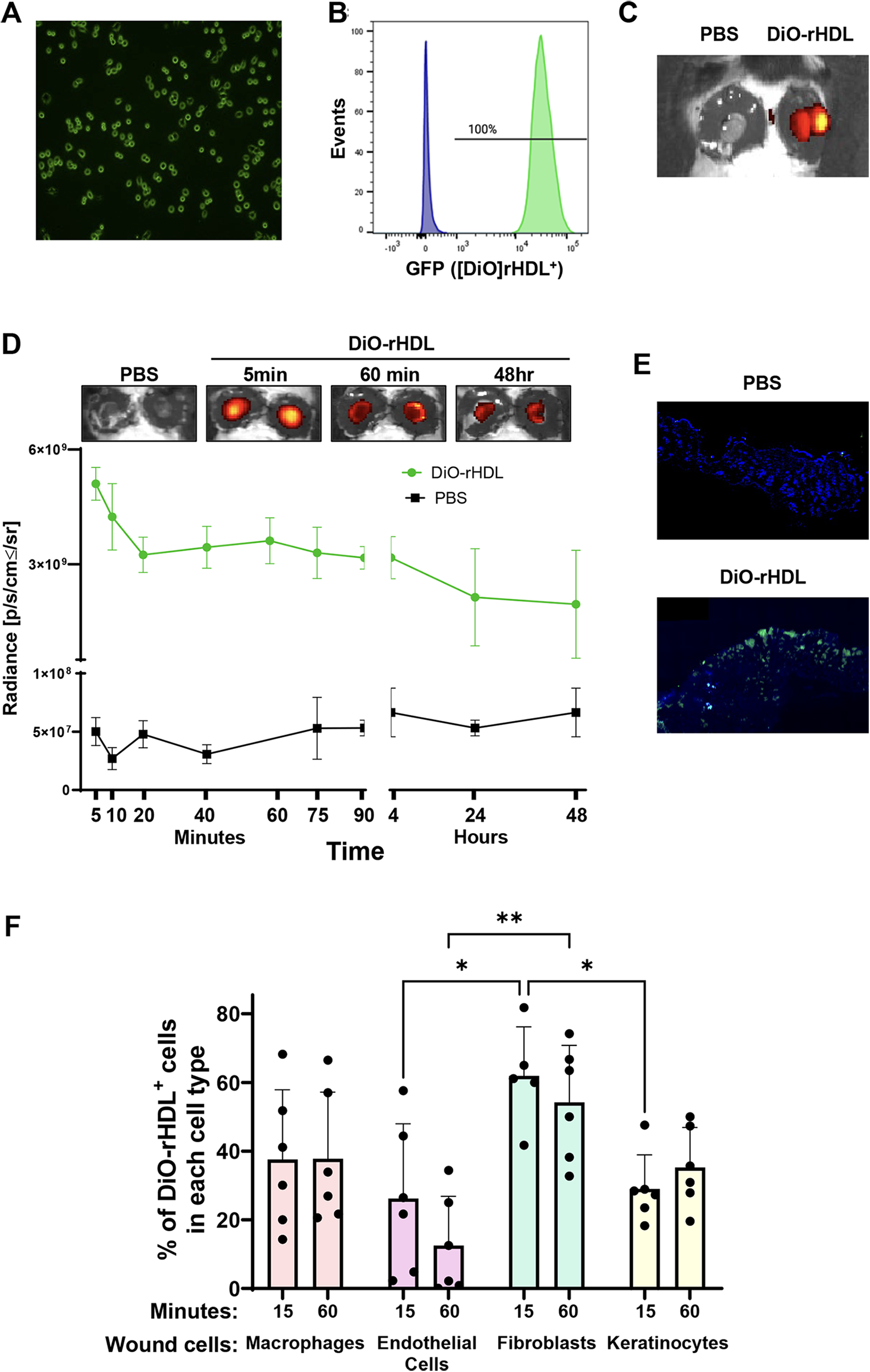
Detection of fluorescently labeled DiO-rHDL in macrophages in vitro and in wounds in vivo. Immortalized bone marrow-derived macrophages were incubated with rHDL fluorescently labeled with DiO dye (DiO-rHDL) for 2 h for
Next, using whole-body fluorescence imaging by IVIS, we confirmed that the DiO-rHDL were retained solely within the DiO-rHDL wound, with no radiance detected in the PBS-treated wound of the same animal (Fig. 6C). Next we tracked the duration that DiO-rHDL are retained within the wounds (Fig. 6D). We found that radiance from the DiO-rHDL was substantially higher in DiO-rHDL-treated wounds 5 min post-topical application than PBS-treated control wounds (100-fold, Fig. 6D). This radiance declined over the first 20 min by 36% and then remained relatively constant until a further decline between 4 and 24 h (58%), where it was stable out to the 48-h time point. Using fluorescent microscopy on wound cryosections, DiO-rHDL were also detected in wounds treated with DiO-rHDL, but not in PBS-treated wounds (Fig. 6E).
