
Abstract
Significance:
Volumetric muscle loss (VML) results in the loss of large amounts of tissue that inhibits muscle regeneration. Existing therapies, such as autologous muscle transfer and physical therapy, are incapable of returning full function and force production to injured muscle.
Recent Advances:
Skeletal muscle tissue constructs may provide an alternative to existing therapies currently used to treat VML. Unlike autologous muscle transplants, muscle constructs can be cultured in vitro and are not reliant on intact muscle tissue. Skeletal muscle constructs can be generated from small muscle biopsies and could be used to generate skeletal muscle tissue constructs to replace injured tissues.
Critical Issues:
To serve as effective therapies, muscle constructs must be capable of generating contractile forces that can assist the function of host skeletal muscle. The contractile force of native muscle arises in part as a consequence of the highly aligned, bundled architecture of myofibers. Attempts to induce similar alignment include applications of tension/strain across hydrogels, inducing aligned architectures within scaffolds, casting tissues in straited molds, and 3D printing. While all these methods have demonstrated efficacy toward inducing myofiber alignment, the extent of myofiber alignment, tissue formation, and force production varies. This manusript critically reviews the advantages and limitations of these methods and specifically discusses their ability to impart mechanical and architectural cues to induce alignment within tissue constructs.
Future Directions:
As tissue-synthesizing techniques continue to improve, muscle constructs must include more cell types than simply myoblasts, such as the addition of neuronal and endothelial cells. Higher-level tissue organization is critical to the success of these constructs. Many of these technologies have yet to be implanted into host tissue to understand engraftment and how they can contribute to traumatic injury, and as such continued collaboration between surgeons and tissue engineers is necessary to ultimately result in clinical translation.

SCOPE AND SIGNIFICANCE
Skeletal muscle injuries account for 77% of health care visits in the United States and are the leading cause of disability among soldiers. 1 –3 While minor skeletal muscle injuries spontaneously regenerate over several days or weeks, muscle cannot repair itself after the loss of massive amounts of tissue. These injuries, defined as volumetric muscle loss (VML), are characterized as the destruction or removal of a significant portion of muscle that results in a permanent reduction in contractile force production as well as a reduction in patient mobility and function. 4 VML is a difficult condition to treat as there is often not enough tissue left after the injury to provide either the scaffolding network or the cellular niche required for spontaneous, normal muscle regeneration, which is required to facilitate functional recovery. 5 For the purposes of this review, we define functional recovery as the ability for skeletal muscle to recover contractile and metabolic function, restoring mobility to the patient.
TRANSLATIONAL RELEVANCE
While factors such as immune response and fibroblast activation play a significant role in mediating VML injury progression, regeneration is severely inhibited if the loss of skeletal muscle is too great. 2,6 Engineered skeletal muscle constructs with functional capabilities similar to native skeletal muscle tissue may provide an alternative to existing therapies for the treatment of VML. The rationale behind the development of these biomimetic skeletal muscle constructs is to cultivate them in vitro to ultimately be implanted directly into the defect, thus avoiding the need for multiple complex and invasive surgeries to procure and graft muscle tissue.
CLINICAL RELEVANCE
The current standard of care for VML is an autologous muscle transplant coupled with physical therapy; while these approaches can return some function to patients, there are currently no clinically available treatments that return full mobility and force production. Bracing of the injured limb is also an option to aid patient stabilization and mobility. Physical therapy can improve remaining muscle strength and reduce muscle stiffness but does not usually induce significant muscle regeneration. 7 Furthermore, autologous transplants are limited by tissue availability, surgical skill, infection, and tissue morbidity. 8 Thus, there is a need for novel therapeutics for VML that are not dependent on patient tissue or multiple, invasive surgeries.
BACKGROUND
To treat VML, tissue engineering–based solutions must (i) encourage self-regeneration of the remaining tissue, 9 (ii) replace or augment the damaged tissue with an effective substitute, or (iii) use some combination of these approaches. In this review, we summarize recent progress toward diverse approaches to engineer skeletal muscle constructs for the treatment of VML, as well as for increased understanding of muscle physiology. Many of these strategies center around the creation or induction of aligned myofibers, as this alignment directly correlates with the ability for skeletal muscle to generate force and subsequently aid in locomotion. 10 For biomimetic skeletal muscle constructs to produce similar force, the cells maturing within these tissues must mimic the hierarchal structure of native skeletal muscle and be aligned along the principle direction of force production.
DISCUSSION
Anatomy of muscle and the role of alignment in skeletal muscle function
