
Abstract
Abstract
We studied humoral long-term adaptive viral neutralization responses in zebrafish (Danio rerio), an increasingly useful vertebrate model for viral diseases actually limited by the absence of standardized anti-zebrafish immunoglobulin M (IgM) antibodies. We established an alternative method, similar to those used in other fish, to achieve a first estimation of zebrafish anti-viral antibody-like responses. We used the viral hemorrhagic septicemia virus (VHSV) model because, although protection after this non-natural infection was demonstrated in cold-acclimatized zebrafish, little is known about their induced anti-VHSV antibody-like responses. Therefore, we first optimized a micro-neutralization method based on immunostaining VHSV-infected fish cell monolayers to detect zebrafish neutralizing activity in plasma samples in one day. We then used the method to measure the specific anti-VHSV neutralization in plasma obtained from individual zebrafish under various VHSV challenges or immunization protocols. The neutralizing activity was inhibited by protein A-sepharose and rabbit anti-zebrafish IgM antibodies, suggesting the implication of IgM zebrafish antibodies in such responses. To our knowledge, this is the first report to demonstrate detectable and significant VHSV neutralization titers in zebrafish surviving VHSV infections. This micro-method might be useful, not only for the follow-up of infection/vaccine development in the zebrafish/VHSV model in particular, but also for similar work involving other in vitro neutralizable zebrafish pathogens. This technique might also further the development of alternative ELISA assay methods to measure specific immunoglobulins in zebrafish.
Introduction
In this context, susceptibility conditions of zebrafish to infections caused by several rhabdoviruses naturally infecting other fish species, 4 such as viral hemorrhagic septicemia virus (VHSV),2,3 infectious hematopoietic necrosis virus (IHNV), 5 snake-head rhabdovirus (SHRV), 6 and spring viremia of carp virus (SVCV), 7 have been described. Salmonids or cyprinids surviving infection with rhabdoviruses such as VHSV,8,9 IHNV,10,11 or SVCV, 12 showed long-term adaptative immune responses that included the production of in vitro neutralizing antibodies (NAbs) in 50% of the survivors, and antibodies (Abs) detectable by enzyme-linked immunosorbent assay (ELISA) in an even higher percentage of these fish. 13 Furthermore, most survivor fish show enhanced resistance to disease upon subsequent exposure to rhabdoviruses. However, despite interest in zebrafish as a disease/immunological model, few studies have addressed the specific long-term Ab responses of these fish to the above-mentioned rhabdoviruses.
Using Western blot analysis and polyclonal Abs (PAbs), others tried to estimate zebrafish immunoglobulin M (IgM) in order to study the development of their immune system. 14 The isolated heavy chain used as an antigen to produce rabbit PAbs confirmed the recognition of their corresponding 84–86 KDa band. 14 However, no ELISA or immunohistochemical assays were reported. Our own previous attempts to raise PAbs to zebrafish IgM either using peptides derived from the CH4 domain or using commercially available anti-carp IgM monoclonal Abs (MAbs) selected for their apparent cross-reactivity with zebrafish IgM failed to estimate viral-specific Abs by ELISA. More recently, the use of MAbs against the heavy chain of zebrafish IgM has been described, 15 but no detailed characterization have been reported to date, nor are these MAbs available for use in ELISAs.
Current methods to estimate VHSV protein G-specific Abs in salmonids include in vitro neutralization assays11,16 or binding to VHSV-captured or recombinant G solid-phase ELISAs;17,18 however, none of these techniques have been reported yet for zebrafish Abs. In particular, in vitro neutralization tests, which are commonly performed in the presence of complement, have been widely used to estimate VHSV protein G-specific Abs in trout,8,11,19 but not in zebrafish. In addition to being highly specific and sensitive, the detection of in vitro neutralizing fish Abs generally shows some correlation with protection against VHSV challenge.20,22
Therefore, to estimate neutralizing-like activity in zebrafish, we adapted a micro-neutralization method for detection of VHSV based on an immunostaining procedure described for trout anti-VHSV Abs. 16 The modified method increases speed (1 day), simplicity, and minimizes the use of reagents and handling of nonsterile cell cultures compared to other assays (5 days, 4–10 times more reagents, and need of strict sterility). By using the micro-neutralization method, VHSV-infected foci of 5–12 infected fish cells in a 96-well monolayer could be easily detected and counted under an inverted microscope ∼24 h after infection of cell monolayers with a high titer anti-VHSV N protein monoclonal Ab (MAb). This micro-neutralization method is more practical for use in zebrafish than other neutralization assays, such as those using agarose or methylcellulose plaques, mainly because it requires only a small sample of blood from individual fish. Following this approach, zebrafish that survived infection with VHSV or were immunized against the VHSV protein or gene G developed detectable neutralizing activity in blood plasma while showing partial resistance to VHSV challenge. The neutralization was most probably caused by specific IgM zebrafish antibodies as suggested by their inhibition by protein A-sepharose and rabbit anti-zebrafish IgM antibodies.
Materials and Methods
VHSV virus and cell culture
VHSV 07.71 isolated in France from rainbow trout Onchorynchus mykiss 22 was grown in the Epithelioma papulosum cyprini (EPC) cells from the fathead minnow fish Pimephales promelas obtained from the ATCC collection (CRL-2872). Cells were grown in 25-cm2 flasks at 28°C in RPMI Dutch modified cell culture medium buffered with 20 mM HEPES (Flow) and supplemented with 10% fetal calf serum (FCS), 1 mM pyruvate, 2 mM glutamine, 50 μg/mL gentamicin, and 2.5 μg/mL fungizone. To prepare VHSV for in vivo challenges, the cell culture media were the same as above, except for the inclusion of 2% FCS and 10 mM Tris pH 8.0. To assay for VHSV infectivity, VHSV was purified by pelleting VHSV-infected supernatants by ultracentrifugation at 60,000 g for 60 min at 4°C.