We next determined the wound cell types that take-up DiO-rHDL in the first 60 min post-topical application. Fifteen minutes after DiO-rHDL application, the fibroblasts had the greatest cellular association with DiO-rHDL, followed by macro phages, endothelial cells, and keratinocytes (Fig. 6F). DiO-rHDL cellular association was significantly less in the wound keratinocyte and endothelial cell populations after 15 min than fibroblasts (p < 0.05 for both). At 60 min postapplication, DiO-rHDL were lower in endothelial cells compared to fibroblasts (p < 0.01).
DISCUSSION
Inflammation is critical in the early stages of wound healing to combat infection, however, prolonged and inappropriate inflammation prevents the progression of wound healing and results in nonhealing, chronic wounds. Diabetes, in particular, is associated with persistent and nonresolving inflammation leading to the delay in DFU healing.6,26 rHDL have been shown to rescue diabetes-impaired wound healing through the promotion of angiogenesis. 4 Despite well-established anti-inflammatory properties,13,15–17,23,27 the effect of rHDL on diabetic wound inflammation has not been previously explored and may contribute to their wound healing benefits. In this study we found that topical application of rHDL: (1) increased the rate of wound closure, consistent with our previous findings 4 ; (2) reduced inflammatory markers Rela, Nfkb1, Ccl2, and CCL2 and attenuated the induction of Ccl5 and Icam1 in diabetic wounds 72 h postwounding; (3) had no effect on wound macrophage content or polarity; and (4) using DiO-rHDL, we found rHDL are retained in the wound for up to 48 h. Our findings provide further mechanistic insight into the known wound healing benefits of rHDL by eliciting anti-inflammatory effects in the early-mid stages post healing in diabetic wounds.
rHDL have well-established anti-inflammatory properties that have been demonstrated in vitro in endothelial cells, monocytes/macrophages, and smooth muscle cells,13,15–17,23,27 key cell types involved in the wound healing process. The anti-inflammatory properties of rHDL have also been demonstrated in vivo in vascular inflammation models.13,14,27 The central mechanism for the anti-inflammatory effects of rHDL is driven by its ability to inhibit the pivotal inflammatory transcription factor NF-κB. NF-κB controls the regulation of inflammatory chemokines (CCL2, CCL5), cell adhesion molecules (VCAM-1, ICAM-1), and cytokines (TGF-β1, IL-6). 13 Our study showed that gene expression of Rela and Nfkb1, which transcribes the p65 active and p50 subunits of NF-κB, respectively, were significantly higher in 72-h diabetic wounds, compared to ND control. However, rHDL treatment attenuated this induction. Consistent with this, we found that rHDL also attenuated diabetes-induced increases in Ccl2, Ccl5, and Icam1 at the 72-h time point and supported an increase in the IκBα inhibitor that sequesters p65 and p50 in the cytoplasm preventing NF-κB activation. Interestingly, rHDL did not elicit any effects on phosphorylated p65 levels. We, therefore, hypothesize that rHDL reduce the transcription of NF-κB subunits and prevents the translocation of NF-κB to the nucleus via an increase in IκBα. This occurs without an increase in phosphorylated (activated) p65, although we find reductions in inflammatory mediators. This anomaly may be a result of the time frame in which these findings were captured in wounds postwounding. Overall our findings support previous studies showing the reduction of inflammatory mediators following rHDL treatment.13,15–17,23,27
We did not, however, observe changes in IL-6 and TGF-β1 in diabetic wounds at either the mRNA or protein level. One possible explanation for this is that transcription of Ccl2 is primarily driven by NF-κB. 28 However, the transcriptional regulation of the Il6 gene is more complex and involves four different transcription factors, including NF-κB, activator protein-1, CCAAT/enhancer binding protein, and cAMP response element-binding protein. 29 There is no current evidence that these additional factors are affected by rHDL and may explain why rHDL have not elicited a change in Il6 gene expression. TGF-β1 is involved in a number of processes in wound healing: inflammation, stimulating angiogenesis, fibroblast proliferation, collagen synthesis, and deposition and remodeling of the new extracellular matrix.30–32 Defective TGF-β1 signaling contributes to delayed wound healing in diabetes. 33 HDL has previously been shown to induce TGF-β2, but not TGF-β1 or TGF-β3, levels in endothelial cells. 34 Our study found that HDL did not affect TGF-β1 expression.