Skeletal muscle is a complex tissue consisting of highly aligned, hierarchal contractile units (Fig. 1). This arrangement is nontrivial; myofiber alignment and arrangement is required for proper force generation and muscle function. 12 Specifically, muscle fibers are arranged relative to the axis by which forces are generated. While overarching skeletal muscle anatomy varies depending on the muscle type, the organization of myofibers within muscle follow similar, hierarchal patterns that are necessary for tissue function. 13,14 Individual myofibers are surrounded by the basal lamina/endomysium and bundle together to form fascicles. Fascicles bundle together and are surrounded by the perimysium, and the entire muscle body is sheathed by the epimysium. 14 –17 These fibrous layers are important to facilitate alignment of myofibers, which is necessary for tissue contractility and force production. Alignment at the cellular level is dependent on cytoskeletal reorganization in response to environmental stimuli. A growing body of evidence suggests that this relationship is heavily reliant on mechanical stimulation; myofiber alignment has been observed when cultured under tension as well as when cultured along straited-surfaces, as discussed further in the “Methodologies for inducing myofiber alignment” section. 18 Alignment appears to be driven by actin reorganization as well as reorganization of intermediate filaments within muscle cells. 19 Attempts to disrupt filament formation, such as mutations to desmin and keratin intermediate filaments, have resulted in myofibers with nonaligned branching morphologies that adversely impact force production and function. Together, these data suggest that alignment is dependent on both external mechanical cues and complex internal cytoskeleton organization. 20 Satellite cells (SCs), the native population of progenitor cells in skeletal muscle tissue, reside between the sarcolemma and the basement membrane/basal lamina of individual myofibers. These cells remain inactive and quiescent in mature, uninjured tissue but play a key role in muscle regeneration. 15 –17 The contractile force production and overall function of skeletal muscle are reduced when tissue structure and myofiber alignment is interrupted by major injury, myopathy, or dystrophy. 5,18
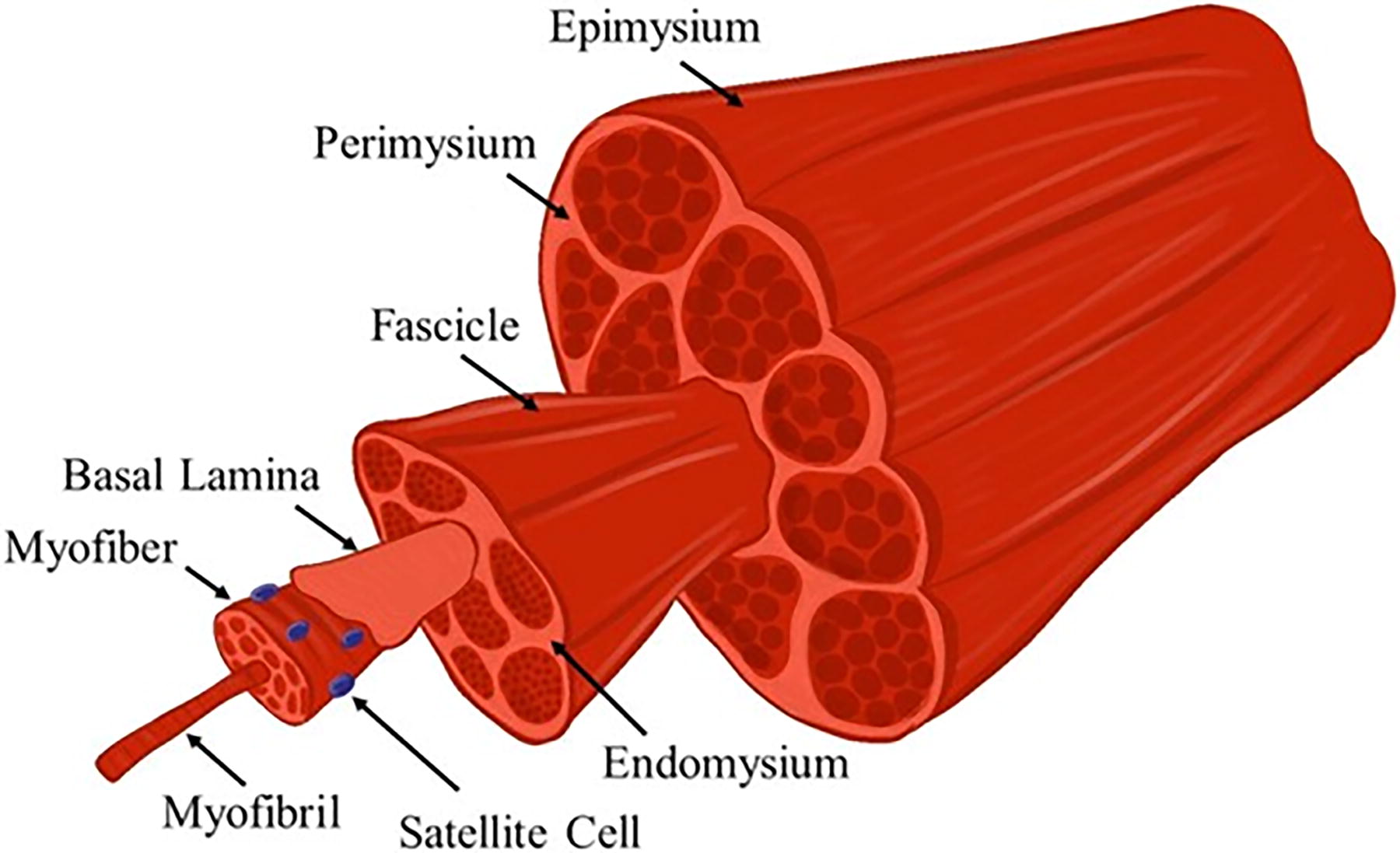
A schematic illustration of skeletal muscle anatomy. Skeletal muscle consists of highly aligned, contractile units that are bundled to form hierarchal structures to produce and transmit uniaxial force. Adapted with permission from Kozan et al. 11 Color images are available online.
Skeletal muscle regeneration in relatively small injuries is characterized by three distinct repair phases: an inflammatory phase, a repair phase, and a remodeling phase, 21,22 and the reader is referred to the following descriptive reviews for an in-depth discussion of these mechanisms. 14,16 This process is guided by mechanical and soluble cues, such as growth factors. Growth factors and cytokines are essential to muscle repair and regeneration, and their local presence, presentation, and abundance change throughout the regenerative process. While this review will highlight some key factors involved in muscle regeneration, the reader is referred to the following for more descriptive reviews of the role of specific growth factors in muscle regeneration. 16,23 –25 Briefly, damage to skeletal muscle tissue causes the sarcolemma to rupture, resulting in extracellular calcium rushing into myofibers, triggering calcium-dependent proteases to initiate myofiber necrosis. 26 Myofiber necrosis initiates the inflammatory phase of muscle regeneration, which recruits neutrophils, as well as phagocytic M1 macrophages, into the injury region. These cells phagocytize necrotic myofibers and damaged extracellular matrix (ECM) and induce myofiber degradation via the release of nitric oxide (NO). 26 –28 Approximately 3–7 days after the initial injury, M1, proinflammatory, macrophages are gradually replaced by M2, proregenerative, macrophages, marking a shift from the inflammatory phase to the repair phase. 29 The repair phase is characterized by the migration, proliferation, and differentiation of SCs into the injured region. The activation and proliferation of SCs are initiated by the release of growth factors such as hepatocyte growth factor (HGF) and fibroblast growth factor 6 from the local ECM. 15 Interestingly, the release of these factors is promulgated by several events within the inflammatory phase, such as the release of NO, which has been shown to facilitate the release of HGF. 30 After migrating into the injured region and proliferating, SCs begin their terminal differentiation and fuse to form myofibers that will ultimately mature to regenerate the force production capacity of the muscle. 26
The remodeling phase of skeletal muscle regeneration initiates as SCs begin to differentiate, and the outcome of this phase is dependent on a balance between fibroblast infiltration into the injury site with the differentiation of regenerating myoblasts. Smaller injuries result in less scar tissue deposition and are often able to fully remodel because of adequate SC infiltration and differentiation, while larger injuries result in increased fibroblast infiltration and scar tissue deposition (Fig. 2). 6 The endogenous regenerative mechanism within skeletal muscle cannot overcome the amount of tissue loss that occurs in VML injuries, as the frank loss of tissue completely removes any regenerative cues from the ECM that would otherwise be released during the repair phase. Instead, the pathophysiology of VML is characterized by an overwhelming inflammatory response facilitated by a chronic and sustained presence of macrophages. 31 These inflammatory macrophages ultimately recruit large amounts of fibroblasts, which results in the deposition of disorganized fibrotic tissue. 22 Scar tissue further interferes with regenerating myofibers to prevent integration with the surrounding musculature to limit the overall regenerative response and functional recovery. 31,32 Regeneration is further impaired by the destruction of neighboring quiescent SCs along with mature myofibers that may aid in SC maturation, fusion, and organization. 33,34 Fibroblast infiltration dominates this process, resulting in the deposition of several proteins including collagens type I and III, resulting in the formation of scar tissue rather than organized skeletal muscle. 35 These changes in cellular and protein composition at the injury site, as well as the fact that VML is characterized by the frank loss of tissue that cannot be recovered, further exacerbate the pathophysiology of VML, facilitating a permanent loss of function of the affected muscle. Therefore, tissue-engineered strategies either need to enhance endogenous regeneration pathways or implant mature muscle that can integrate with the remaining host musculature to bypass the injury response altogether.