Construction of recombinant glycoprotein G and its frg11 from VHSV
Purified recombinant glycoprotein G and frg11 from VHSV were used to immunize zebrafish. The DNA sequence 24 corresponding to the glycoprotein G gene of VHSV 07.71, was used for the cloning experiments (Gene Bank accession number X59148).
For insect larval expression of the glycoprotein G, the corresponding G gene was cloned into the appropriate plasmids by substituting their hydrophobic signal peptide for that of honey bee melittin in the absence of their transmembrane region (amino acid 21–465, G21–465), as described before in detail. 25 Briefly, G21-465 was subcloned into the carboxy-terminal polyhistidine tail-containing pFastMelB2 vector coding for the melittin signal peptide (Invitrogen). 26 Constructs were transformed in DH10 Bac™ competent E. coli cells using the Bac-to-Bac® baculovirus expression system (Invitrogen) and transfected into Spodoptera frugiperda (Sf21) cells. 27 To express the glycoprotein G21-465, Trichoplusia ni (cabbage looper) larvae were injected with the corresponding recombinant baculoviruses. 27 About 72 h later, ∼15 g of infected larvae were homogenized in 6 M guanidinium chloride, 1 M sodium chloride, 25 mM imidazole, protease inhibitor cocktail (Complete, Roche), and 40 mM phosphate buffer pH 7.8 as described previously. 25 After removing the pellet, the extracts were purified as described below.
In addition, the corresponding DNA sequences of frg11 of VHSV protein G (amino acids 56–110) flanked by the appropriate restriction enzyme sequences were chemically synthesized (BioS & T Inc, Montreal, Canada) and subcloned into the E. coli pRSETa expression plasmid (Invitrogen, San Diego, CA), as described previously in detail. 18 To achieve expression of the cloned frg11 in bacteria, we transformed E. coli BL21 DE3 cells with the pRSETa-frg11 plasmids. These cells were then grown in TB medium with ampicillin and induced with IPTG, as described previously. 18
Given that, in addition to their protein G sequence, the recombinant proteins contained 33 amino acids (4 KDa) with polyhistidines (polyHis) in their amino terminal tails, their extracts were applied to a Ni affinity column (ProBond™, Invitrogen) previously equilibrated with binding buffer (6 M guanidinium chloride, 1 M NaCl, 25 mM imidazole, and 40 mM NaH2PO4, pH 7.8). The same buffer was passed through the column to wash unbound proteins. Elution buffer containing 150 mM imidazole was used to obtain the polyHis-containing G21–465 and frg11. Peak fractions, identified by absorbance at 280 nm, were pooled, adjusted to pH 4 to prevent precipitation, chromatographed through Sephadex G-100 in 20 mM sodium acetate pH 4, and extensively dialyzed against 20 mM sodium acetate pH 4. Protein concentrations were determined using the BCA method, and purity was confirmed by densitometry of Coomassie blue-stained protein bands separated on 10%–20% polyacrylamide gradient gels (PAGE, BioRad).
Production of rabbit antibodies against the CH4 constant region of the zebrafish IgM heavy chain
The 891–1223 nucleotide sequence corresponding to the CH4 domain of the constant region of the heavy chain of serum immunoglobulin M (IgM) from zebrafish (Gene Bank accession number AY643753) 28 was used to obtain the corresponding recombinant protein to raise rabbit antibodies (anti-IgM). Briefly, the CH4 sequence was flanked by BamHI and XhoI restriction enzyme sequences, chemically synthesized (BioS & T Inc) and subcloned into the pRSETa plasmid (Invitrogen). Recombinant CH4 expression and purification were performed as described above. 18 Rabbit antibodies (anti-CH4 IgM zebrafish) were raised by intramuscular injection by using Freund's protocols (GenSpring, Hong-Kong, China). The titer of the rabbit antiserum, estimated by indirect ELISA on solid-phase coated with recombinant CH4, was a dilution of 1/100,000 to obtain background values. Confirmation of zebrafish IgM recognition by anti-CH4 IgM zebrafish rabbit antibodies was demonstrated by Western blot analysis on SDS polyacrylamide gels in buffer containing ß-mercaptoethanol, as described before. 18
Western blot analysis to determine the presence of the heavy chain of zebrafish IgM in zebrafish plasma
Western blot analysis was used to detect the presence of the heavy chain of zebrafish in plasma after their treatment with protein A-sepharose beads and anti-IgM zebrafish rabbit antibodies. To perform the immunoblotting, ∼60 μg of treated zebrafish plasma protein per lane were loaded in 10%–20% SDS-polyacrylamide gels, separated, and then transferred to nitrocellulose membranes (Schleicher & Schuell). These membranes were first treated overnight with a blocking solution of phosphate buffer saline 0.05% Tween 20 (PBS-T) with 4% skin milk. Then the membranes were incubated for 1 h at room temperature with rabbit anti-CH4 IgM zebrafish. After washing with PBS-T, lanes were incubated with 200-fold diluted anti-rabbit horseradish peroxidase-conjugated IgG (Sigma). Finally, the bands were visualized by chemiluminescence with an ECL kit (Amersham).