The reduction in wound Rela and Ccl2/CCL2 took 72 h to come into effect following daily topical application of rHDL. Using fluorescently labeled DiO-rHDL, we demonstrated that rHDL are retained in the wound for a prolonged period following application, predominantly for the first hour, but could be visualized in wound tissue sections after 4 h by fluorescence microscopy and was retained at low levels out to 48 h postapplication. This suggests that rHDL do not enter the circulation immediately and has the opportunity to interact with wound cells such as endothelial cells, macrophages, fibroblasts, and keratinocytes. Consistent with this, we found that DiO-rHDL were taken up by all these cell types within the first hour of application. The prerequisite for an anti-inflammatory effect appears to be three repeat daily doses over 72 h. In vitro studies with rHDL that investigate its anti-inflammatory properties predominantly use incubation periods of 16–24 h. This is perhaps an equivalent accumulative time frame to our in vivo application in wounds when taking into consideration the loss of rHDL retained in the wound over time. Furthermore, a reduction in inflammation at 72 h postwounding is perhaps an optimal time point. It is just after the earliest stages when inflammation is needed to fight infection and in time to prevent excessive inflammatory cell accumulation in the mid-stage that risks development of a chronic nonhealing wound. Supporting this is our finding of an increased rate of wound closure in rHDL-treated diabetic wounds.
Our study showed a significant increase in the total macrophage population in diabetic wounds, compared to ND wounds 7 days postwounding, consistent with previous literature.7,11 There were, however, no changes in the total number of wound macrophages in rHDL-treated wounds. This was an unexpected result, yet in studies of atherosclerosis, the more common application of rHDL, infusions of rHDL also show either very modest or no effect on plaque macrophage content and this is despite reductions in inflammatory chemokines, 13 similar to the current study. Based on our data showing that rHDL reduce diabetic wound inflammation, it may have been anticipated that there would also be fewer wound macrophages. CCL2 is an important chemokine that recruits monocytes from the circulation into the neighboring wound tissues. 24 However, other chemokines can also perform this function including CCL5 and C-X3-C motif chemokine ligand 1 24 which are released by endothelial cells 13 suggesting that the inflammatory response may have been exerted primarily by other cell types such as endothelial cells rather than monocytes. In contrast to previous findings,35,36 we did not observe an imbalance between CD86+ M1-like or CD206+ M2-like macrophage populations with diabetes. It is possible that we may not be capturing the correct timing of the inflammation process. Our studies were performed 2.5 weeks after mice were rendered diabetic by STZ. In previous studies, the changes in M1 and M2 macrophages were seen in STZ-induced mice that had hyperglycemia for a longer duration (at least 1 month post-STZ before wounding).35,36
rHDL have previously been shown to drive a shift from M1-like to M2-like macrophages in vitro, with a significant increase in the expression of M2-like markers Arginase-1 and Fizz-1. 18 In contrast, an ex-vivo study found HDL did not influence macrophage polarization in human monocyte-derived macrophages. 37 We observed no differences in M1 or M2 macrophage phenotypes or the cytokines they express (IL-6 or TGF-β1, respectively). Taken together, it is possible that in a biologically complex in vivo environment, with diverse cell types that coinhabit the wound with macrophages, that the macrophage polarization properties of rHDL that have been demonstrated in vitro, are diluted and less effective.