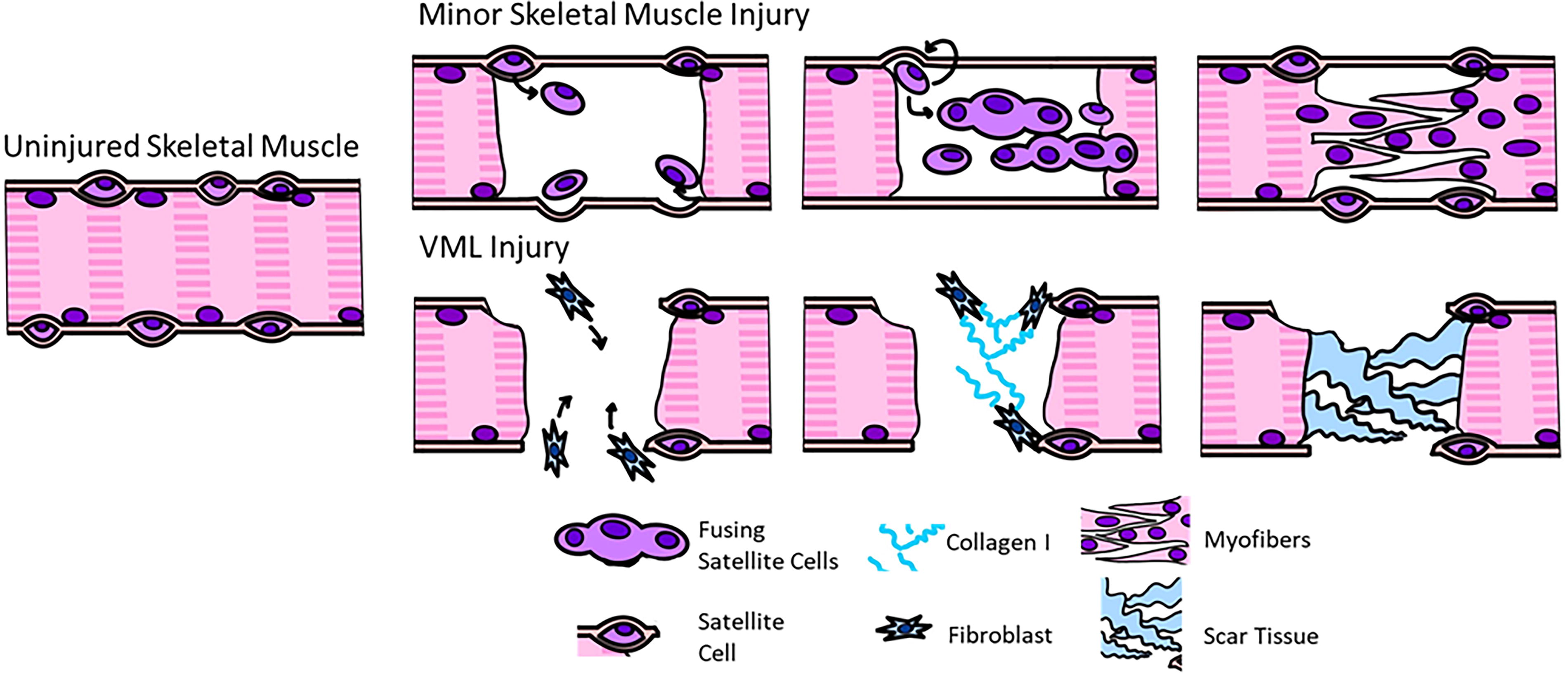
Schematic diagram of repair and regeneration phase of minor muscle injuries and VML injuries. In minor injuries, skeletal muscle is capable of regenerating myofibers. In VML injuries, muscle does not result in muscle fiber regeneration, and instead disorganized scar tissue is deposited within the region of injury. VML, volumetric muscle loss. Color images are available online.
Tissue-engineered strategies for skeletal muscle tissue engineering
Tissue-engineered approaches for the treatment of VML broadly focus on two separate approaches: acellular therapeutics that encourage the development of nascent skeletal muscle tissue within the injury site (Fig. 3A) or the implantation of cell-laden engineered skeletal muscle tissue constructs (Fig. 3B). Acellular strategies for the treatment of VML utilize a biomaterial construct with optimized structural, mechanical, and biochemical properties to induce SC migration and ultimately support myofiber formation within the defect site. Common materials used in the fabrication of acellular scaffolds are natural biomaterials such as collagen, fibrin, alginate, and decellularized ECM and synthetic materials such as poly(lactic-co-glycolic acid) (PLGA) and poly-ε-caprolactone (PCL). The reader is referred to the following for more comprehensive reviews on these topics. 11,14,36,37
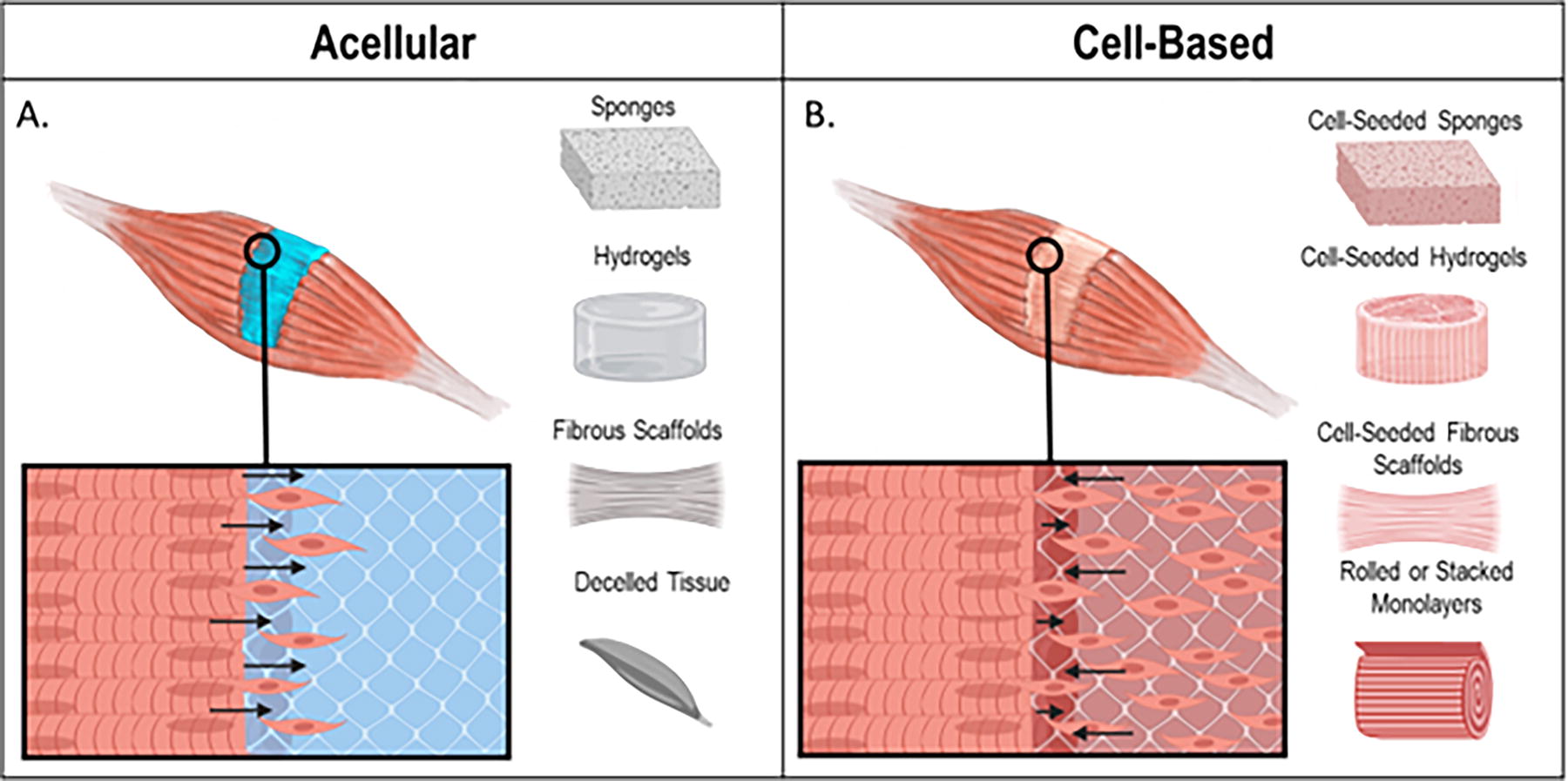
Representative diagram of differences between acellular and cell-based therapies for VML.
Cell-laden strategies for the treatment of VML can be defined as any approach where cells are incorporated into a biomaterial prior to implantation or in vitro study. Various methodologies have been employed in the generation of engineered muscle constructs, encompassing approaches such as seeding myoblasts into hydrogels, construction of 3D muscle units through stacking or rolling 2D myofiber monolayers, myoblast seeding into decellularized tissue, 3D bioprinting, and combinations thereof. 37 –43 Each strategy has various benefits and limitations, but the overarching goal remains the same: to engineer functional, biologically relevant skeletal muscle tissue. In order to create such, these engineered tissues must be structurally similar to native skeletal muscle, strong enough to survive implantation, and able to generate contractile force similar to native skeletal muscle tissue. The following sections will introduce common methods utilized to induce alignment within engineered skeletal muscle tissues.