Plasmid immunization of zebrafish
The pT2MCV1.4 and pT2MCV1.4-G plasmids, previously described in detail, 29 were used for oral delivery. The expression cassette of the latter is flanked by both left (L) and right (R) inverted terminal repeats (ITR or T2) from the sleeping beauty (SB) transposon. 30 The expression cassette contains the MCV1.4 promoter, an immediate early cytomegalovirus (IE-CMV) promoter of 994 bp, which includes a synthetic intron 31 and a SV40 terminator. This cassette was selected as one of the best G expressing plasmids from a previous screening of various fish promoters. 29 The pT2MCV1.4G plasmid coded for the protein G of VHSV-0771. The G gene of VHSV was chosen because the VHSV is an important viral disease in European aquaculture and because it codes for a membrane glycoprotein that is difficult to express as a conformationally intact trimer. 25
Zebrafish maintenance
Adult zebrafish weighing 600–800 mg (∼4 cm in length) were obtained from a local pet shop (Aquarium Madrid, Madrid, Spain). They were maintained at 20°C in 30-liter aquaria (1 g CaCl2, 1 g NaHCO3, and 0.5 g Instant Ocean sea salts, resulting in a water of 200–300 μS, pH 7.8–8.2) provided with biological filters, and fed with a commercial fish feed. We chose 20°C because the protein G conformation of VHSV (the antigen to be expressed in immunization experiments) is conformationally unstable at higher temperatures. 32 The use of tamponed water with high pH facilitated the reproducibility of the pH-sensitive infection with VHSV. 33 All the experimental protocols were performed with the approval of the Ethics Committee at INIA.
Zebrafish immunization
For the immunization by injection, 50 adult zebrafish were intraperitoneally (ip) injected with 10 μg each of G21-465 and frg11 in 5 μL of saline buffer mixed with 5 μL of complete Freund's adjuvant. The injections, diluted 1:1 with incomplete Freund`s adjuvant, were repeated every month for 3 months.
For the oral immunization, each plasmid (see legends in Figs. 3 and 4) was complexed with alginate following the procedure described previously for infectious pancreatic necrosis virus (IPNV).34,35 Briefly, 250 μL of 3% (w/v) of sodium alginate were mixed with 100 μg of pT2MCV1.4-G in a total volume of 450 μL. The mixtures were stirred for 10 min in 4 mL glass tubes provided with a Teflon fly at maximal speed. Then 1.3 mL of paraffin oil containing 350 μL of Span 80 were added to each tube, and the mixture was emulsified for 30 min by stirring, as above. Microspheres were prepared by adding 240 μL of 0.15 M CaCl2 drop by drop to the emulsion and stirring for 30 min. Microspheres were then collected by centrifugation at 2000 g for 10 min, washed twice with 70% ethanol, and stored at 4°C until the next day. Immediately before fish vaccination, the microspheres were re-suspended with 100 μL of phosphate-buffered saline (PBS) and vortexed. Zebrafish were orally vaccinated with 10 μL of suspension of the vaccine microspheres each containing 10 μg of plasmid diluted in 10 μL of PBS. Vaccination was performed with an automatic pipette with a 20 μL tip, which was introduced into the mouth of each fish, supporting the tip end at the entrance of the esophagus.34,35
Zebrafish plasma harvest
Two months after immunization, the zebrafish were anesthetized prior to handling by immersion in 90 mg of tricaine-ethanesulfonate (MS-222, Sigma, Madrid, Spain) per liter of phosphate-buffered saline (PBS). They were decapitated, and their blood individually collected in Eppendorf tubes filled with 100 μL of sterilized 0.64 g sodium citrate, 0.15 g EDTA, and 0.9 g sodium chloride in 100 mL of distilled water (anticoagulant media). The individual blood samples were immediately centrifuged to obtain 1/20–1/30 diluted plasma. Protein content of plasma was between 1–3 mg of protein per mL as evaluated by nanodrop absorbance at 280 nm. Plasma was de-complemented by heating to 45°C for 30 min, and kept frozen at −20°C until used.
VHSV challenge to immunized zebrafish
For the VHSV challenge protocol, groups of 10 adult fish were moved from their aquaria to 2-liter mini-aquaria provided with biological filters at 14°C. After acclimation to this temperature for 7 days, the groups were infected by immersion in 2×106 focus-forming units (ffu) per mL of VHSV or with cell culture media (mock-infected) at 14°C for 2 h in 50-mL bottles filled with aquarium water. Infected and noninfected fish were then released to the 2-liter aquaria by extraction with a net, and mortality was recorded over 30 days at 14°C. Under these conditions, cumulative mortality after 30 days was between 80% and 100%. 3 Relative percent survival (RPS) was calculated by the formula [1 - (mortality in vaccinated fish/mortality in nonvaccinated fish)]×100, provided the cumulative percent mortality of the nonvaccinated control group was >60%. 36
To increase the yield of VHSV survivors, we developed an alternative protocol by first using intraperitoneal (ip) injection of each zebrafish with 106 ffu of VHSV in 10 μL of volume and then incubating the infected fish for 30 days at 18°C. This method reduced the mortalities ∼2–3-fold (Fig. 1). The survivors of ip injection were then maintained for ∼2 months at 24°–26°C and finally challenged with VHSV by following the protocol described above.