Several studies have examined the wound healing effects of topical rHDL to full-thickness murine wounds,4,19 however, no study has tracked the cellular association and fate of topically applied rHDL. We labeled rHDL with the fluorescent dye DiO, previously used to track the biodistribution of spherical [S]-rHDL in murine atherosclerosis models following intravenous administration. 23 We found robust association of DiO-rHDL with macrophages in vitro and after topical application in murine wound tissue (up to 48 h). Furthermore, we found that DiO-rHDL were associated with fibroblasts, macrophages, and endothelial cells and less so by keratinocytes within the first 15 min of application. Information regarding the retention of rHDL in wounds is important for determining the required frequency of application essential for its future translation as a wound healing therapy.
There were a number of limitations in this study, which should be noted. First, while we did see a significant improvement in wound closure with rHDL treatment, we were unable to assess the extent of epithelialization as it is difficult to measure robustly in wound tissue sections. We also acknowledge that assessment of wound macrophage phenotypes needs further refinement with additional markers to better delineate the different macrophage subtypes. Additionally, we were unable to replicate the imbalance in M1/M2 macrophage populations that have been previously reported.35,36 This is likely due to the short duration of hyperglycemia that our mice endured (2.5 weeks versus 1 month post-STZ).
In conclusion, rHDL rescues diabetes-impaired wound healing and reduced inflammatory markers Rela (−56%) and Ccl2 (−63%)/CCL2 (−29%) in diabetic wounds, 72 h postwounding. Despite this, we did not observe changes in wound macrophage population number following rHDL treatment. Using DiO-rHDL, we confirmed rHDL are retained in the wound for a prolonged period of time up to 48 h. The combined anti-inflammatory and proangiogenic properties of topical rHDL in diabetic wound healing present rHDL as an effective pleiotropic treatment option for DFU.
INNOVATION
We have previously shown that HDL promotes wound healing by correcting key angiogenic signaling pathways that are impaired in diabetes. HDL has well-established anti-inflammatory properties. This study explored whether rHDL elicited anti-inflammatory effects in diabetic wounds. Diabetes prolongs wound inflammation and slows healing. For the first time, we demonstrate here that HDL exerts anti-inflammatory effects early postwounding in diabetic mice which may contribute to its significant wound healing benefits. These findings highlight the potential of topical rHDL as a novel strategy to improve wound healing in patients with DFUs and prevent lower-limb amputations.
Topical rHDL increased the rate of wound closure in diabetic mice, consistent with previous findings.
4
Topical rHDL suppressed expression of inflammatory markers Rela, Nfkb1, Ccl2, Ccl5, and Icam1 in diabetic wounds early postwounding, with no effect on wound macrophage number or polarity. Tracking of fluorescently labeled DiO-rHDL showed that DiO-rHDL were retained in the wound for up to 48 h and taken up by various wound cell types including endothelial cells, macrophages, fibroblasts, and keratinocytes.KEY FINDINGS
AUTHORS’ CONTRIBUTIONS
Z.L.: Investigation; formal analysis; methodology; visualization; writing—original draft. J.T.M.T.: Investigation; formal analysis; methodology; validation; visualization; writing—original draft; writing—review and editing. V.A.N.: Investigation. L.S: investigation. S.L.: Investigation. E.L.S.: Investigation. A.E.W.: Investigation. J.D.: Writing—review and editing. S.J.N.: Writing—review and editing. P.J.P.: Writing—review and editing. R.F.: writing—review and editing. C.A.B.: Conceptualization; formal analysis; funding acquisition; methodology; supervision; validation; visualization; writing—original draft; writing—review and editing.
Footnotes
ACKNOWLEDGMENTS AND FUNDING SOURCES
This work was supported by a Diabetes South Australia grant and a Lin Huddleston Heart Foundation fellowship to C.A.B.
AUTHOR DISCLOSURE AND GHOSTWRITING
No competing financial interests exist. The content of this article was expressly written by the authors listed. No ghostwriters were used to write this article.