Methodologies for inducing myofiber alignment
Induction of myofiber alignment with tension
A common method to induce myofiber alignment is to culture myoblast-seeded hydrogels under tension, these approaches are summarized in Table 1. These methodologies are reflective of in situ development of muscle tissue, where strain and tension each play a key role in the directional growth and maturation of embryonic skeletal muscle. 18 Generally, models that utilize tension consist of systems where hydrogels are cast around posts (Fig. 4D), 18,44,48 “fused” to the edges of a mold, 46,47 or suspended under tension via clamps (Fig. 4A–C). 49,50 Tension models can be further split into two categories: static and dynamic. Static systems induce tension passively by allowing tissues to compact between two or more points of restraint. Dynamic systems further generate strain by actively stretching tissues in a bioreactor. 51 Multiple studies have shown that myofiber arrangement and alignment can be modified by varying the shape and geometry of fabricated anchoring points that hold the tissue under tension. 46,52 Early work by Bian et al. demonstrated that culturing C2C12 loaded collagen or fibrin gels around micropatterned polydimethylsiloxane (PDMS) molds with elongated hexagonal posts resulted in improved myofiber alignment and bundling between posts, suggesting that post geometry could be used to control alignment and cell orientation. 44 Post number also impacted fiber alignment; tissues suspended between multiple cylindrical posts resulted in increased myofiber alignment than when cast around a single post, suggesting that the strain fields generated by these connection points influence myofiber organization. 18 Cell alignment can also be impacted by post orientation; myofiber alignment and elongation were significantly increased when elongated hexagonal posts were oriented 90° to one another rather than when the posts were arranged in parallel. 44 Collectively, these studies demonstrate that myofibers align in the presence of tension and are sensitive to strain distributions across the tissue. Varying post morphology and orientation can manipulate strain fields within a tissue and, since myofibers develop along regions of higher tension, may allow for the creation of tissues with unique myofiber organization and orientation.
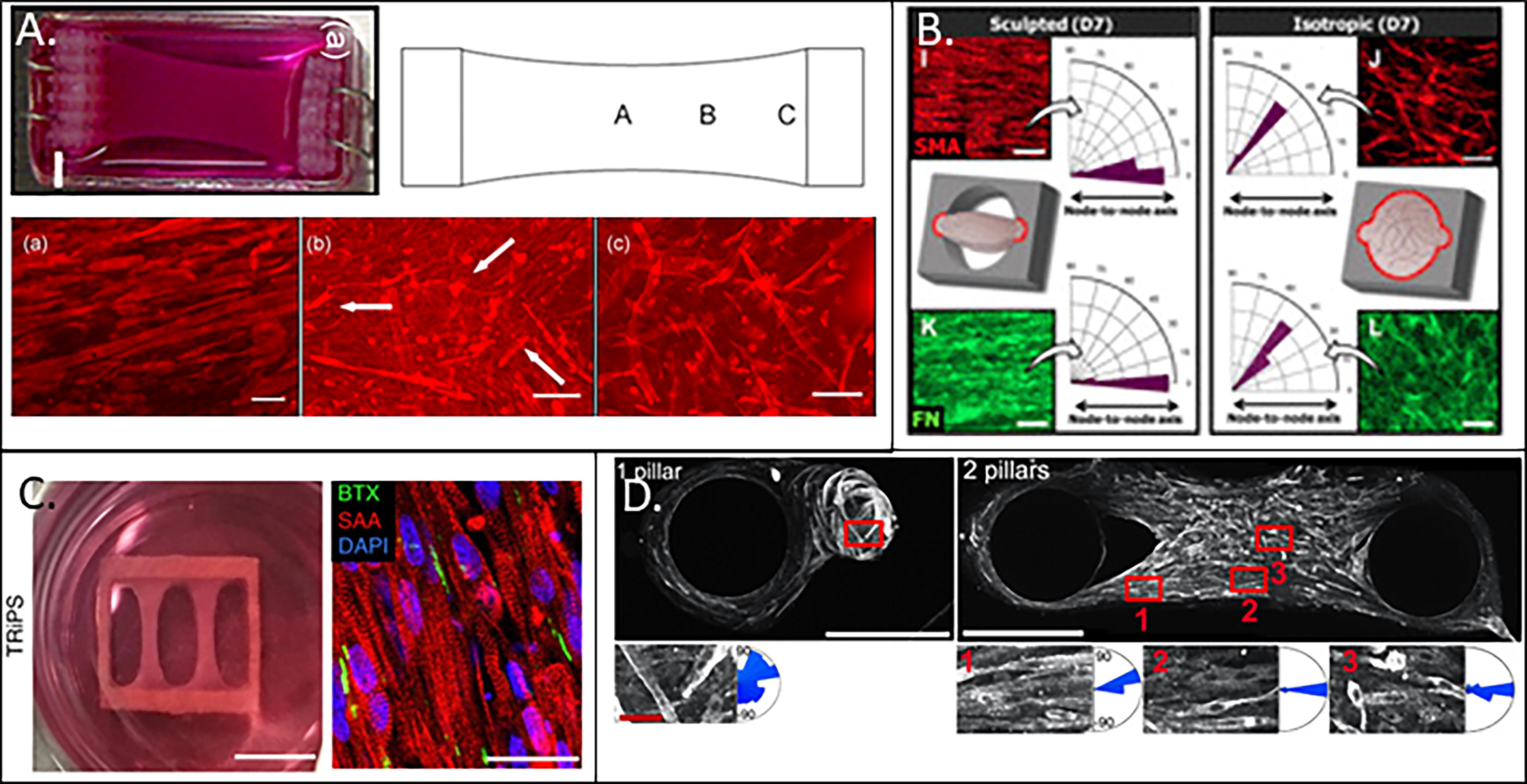
Myofiber alignment as a consequence of tension across a variety of different designs.
Summary of approaches that utilize tension to induce myofiber alignment
PDMS, polydimethylsiloxane; iPSC, induced pluripotent stem cells.
A separate approach that results in consistent, uniaxial alignment throughout the entire tissue involves clamping or “fusing” the tissue construct along the entire width of a supporting structure rather than at a single point. 46,47 Primary human myoblast-seeded fibrin hydrogels were fused along the edges of a 9 × 9 mm2 rigid frame using Velcro to generate a suspended construct (Fig. 4C). 47 The authors reported dense, highly aligned myofibers after 2 weeks in culture that exhibited twitch-to-tetanus ratios similar to primary myofibers and adult human skeletal muscle tissue when electrically stimulated. 47 Anisotropic myofiber alignment was also observed by Mondrinos et al. in human mesenchymal stem cell–seeded collagen gels that were anchored in 1-mm-thick PDMS nodes consisting of an ellipsoid chamber with anchorage nodes on either end of the chamber. Tissues in this anchored environment expressed significantly elevated levels of myogenin than control tissues cultured under isotropic conditions (Fig. 4B). 46 These results suggest that the utilization of clamps or parallel attachment points can result in a uniform strain field and that the application of uniform strain results in tissues with more consistent alignment.
In contrast to static systems, dynamic systems generate strain by stretching tissues utilizing purpose-built bioreactors, as summarized in Table 2. The application of strain in in vitro dynamic models has been correlated to increases in myoblast proliferation, arrangement, and maturation. 49,53 –56 Myoblasts cultured on Bioflex plate systems with a radial cyclic strain of 10% or 17% resulted in increased myoblast proliferation and also a subsequent decrease in both myoblast differentiation and alignment. 57,58 The introduction of uniaxial cyclic strain of 10% supported an increase in myotube formation parallel to the direction of strain while suppressing cell motility and proliferation. 59 It is not clear whether these varying results are a consequence of the direction of strain (radial vs. uniaxial) or of the strain regimen itself, and more studies are necessary to understand the differences between direction and regimen on myofiber formation and alignment.