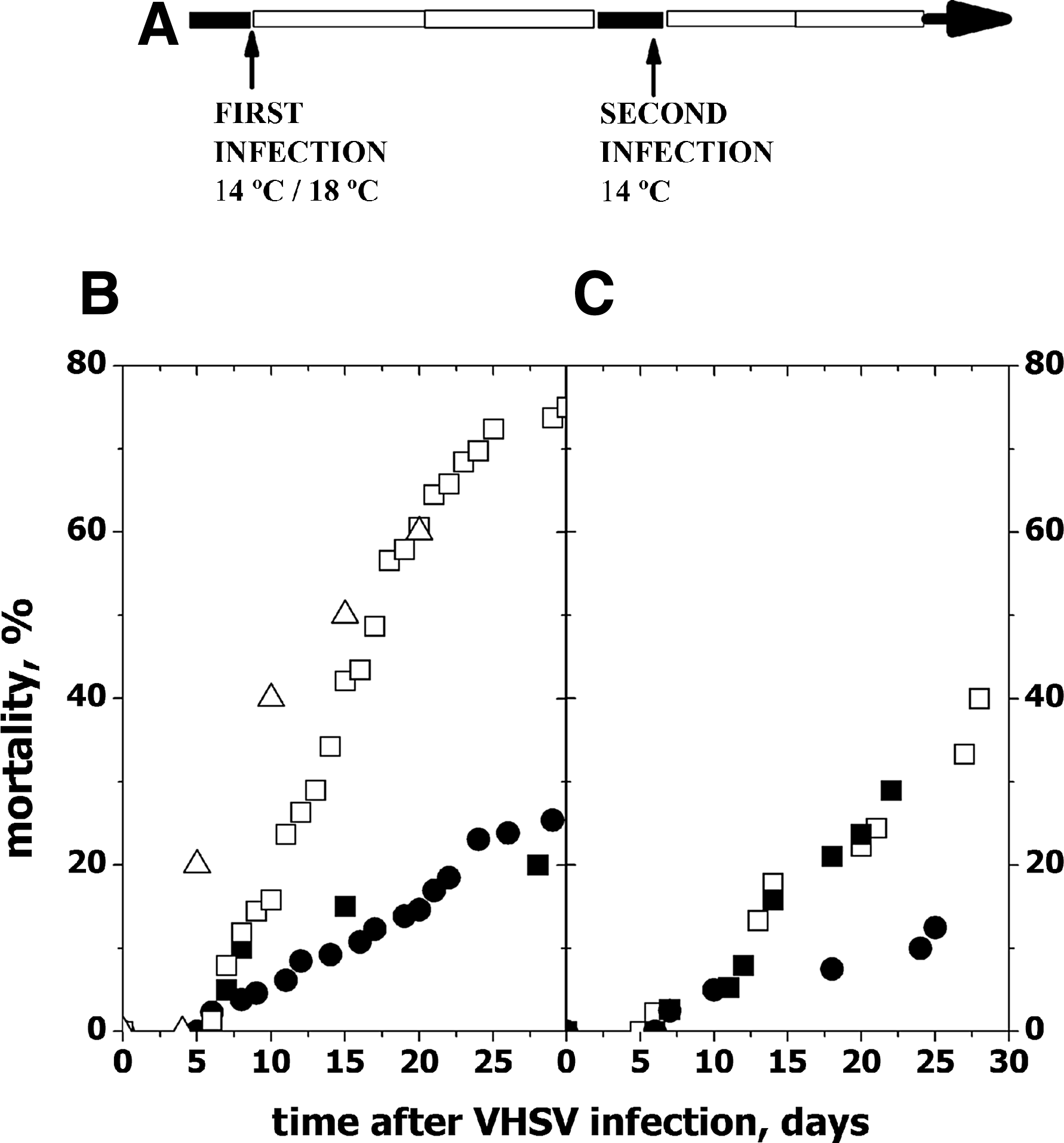
Time course of zebrafish mortalities after first and second VHSV infections.
Estimation of VHSV neutralizing activity by micro-neutralization
EPC cell monolayers in wells of 96-well plates coated with poly-D-lysine (Corning) were used for the assay. To infect the cell monolayers, they were incubated with 104 ffu per mL of VHSV purified by ultracentrifugation (purified VHSV) overnight at 14°C with serial dilutions of de-complement zebrafish plasma. To assay for zebrafish complement-dependent activity, 50 μg of fresh fish plasma per well was added to the virus/plasma mixtures as a source of complement during the overnight incubation. As a positive control for neutralization, a mix of anti-VHSV protein G monoclonal antibodies (MAbs) was used throughout the experiments, namely the C10+ I16+3F1A12 mix described before. 29 The following day, monolayers containing 100,000 EPC cells per well were infected with 100 μL of the virus/plasma mixtures. These mixtures were allowed to adsorb to the cell monolayers for 2 h at 14°C by means of continuous gentle agitation. The monolayers were then washed with cell culture media to remove nonadsorbed VHSV and carefully filled out with 100 μL/well of cell culture medium to prevent the removal of EPC cells. VHSV-infected EPC cell monolayers were incubated overnight at 14°C before being fixed during 10 min in cold methanol and dried. To detect the N protein of VHSV, MAb 2C9 37 1000-fold diluted in dilution buffer (130 mM NaCl, 2 mM KCl, 8 mM Na2HPO4, 1.4 mM KH2PO4, 0.24 mM merthiolate, 5 g Tween 20/L, and 50 mg phenol red/L, pH 6.8) was added to the wells (100 μL/well) and incubated for 1 h. After washing the cell monolayers with distilled water, 100 μL of peroxidase-labeled anti-mouse IgG (Nordic, Tilburg, The Netherlands) was added to each well, and the incubation continued for 30 min. After washing the cell monolayers 3 times by immersion in distilled water, 1 mg/mL per well of diaminobenzidine (DAB) (Sigma, Che, CO) in 50 μL of the appropriate buffer was added. The reaction was allowed to proceed until brown foci in the controls containing VHSV alone without zebrafish plasma 16 were detected by inverted microscopy. The neutralization was expressed as the reduction of the percentage of ffu as calculated by the formula, 100×ffu/number of noninfected cells. Titers were defined as the amount of zebrafish plasma protein required to cause a 50% reduction in the number of ffu obtained in wells infected with VHSV alone.