Effect of mechanical strain on fiber alignment in skeletal muscle tissue constructs
The utilization of dynamic uniaxial strain in 3D skeletal muscle constructs induces similar effects as observed in 2D. 59 A cyclic strain regimen of 10% applied three times per minute every hour supported significantly more aligned, dense myofiber maturation in human muscle precursor cell–seeded decellularized extracellular matrix (dECM) scaffolds than scaffolds cultured under static conditions. 49 Separately, when tissues were constitutively held at 3% strain except for 6-h periods each day where they were stretched to 10% strain, significant increases in myofiber alignment were observed after 9 days in culture, as well as upregulation of myoblast determination protein 1 (MyoD) and troponin T1 expression, compared against unstrained controls. 53 There is currently no standardized strain threshold for culturing skeletal muscle constructs. Constructs cultured at strains as low as 3% have shown improved myofiber alignment and maturation when compared to un-restrained controls. 53 However, the application of 10% strain is most common as it mimics strain experienced during embryonic development of the musculoskeletal system. 49,59 These findings suggest that the absolute strain value may be more important than strain rate, although this has not been thoroughly investigated in the literature and requires further exploration. It is important to note that in addition to tension, topographical cues of the substrate are also sufficient to induce cell alignment but not necessarily myoblast maturation. Electrospun fibrin microfibers have been shown to support similar amounts of highly aligned growth of C2C12 myoblasts with and without 10% cyclic strain for 5–7 days. 50 Despite no changes in alignment, myotube diameter and the expression of myosin heavy chain (MyHC) were significantly upregulated when exposed to the strain regimen, demonstrating that dynamic strain enhanced myofiber development. 50 In support of this observation, murine myoblasts exhibited an eightfold increase in MyHC expression when seeded on electrospun membranes that were uniaxially strained cyclically, followed by 5 days of static culture, as compared with constructs cultured completely in static conditions. 54 Taken together, these studies demonstrate that strain-induced tension is beneficial for improving myofiber maturation in addition to facilitating myofiber alignment, although there have been several examples of synergistic signaling between the application of both tension and matrix topography. It should be noted that absolute strain values beyond 20% appear to negatively impact myoblast behavior and are associated with reduced cell proliferation and even cell death. 51 Interestingly, there is evidence that the incorporation of physical therapy (i.e., strain in vivo) after VML impacts collagen and fibrotic tissue reorganization in vivo. Mice subjected to a VML injury in the tibialis anterior (TA) muscle exhibited significantly reduced collagen deposition when provided with unlimited access to a running wheel as compared with sedentary controls. 60 Rehabilitative practices such as running wheel access have been increasingly included as part of the treatment paradigm for VML repair and have shown modest increases in force production or improvements in overall gait. 61 –63 The exact mechanisms for these increases are unclear; however, there is some evidence that improvements can be attributed to exercise-induced inflammation, 60 highlighting the importance strain plays in muscle reorganization and also the need to generate more complex in vitro models of skeletal muscle tissue to understand complex interactions with other cell types, such as macrophages or other inflammatory cells. Therefore, more investigations regarding the impact of the absolute value strain values, as well as strain rate, are necessary to fully understand the potential for tension to be used to improve myofiber alignment and overall maturation within tissue constructs.
Induction of myofiber alignment with aligned scaffold architecture
Aligned topography or anisotropic scaffolds are another method to induce myofiber alignment, as briefly discussed in the previous section. As summarized in Table 3 and Figure 5, these designs function by recapitulating the structure and/or composition of native skeletal muscle ECM by culturing cells on a biomaterial that encourages cell growth along the surface or within the bulk of a material. 73
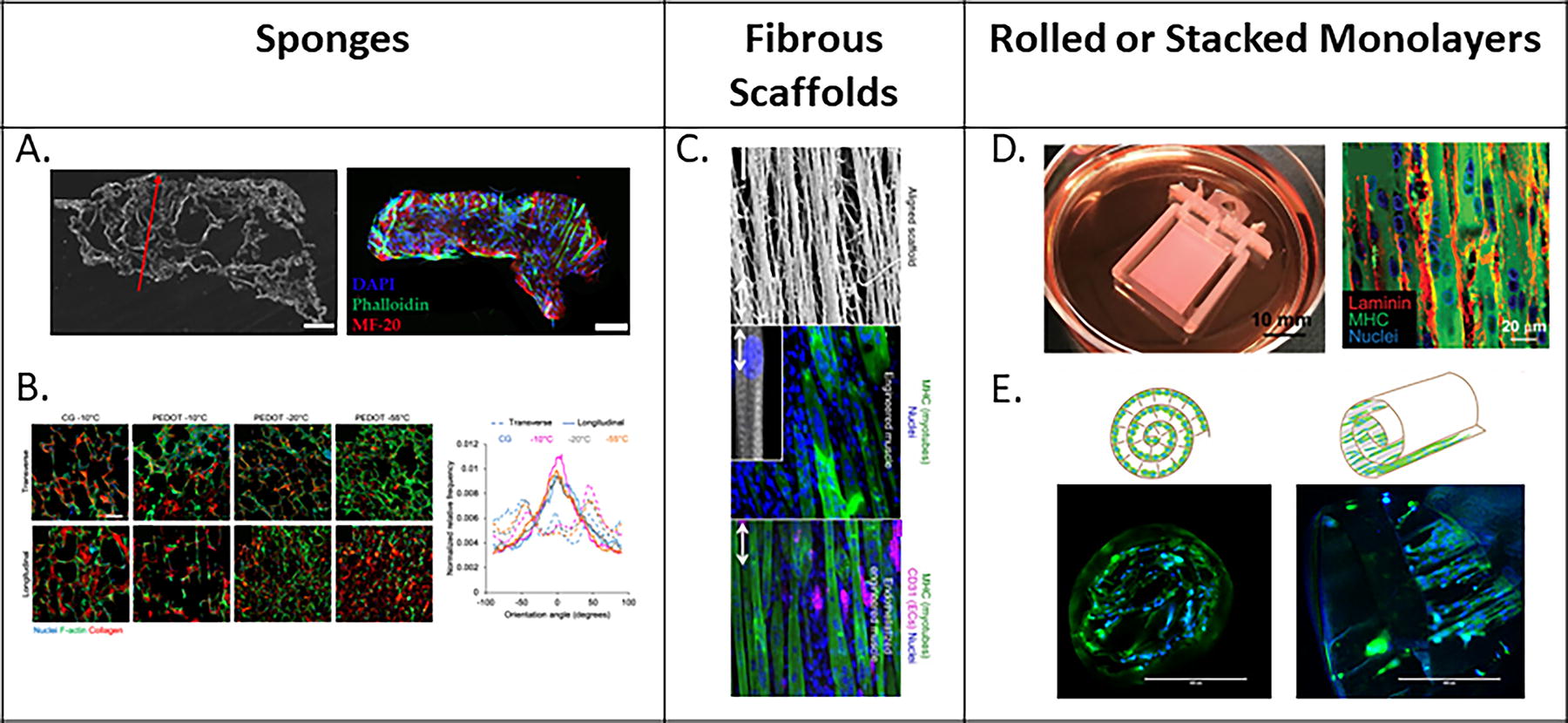
Prealigned methods of inducing myofiber alignment. Myofiber alignment facilitated by
Methods of inducing myofiber alignment with aligned scaffold geometries
hMPC, human muscle progenitor cells; PCL, poly-ε-caprolactone; PLGA, poly(lactic-co-glycolic acid); SCs, satellite cells; MyHC, myosin heavy chain; MyoD, myoblast determination protein 1.
In 2D, aligned or anisotropic skeletal muscle tissue models generally consist of substrates where grooves have been patterned via photolithography or soft lithography. Materials that have been utilized to create these microgrooved surfaces include methacrylated gelatin, glass, PDMS, polyacrylamide, fibrin, collagen, among others. 64,65,70,74 –76 Substrates may also be treated by coating with varying proteins in order to better mimic ECM composition and to improve cell attachment. 73 Both micropatterned geometry and size impact cell morphology, migration, and differentiation. By modifying the surface geometry of PLGA membranes, Piscioneri et al. demonstrated that primary human myoblasts exhibited an elongated morphology and a ∼65% fusion index in micropatterned channels with a width of 30 µm and height of 40 µm as compared with surfaces with a pillar and pit-shaped morphology. 68,75 Similar responses to environmental shape have also been observed at the single cell level. 66,68,75 The phenomenon of myofibers aligning on straited environments is conserved across a variety of studies regardless of the substrate material. 64,74,77 While grooves are capable of inducing myofiber alignment, groove depth and width may impact cell morphology. Studies exploring the effect of groove size on myofiber maturation determined that narrow grooves (0.4 µm and 2 µm) resulted in a significant reduction in both myotube maturation (density and diameter) and gene expression (alpha-actinin 2 and MyHC). These results suggest that controlling for alignment and shape is not sufficient to control myofiber development but that the size of topographical features must also be considered. In summary, these data demonstrate that skeletal muscle myoblasts are highly sensitive to surface mechanical and topographical cues.
Strategies utilizing anisotropic architectures to facilitate myofiber alignment have been translated into 3D tissues with aligned architectures (Fig. 5D, E). Several groups have taken monolayers from 2D striated surfaces and then either stacked or rolled these layers to create 3D tissues. 66,68,75 Takashi et al. demonstrated that muscle constructs synthesized from stacking six layers of aligned, differentiated human skeletal muscle myoblasts generated significantly larger twitch and tetanus forces than both single-layered, aligned tissue constructs, as well as multilayered, nonaligned tissue constructs. 66 Beyond stacking aligned 2D monolayers, there are a variety of strategies to impart anisotropy into 3D biomaterials. One such approach is to fabricate scaffolds with channels/pores, the aim of which is to create aligned negative spaces for cells to grow within (Fig. 5A, B). A common methodology of creating aligned, porous sponges involves the development of pores via controlled freezing and lyophilization of collagen sponges. 71,72,78 Porous scaffolds can also be synthesized via porogen leaching, in which a material is polymerized around a sacrificial component that is later dissolved or removed, thus leaving empty pores within the scaffold. 67,79,80 Attempts to utilize these scaffolds have been positive, enhancing both myofiber alignment and maturation in vitro, and have resulted in recovery of contractile force after VML injury in vivo. 11,71,78,81 While these materials are sufficiently robust to survive implantation, they are limited by insufficient myoblast migration into the scaffold and are not currently capable of inducing complete force recovery. 42
Utilizing biomaterials composed of discrete fibers provides a physical network for the cells to grow and align along (Fig. 5C). Unlike channel-based models, cells are encouraged to grow and incorporate between and within fibers rather than be constrained by the negative space between fibers. Electrospinning is the most common methodology to create fibers, although extrusion and shear-based methodologies are utilized as well. 69,82 Methods for inducing alignment from electrospinning include electrospinning along two-pin collectors, rotating drums, or rotating disc collectors as well as other collector designs. 73 Common materials utilized in the creation of fibrous scaffolds, as well as sponges or fibers, include fibrin, collagen, gelatin, polyester urethane, alginate, and more. 65,70,71,83 Contractile, aligned skeletal muscle tissues have been developed via the seeding of myogenic cells onto aligned, fibrous constructs. Both electrospun fibrin scaffolds and aligned, extruded nanofibrillar collagen scaffold strips seeded with C2C12s were capable of generating mature, highly aligned, contractile muscle constructs that exhibited contractile forces greater than fibrous scaffolds with random orientation. 69,70 These materials can generate aligned skeletal muscle tissues; however, layering of the fibers on top of each other results in lower porosity and higher tortuosity of the pore structures, which may make it difficult for cells to migrate through the entire material. Recent efforts to electrospin fibrous meshes with cells, as compared with seeding cells into fibrous meshes after fabrication, may allow for cells to permeate the entire tissue and thus produce denser, larger skeletal muscle constructs. 65 Bundles of wet extruded fibrin microthreads can also facilitate migration of myoblasts since these materials are discrete fibers without a bulk scaffold. 84 Furthermore, the cylindrical architecture of these fibers has been shown to direct myoblast alignment along the length of individual fibers, which can support functional recovery after VML injury. 85,86 Attempts to blend techniques, such as combining lyophilization or inducing pore formation within aligned fibrous scaffolds, may improve cell migration and thus myofiber formation by providing empty spaces for cells to migrate into while still maintaining structural, elongated fibers to provide architectural cues for cell growth. Combining aligned scaffold architectures with mechanical conditioning, such as the application of cyclic strain, has also been associated with a higher degree of myofiber maturation and development. 49,50 Continued investigation in combining and mimicking both architectural cues as well as mechanical cues in native skeletal muscle tissue would be an exciting area of growth for skeletal muscle tissue engineering.
Magnetically induced myofiber alignment
Magnetic fields have also been used to induce alignment in skeletal muscle constructs. These methods generally fall within two categories: directly binding magnetically reactive particles to cells within a material to induce alignment by moving cells in the direction of the applied field and the incorporation of magnetically reactive materials into a scaffold backbone to produce aligned channels or fibers within the material while in the presence of a magnetic field.
Demri et al. were able to create aligned cell chains 10–100 μm in length by exposing magnetically labeled C2C12s in collagen hydrogels to a magnetic field. 87 The amount of magnetic labeling (facilitated by iron content) directly impacted the degree of cell alignment, with increased iron content resulting in longer cell chain lengths. Myoblasts did not remain aligned when the magnets were removed; in fact, they rapidly lost any aligned organization implying that continuous exposure to magnetic fields is required to maintain alignment. This study did not examine how removing the magnets after the generation of organized myofibers would impact tissue alignment, making it unclear whether these methods can be utilized without a persistent magnetic field. Despite this limitation, this study demonstrated that myofibers were able to be strained and relaxed under the presence of the magnetic field, exhibiting a novel mechanism for inducing tensile strain on engineered skeletal muscle tissues. 87 Similar morphological manipulations were also observed by Ito et al., in which C2C12s labeled with magnetite cationic liposomes were organized to form string-like assemblies of cells when cultured under a magnetic field. Myofiber alignment was lost upon differentiation as a consequence of cells fusing and the generated tissue sheet shrinking, suggesting further work is necessary to advance this technology. 88 While the use of magnetism remains an exciting and new area of research to induce myoblast alignment, a limitation of this approach is that current methods suggest that the magnetic field must be continuously applied to facilitate alignment. This requirement extends to both the magnet and the cell-associated magnetic particles, and considerations will need to be made for changes in the magnetic particle concentration as a consequence of cell metabolism over time. Further studies are needed to determine such temporal and metabolic considerations, focusing on whether myofiber alignment post fusion of the myoblasts in culture will be maintained upon removal of the magnetic field. While this method provides a novel mechanism for organizing cellular structures, it may benefit from combining with tension or aligned scaffold methodologies.