Inhibition of VHSV neutralization
Inhibition of VHSV neutralization was performed by several treatments of plasma from zebrafish surviving VHSV infections or from noninfected zebrafish before the neutralization assay overnight at 4°C. Treatments were performed with 50 μL of plasma and 50 μL of PBS, of PBS-washed protein A-sepharose beads (Pharmacia), of rabbit antiserum obtained against the recombinant CH4 domain of the heavy chain of zebrafish IgM (anti-IgM), or of protein A-sepharose beads and the subsequent supernatant with anti-IgM. Next day, the mixtures were centrifuged 10 min at 20,000 g and their supernatants assayed both for the presence of the heavy chain of IgM by Western blot using rabbit anti-CH4 IgM zebrafish and for VHSV by the micro-neutralization assay. The relative amounts of heavy chain (∼80 KDa) in Western blots were classified as: ++++, highly-saturated stained bands; ++, visible stained bands; and±slightly stained bands. The inhibition of neutralization was then calculated in percentages by the formula: 100×percentage of ffu obtained with treated plasma from survivor zebrafish/percentage of ffu obtained with treated plasma from noninfected zebrafish.
Results
To obtain zebrafish plasma with a high probability of containing anti-VHSV Abs, fish received intraperitoneal (ip) injections of VHSV recombinant protein G and frg11 (injected zebrafish) or were infected twice with VHSV (VHSV survivors). In the former case, the conditions for infection were optimized to produce the highest possible number of survivors to VHSV, since cold-acclimatized zebrafish infected-by-immersion at 14°C suffer high ∼75% mortalities, as shown both in previous reports2,3 and in this work (Fig. 1B, open symbols). To increase the number of zebrafish surviving VHSV infection, zebrafish were injected ip (infection-by-injection) with VHSV, while the temperature was increased to 18°C to attenuate infection, thus resulting in 20%–25% (n=2) mortalities (Fig. 1B, closed symbols). Both at 14°C or at 18°C and injected or immersed, the zebrafish that died after VHSV infection exhibited similar external and internal petechial hemorrhages, exophthalmia, and congestion of internal organs. About 2 months later, the survivors of the infection-by-injection at 18°C were challenged with VHSV by infection-by-immersion at 14°C, resulting in mortalities of 10%–30% (n=2) (Fig. 1C, closed symbols). Slightly higher mortalities (∼40%) were obtained for survivors of the infection-by-immersion at 14°C challenged by infection-by-immersion at 14°C (Fig. 1C, open symbols). Two different groups of zebrafish surviving VHSV by infection-by-injection at 18°C (first infection) and infection-by-immersion at 14°C (second infection) corresponding to Experiments 1 and 2 (Fig. 1C, closed symbols), were used to obtain two batches of pooled plasma for the study.
To evaluate the presence of anti-VHSV activity in zebrafish plasma, VHSV was adjusted to obtain 200–300 DAB-positive ffu per well in VHSV-infected EPC cell monolayers 1 day after monolayer infection (Fig. 2D). To prevent fish-to-fish variation due to individual differences in complement-dependent neutralization, all the plasma samples were heated to inactivate their complement components prior to their assay (de-complemented plasma). A range of dilutions of de-complemented plasma from several immunization procedures were incubated with VHSV, and then used to infect EPC cell monolayers. Dilutions of pooled plasma (corresponding to ∼75–300-fold diluted sera) from survivor zebrafish showed the highest VHSV-neutralizing activity, with 6–12 μg/well causing 50% reduction in the number of ffu per well, depending on the batch of survivors (Fig. 2A). Titers of >30 μg/well were obtained under the same assay conditions when plasma from fish injected with G and frg11 was tested. The addition of fresh pooled plasma as a source of complement to the virus–plasma incubation mixtures reduced the above mentioned titers (or increased their neutralization activity). Thus, in the presence of a source of zebrafish complement, only 5–5.5 and >25 μg/well were required to reduce the ffu by 50% for survivor and injected zebrafish, respectively. The use of >50 μg/well of fresh plasma as a source of complement caused cytotoxic effects on the EPC cell monolayers (data not shown).On the other hand, the use of carp plasma as an alternative source of complement was discarded because it resulted in a 50%–70% reduction of ffu, and it was also toxic to the EPC cell monolayers (data not shown). The size profiles of the DAB-positive ffu correlated with the number of ffu, being higher for the wells with greater numbers of ffu and vice versa (Fig. 2C). These observations show that both the number and size of ffu were inhibited by plasma from fish immunized against VHSV.