3D printing muscle constructs
Bioprinting is a technology that involves the direct application of bioinks in precise geometries dictated by a computer-aided design rendering to generate 3D constructs. These systems typically create tissues using a layer-by-layer approach, thereby allowing for complex architectures to be realized with multiple cells or bioinks within the same construct. 89 As summarized in Table 4, bioprinting has been increasingly utilized for skeletal muscle tissue engineering as this technology allows for precise control of tissue architecture. This precise control has facilitated the development of aligned, multilayered tissues that resemble the native skeletal muscle tissue architecture. 52,90 –92 Materials utilized in bioink formulations include decellularized ECM, alginate, gelatin, fibrinogen, PCL, and collagen, among others, and the reader is referred to the following reviews for a more comprehensive overview of this topic. 89,96 The most common mechanism used for the bioprinting of engineered skeletal muscle tissue is extrusion printing, by which a hydrogel is extruded from a nozzle and rapidly polymerized either enzymatically, thermally, ionically, or chemically. 96 There are three general methodologies utilized in extrusion printing; printing in a dissolvable support bath, printing with a sacrificial scaffold, and printing only the material/bioink. 89,90,93 While all of these methods allow for the creation of aligned tissues, the usage of support baths or sacrificial scaffolds allows for the printing of more complex tissue geometries, such as the inclusion of negative spaces or channels to improve medium diffusion and cell infiltration throughout the tissue. 90,93,94 The inclusion of channels throughout 3D printed constructs has been shown to improve cell viability and maturation while maintaining myofiber alignment, suggesting that these techniques may allow for the easy creation of vasculature without disturbing myofiber arrangement within a construct. 90,94
Strategies to generate aligned 3D printed skeletal muscle tissue constructs
IGF-1, insulin-like growth factor 1; PCL, poly-ε-caprolactone; PEG, polyethylene glycol; hMSC, human mesenchymal stem cell; MyHC, myosin heavy chain.
A common approach to inducing alignment has been to print cell-seeded aligned architectures without any additional tension. 52,92,94 Tissue alignment and maturation have been controlled through bioink optimization to develop materials with mechanical properties similar to other scaffolds for muscle tissue engineering. This includes the utilization of fast relaxation viscoelastic bioinks, bioinks with porosity, and bioinks that can be stacked or rolled to create more complex 3D shapes. 92,94,97 The extrusion of cells within the bioink allows for the creation of cell-laden tissues with controlled internal alignment as well as controlled bulk tissue structure, resulting in the generation of aligned skeletal muscle fibers. However, multiple models have also begun to include anchoring supports during or post printing. Examples include extruding polyethylene glycol (PEG)-fibrin hydrogels around rotating C-shaped supports and loading muscle-derived ECM hydrogels into polycaprolactone anchoring molds in order to apply tension to the 3D-printed construct. Extrusion printing of PEG-fibrin hydrogels loaded with muscle-derived human mesenchymal stem cells around a rotating C-shaped support produced mature, spontaneously contracting engineered skeletal muscle constructs. 91 This model resulted in denser fiber formation as compared with a similar structure synthesized by bulk casting of PEG-fibrin, suggesting that the use of anchoring supports along with 3D printing specific architectures is capable of generating highly aligned tissues that also exhibit improved myofiber maturation. 91 Interestingly, this approach combines tension-based methodologies along with the advantages of 3D printing techniques to generate aligned scaffolds with a high cell density, and further studies should exploit these approaches, particularly for implantation uses.
Implantation of aligned constructs
While aligned skeletal muscle tissues can serve as platforms for drug testing, another common application of these constructs is to serve as potential therapeutics for VML injuries (Fig. 6). Unlike autologous muscle flap transfer, skeletal muscle constructs are not dependent on a patient having intact, transplantable skeletal muscle tissue and instead can be generated from less invasive biopsies of muscle tissue or derived from patient-specific induced pluripotent stem cells. 8 The use of minced muscle in a rat TA VML injury, where healthy muscle was minced and replaced into the surgical site, resulted not only in cell migration and de novo myofiber formation but also in limited, aligned myofiber recovery. Work by Ward et al., which implanted minced muscle grafts into the TA of male Lewis rats, exhibited some graft survival and improved contractile force recovery as compared with untreated controls; however, this approach was limited by a lack of aligned myofibers within the injury region (Fig. 6A). 98 This is further supported by a multitude of studies where the implantation of acellular, aligned scaffolds is associated with improved force recovery and tissue regeneration as compared with the implantation of unaligned, randomly oriented, or bulk-fabricated materials, again highlighting the need to induce organized regeneration to contribute to force production. 69,70,82
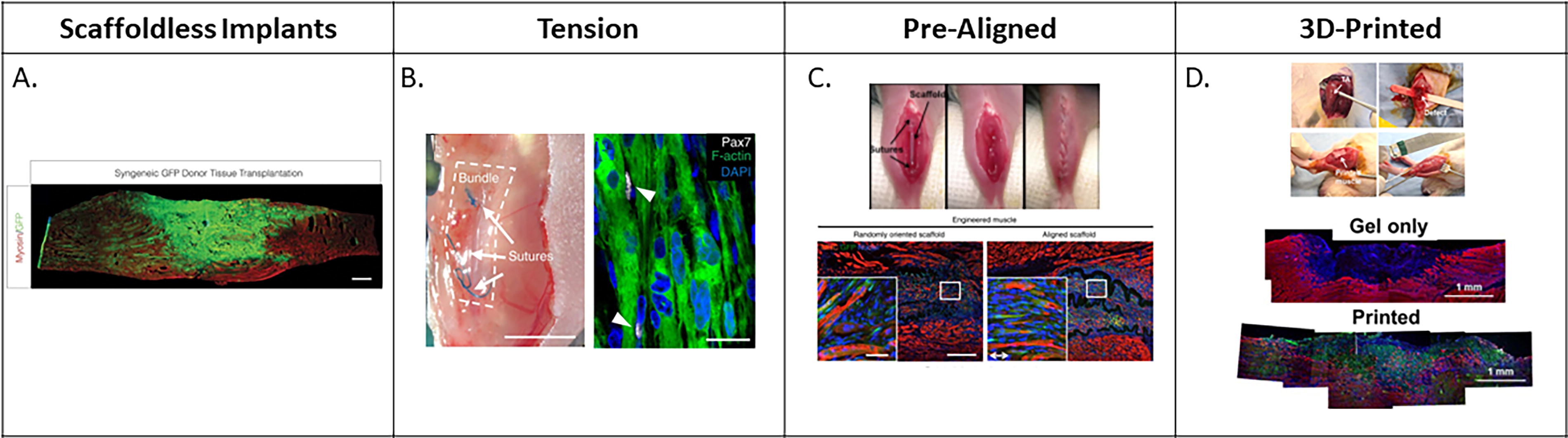
An overview of different tissue construct strategies that have been implanted in preclinical models of VML.
There are varying approaches to the implantation of cellular-based constructs for skeletal muscle tissue engineering. Recapitulating developmental cues to recruit myogenic progenitor cells to form new myofibers in situ is an attractive method also utilized by acellular approaches. The alternate approach is to implant mature myofibers that can remodel to engraft and immediately contribute to force production. Mechanical preconditioning of constructs for 1 week prior to implantation resulted in constructs that not only integrated into the injury site but also contributed to the mechanical recovery of functional output. 49 Separately, incubating myoblasts in differentiation medium on electrospun fibrin fibrous bundles for 9–12 days prior to implantation produced dense myofibers as well as elevated MyHC expression while also enhancing construct engraftment, providing evidence that in vitro tissue maturation prior to implantation facilitates and maintains myofiber alignment and maturation. 41,49,70 These data suggest that myofibers maintain maturation that was developed in vitro, and the constructs continued to mature after implantation to aid in force production. This is further supported by the successful integration of scaffold-free, cellular tissues into skeletal muscle. These constructs deposited new ECM and maintained myofiber alignment similar to the surrounding native tissue, suggesting that exposure to the native skeletal muscle environment encouraged further construct maturation and development as compared with in vitro constructs. 43 Taken together, cell-laden tissue constructs integrate with native skeletal muscle, and their survival and maturation are enhanced after implantation. Tissues aligned under tension (Fig. 6B), as well as prealigned (Fig. 6C) and 3D-printed tissues (Fig. 6D), all appear to integrate more effectively with native tissue as compared with unaligned controls. 47,69,90,98 While maturation improves after implantation, implant architecture and preconditioning techniques appear to be necessary to initially optimize myofiber alignment and implant integration in vivo.