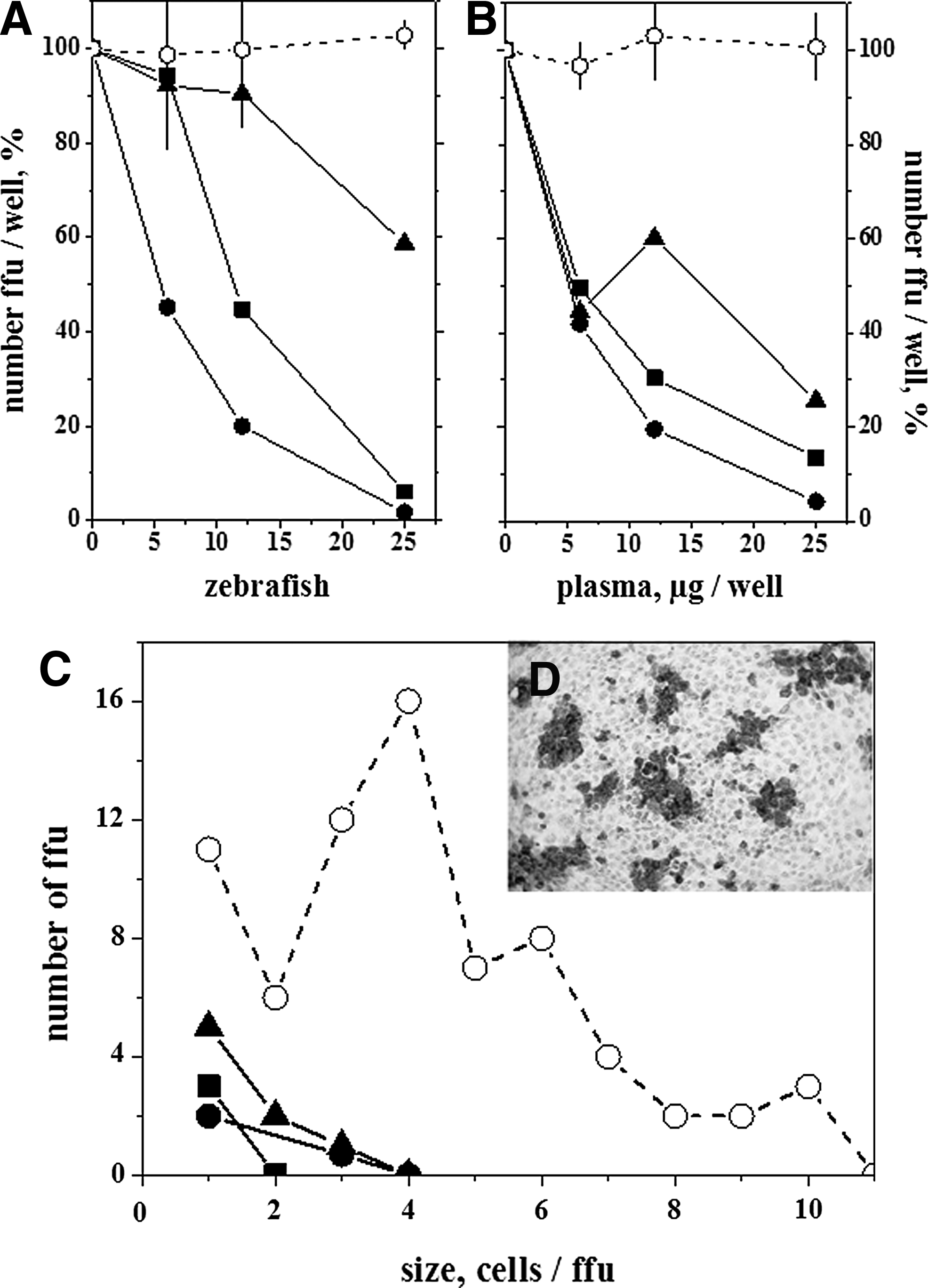
Zebrafish plasma protein concentration dependence of VHSV focus forming units (ffu)
Evidence suggesting that the observed neutralization activity was due to the presence of zebrafish immunoglobulin M (IgM) antibodies in the plasma from immunized zebrafish was obtained by the capacity of protein A-sepharose beads, anti-IgM, or protein A-sepharose beads+anti-IgM antibodies to significantly reduce neutralization of VHSV by plasma from survivor fish (Table 1). Supernatants resulting from the above mentioned treatments showed reduction of the levels of heavy chain of zebrafish IgM by Western blotting according to their reduced neutralization activity (Table 1).
Plasma from zebrafish surviving VHSV infections or from noninfected zebrafish were treated overnight with PBS, with protein A-sepharose beads, anti-IgM, or with protein A-sepharose beads, and then with anti-zebrafish IgM. Supernatants obtained after centrifugation were then assayed for the presence of the heavy chain of IgM by western blot and for VHSV neutralization, as described in methods. Under the conditions used, the extent of the in vitro VHSV infection or percentage of ffu in the absence of zebrafish plasma varied from 31.1% to 64.7%, depending on the experiment. The inhibition of neutralization was calculated in percentages by the formula: 100×percentage of ffu obtained with treated plasma from survivor zebrafish/percentage of ffu obtained with treated plasma from noninfected zebrafish. Treatment of purified VHSV with anti-VHSV MAbs C10, I16 and 3F1A12 (neutralizing Abs against the protein G 24 ) reduced the ffu to 2.3±1.93% (n=4). PBS, supernatant from zebrafish sera treated with phosphate-buffered saline. Anti-IgM, supernatant from zebrafish plasma treated with rabbit antiserum obtained against the recombinant CH4 domain of the heavy chain of zebrafish IgM (titre of 1/100000 to CH4); Protein A, supernatant from zebrafish plasma treated with protein A-sepharose beads. Protein A+anti-IgM, supernatant from zebrafish plasma treated with protein A-sepharose beads and the resulting supernatant treated with anti-IgM. The means and standard deviations from n experiments were tabulated. *Significantly different from the percentage obtained after the PBS treatment at the p<0.05 level.
To provide an example of the possible application of the micro-neutralization method to follow up immunization attempts, we tested plasma from individual zebrafish orally immunized against VHSV using plasmids coding for the VHSV protein G gene complexed with alginates. In addition, we also tested plasma obtained from individual zebrafish surviving two consecutive VHSV infections. For these purposes, 2 months after oral immunization, de-complemented zebrafish plasma samples were assayed by micro-neutralization, while 3 months later fish were assayed for resistance to VHSV infection. In this example, the de-complemented plasma samples were assayed in the absence of added fresh complement because of their possible variability. Because of the high number of individual samples to be assayed and on the basis of previous concentration dependence profiles (Fig. 2B), we used 40 μg of plasma protein per well to estimate neutralizing activity. The percentage of ffu was reduced to 20%–98% in plasma from fish orally immunized with 10 μg of pT2MCV1.4G (n=35) and to 5%–38 % (n=5) in plasma from VHSV survivors when compared to ffu of plasma from pT2MCV1.4 immunized fish (Fig. 3). The plasma samples from fish orally immunized with 10 μg of pT2MCV1.4-G, or survivors of VHSV infection were significantly different from those immunized with pT2MCV1.4 at the p<0.05 level (Student t test).
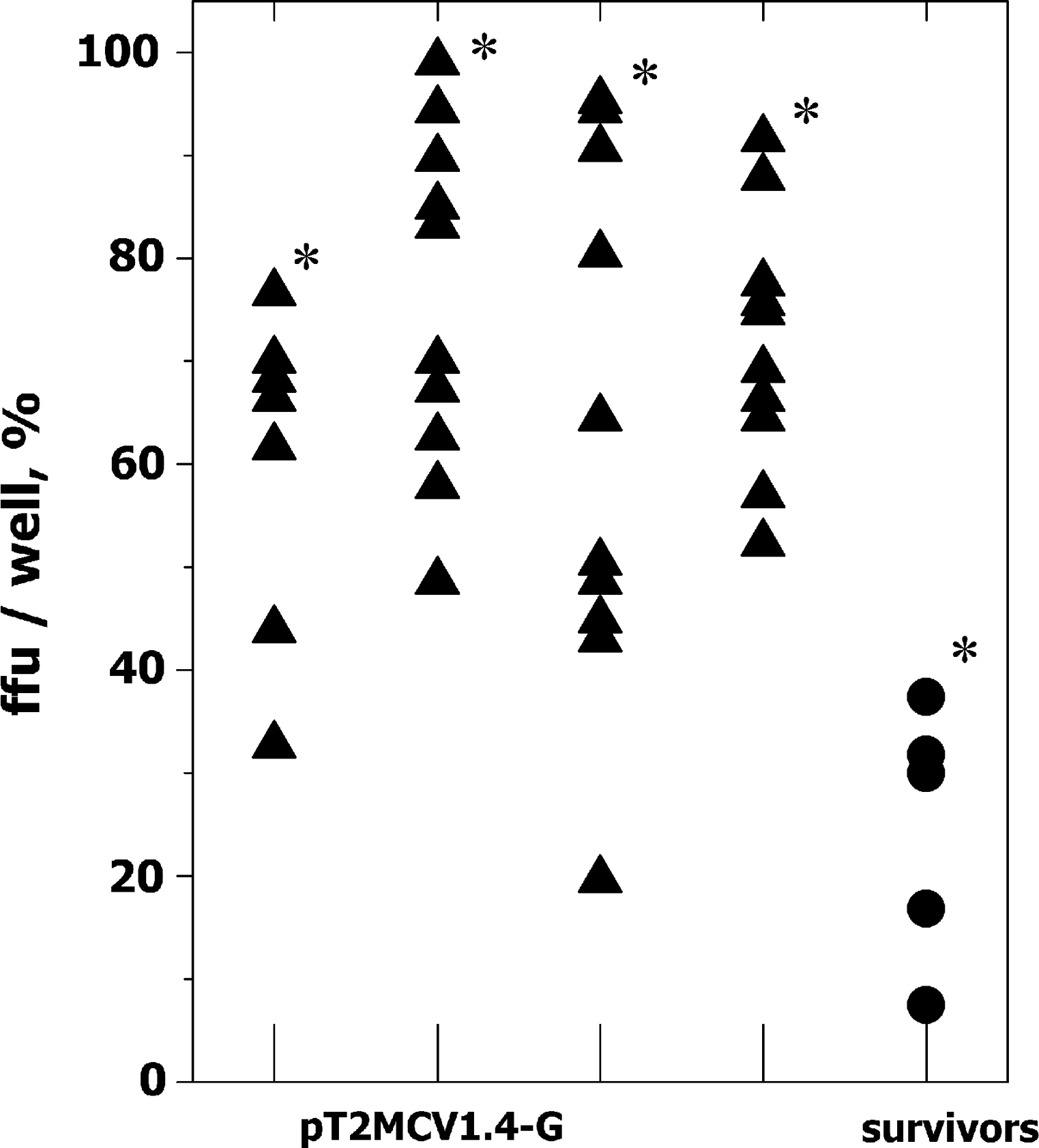
Ffu in EPC cell monolayers infected with VHSV after being incubated with plasma from either orally immunized or survivor zebrafish. Zebrafish were immunized either orally with pT2MCV1.4 and pT2MCV1.4-G plasmids or by infection with VHSV (survivors of two infections). Two months later, the fish were sacrificed and their individual plasma harvested. Inhibition of VHSV infection was assayed by micro-neutralization of purified VHSV with 40 μg protein per well from each zebrafish plasma sample. The number of ffu in the presence of plasma from zebrafish immunized with pT2MCV1.4 (214±32) was used to normalize the data by the formula 100×number of ffu in the presence of plasma from each pT2MCV1.4-G orally immunized or survivor zebrafish/number of ffu in the presence of pT2MCV1.4. The results of one experiment of two are shown in the figure. Each point corresponds to an individual zebrafish: ▲, plasma from zebrafish from 4 distinct aquaria orally immunized with 10 μg of pT2MCV1.4-G per fish; ●, plasma from zebrafish surviving two VHSV infections. *Significantly different from pT2MCV1.4 at p<0.05 (Student t test).
Zebrafish immunized in parallel by the same protocols as those used to obtain the plasma for the micro-neutralization assays mentioned above were given a VHSV challenge 3 months after immunization, and their mortality rate was followed for 25 days. At the end of this period, most VHSV survivors showed resistance to VHSV infection (89.5% RPS), the fish orally immunized with 10 μg of pT2MCV1.4-G showed a RPS of 33.33%, and 100% of those immunized with 10 μg of pT2MCV1.4 died (0% RPS) (Fig. 4).