Comparison between methodology of inducing alignment and wound-healing capabilities is also further complicated by differences in the animal model utilized, injury methodology, and implant methodology as outlined in Table 5. Inconsistencies between injury models, such as variations in the percent of muscle removed, 69,70 the specific muscle removed, 41,69 and how the muscle injury was induced 91,98 all have downstream effects on how a therapeutic might reintegrate with the remaining tissue. 10 Furthermore, while mouse and rat models are routinely used to provide insight into evaluating repair mechanisms for the treatment of VML injuries, their clinical relevance is somewhat limited by their relative size with respect to humans, particularly in terms of muscle mass. These VML injuries do not accurately recapitulate the clinical scenario, where defects are on the order of cubic inches rather than cubic millimeters in size, an approximate 15,000× increase. Therefore, there is also a need to develop larger animal models, such as the sheep, 99,100 to achieve clinically relevant results in preclinical trials before progressing to first-in-human trials. For more information, the reader is referred to the following review articles. 10,37 An additional consideration to the generation of accurate preclinical models is that many of the existing models solely consist of the removal muscle tissue. VML typically manifests from traumatic injuries that result in the creation of composite injuries, including the destruction of bone and skin tissues in addition to skeletal muscle, complicating the injury. Studies exploring more complex musculoskeletal injuries have demonstrated that combined attention to repairing bone and muscle simultaneously appears to improve overall recovery, 101 although more investigation is needed in this area. How an aligned construct is implanted may also further dictate the extent of tissue and force recovery. Techniques to implant tissue constructs vary from suturing directly into the remaining host muscle, 41,70 to securing the construct in the tissue by suturing the surrounding fascia in place. 69,91,98 Unfortunately, failure to properly secure implanted tissues may reduce force recovery, and implants that are sutured into the injury site may integrate more efficiently as compared with implants passively positioned into the injury site. A recent study implanted aligned constructs deliberately misaligned to the muscle and demonstrated that this had a significant impact on force recovery and tissue integration. 102 Broadly, aligned muscle constructs appear to integrate and improve force production in VML injuries more successfully than unaligned constructs. Factors such as material selection, tissue maturation prior to implantation, and surgical technique have been shown to impact implant success, integration, and functional recovery. However, further studies are needed in order to optimize and further the creation of implantable, aligned muscle constructs to serve as therapeutics for VML.
Effect of construct alignment on scaffold integration post implantation
EDL, extensor digitorum longus; EHL, extensor hallucis longus; TA, tibialis anterior.
SUMMARY
It is becoming increasingly clear that myofiber alignment is essential not only to repair VML injuries, but also to generate mature muscle constructs in vitro. Attempts to create engineered skeletal muscle constructs with precise geometries have shown that aligned tissues exhibit morphology similar to native, striated skeletal muscle and have a higher degree of success after implantation as compared with randomly oriented tissues. 14,65,69,70,82,91,94 Myoblast alignment can be directed with mechanical cues (e.g., tension and cyclic strain, magnetic alignment), as well as architectural cues (e.g., aligned scaffold architecture, 3D-printing), and these cues can be combined to further enhance alignment and maturation. The most successful strategies for skeletal muscle tissue engineering include both sets of cues, to exclude architectural considerations may reduce myofiber maturity and function, 46,47 as these have been demonstrated to provide 3D structures for cells to travel, mature, and elongate. 49,50 While application of tension greatly enhances myofiber maturity, it cannot always allow for the customizability and control of myofiber arrangement seen with architectural cues. 18,44,46 –48 Therefore, combinatorial approaches involving both architectural and mechanical stimuli have shown promising results in restoring force production and integrating with host tissue in vivo and may provide optimal conditions necessary for tissue development to facilitate tissue repair. 41,42,50,103 Since alignment improves tissue integration with native skeletal muscle tissue and recovered force production, recovery may be further improved with better surgical practices that more efficiently connect aligned portions of engineered constructs to remaining native skeletal muscle. As more strategies utilize 3D printing approaches for muscle regeneration, another approach can be to directly print materials into injury sites, as this is another promising technique to both facilitate alignment and fill the wound site. 104,105 In order to translate skeletal muscle constructs to the clinic, these tissues must be biocompatible and capable of integrating with native tissue without inducing a detrimental immune response or suffer from necrosis. Other limitations include the production timeline of constructs, shipping requirements, and construct shelf-life. Currently, biomimetic solutions that are in preclinical or clinical testing are acellular and do not require complicated procedures prior to implantation. These devices are typically shipped dehydrated or lyophilized to increase shelf-life and require a facile rehydration step prior to deployment. While these therapeutics have established solutions for many of the major barriers toward clinical translation, it is becoming increasingly clear that cell migration into an acellular scaffold is not sufficient for the complete regeneration of excessively large defects within skeletal muscle. Cellular constructs, on the other hand, are generally thought to be more capable of encouraging more regeneration than acellular scaffolds, but logistics behind the major barriers continue to exist. 106 Efficacy needs to continue to be established regarding the capacity of cellular constructs to repair VML. The translation of these techniques from preclinical research to clinical settings will require the creation of infrastructure that allows for the collection, expansion, and culturing of patient-specific cells to construct these implants, either through collection of biopsy tissues or through the generation of iPSC lines. The procedures to develop these constructs will need to be streamlined to minimize the delay between time of injury and treatment, although current approaches toward biomaterial-mediated VML repair in human patients similarly occurs well after injury. 107 These constructs will need to be mechanically robust to not require further specialization from surgeons to ensure widespread adoption over the autologous tissue transplant procedure. Ultimately, the successful creation of tissue-engineered skeletal muscle constructs is dependent on the recapitulation of tissue structure. Proper tissue integration as well as contractile function requires that designed tissue comprises aligned, organized myofibers. While much work has been done to develop these constructs, future approaches will need to consider the addition of cell types, such as vasculature and nerves, in order to better recapitulate muscle structure, composition, and components to maximize force production and, therefore, improve patient quality of life.
TAKE-HOME MESSAGES
Tissue-engineered skeletal muscle constructs may be used as effective, alternative therapeutics for VML that are not limited by the availability of intact native skeletal muscle tissue. Myofiber alignment can be controlled by external mechanical forces as well as by architectural cues within a scaffold. Muscle maturation within constructs is improved with the addition of external stimuli that recapitulates the native skeletal muscle tissue environment. Scaffolds that facilitate myofiber alignment integrate with native tissue more effectively than scaffolds with random myofiber arrangement. Animal models need to be carefully considered and developed to ensure accurate recapitulation of the clinical reality of VML injuries.
Footnotes
ACKNOWLEDGMENTS AND FUNDING SOURCES
The authors would like to thank Marlin Abdelmasih for her contributions to this work. This research was funded in part by the National Institutes of Health (KL2-TR003018 & R21-AR079708) and by startup funds provided by New Jersey Institute of Technology.
AUTHORS’ CONTRIBUTIONS
S.T.S. and J.M.G. designed and formatted the article. S.T.S., N.H., and Y.P. collected data in the form of literature reviews and summaries of published manuscripts. S.T.S., N.H., and Y.P. wrote the initial draft of the article. S.T.S. and J.M.G. revised and finalized the article. All authors approved the final version of the article.
AUTHOR DISCLOSURE AND GHOSTWRITING
The content of this article was written by the authors listed. All figures were adapted from open-source publications and are properly attributed by referencing in accordance with STM guidelines. No ghostwriters were used to write this article.
ABOUT THE AUTHORS