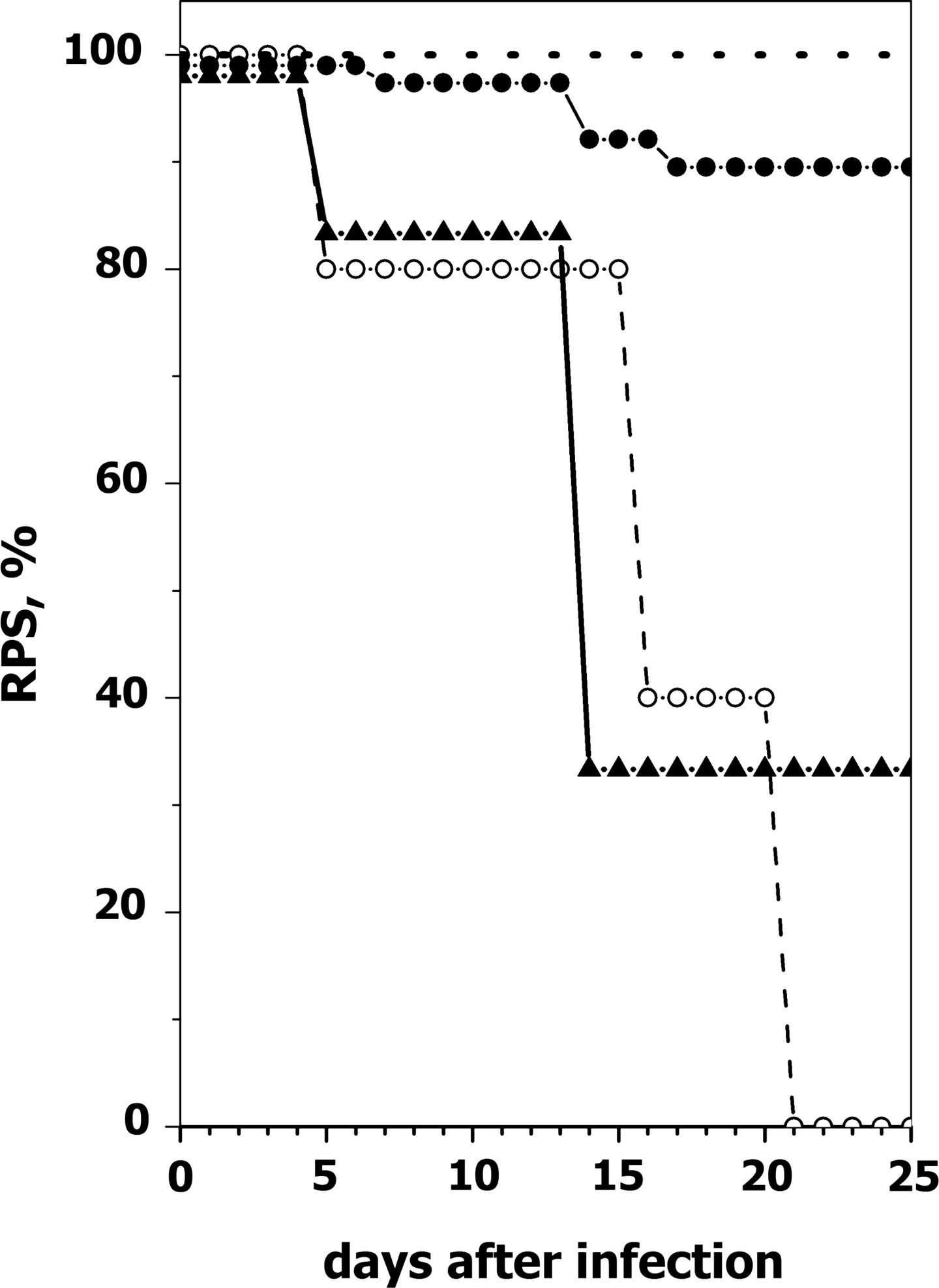
Time course of relative percent mortalities (RPS) of orally-immunized or survivor zebrafish after challenge with VHSV. Adult zebrafish (n=10 for each immunization procedure) were infected by immersion with 2×106 ffu/mL of VHSV-07.71 or virus-free cell cultured medium at 14°C in 50 mL of water. Two hours later they were released into 2-liter aquaria and maintained at 14°C. A mortality rate of 83.2% was recorded in nonvaccinated but VHSV challenged zebrafish. Relative percent survival (RPS) was calculated by the formula [1-(mortality in vaccinated fish/mortality in nonvaccinated fish)]×100. Dash line, nonvaccinated and non-challenged zebrafish; ○, zebrafish orally immunized with 10 μg of pT2MCV1.4; ▲, zebrafish orally immunized with 10 μg of pT2MCV1.4-G; ●, zebrafish surviving two VHSV infections.
Discussion
Here we successfully applied an in vitro micro-method to detect neutralization of VHSV in plasma from immunized zebrafish (Danio rerio). The neutralization was detected in low amounts of plasma from individual (corresponding to 6–10 μL of sera/zebrafish) as well as from pooled zebrafish 2–3 months after surviving VHSV infections and/or in fish orally immunized with a VHSV protein G-coding plasmid. Inhibition of neutralization by protein A-sepharose beads and/or anti-IgM suggested that the neutralization observed was dependent on the presence of anti-VHSV antibodies in zebrafish plasma.
The zebrafish/VHSV model was selected for these studies because cold-acclimatized zebrafish have demonstrated adaptive immune responses after VHSV infection and vaccination.2,3 However, VHSV is not a natural zebrafish pathogen because of their different optimal temperatures (<14°C for VHSV and 28°C for zebrafish). Therefore, little was known about the nature of the adaptive responses during the infections mentioned above, especially about the production of long-term neutralizing Abs when temperature decreases and viral susceptibility becomes maximal, as occurs during most rhabdoviral natural outbreaks in other fish species. 38 Therefore, the development of an assay with the capacity to detect long-term Ab-like responses of zebrafish is highly relevant to develop the zebrafish/VHSV as an infectious disease model. In the absence of any other well-characterized zebrafish IgM reagents, the detection of VHSV-neutralizing activity is a first approach to estimate and demonstrate such long-term antibody-like responses.
A recent survey on several fish species confirmed that some of their sera did not neutralize VHSV (Ab-negative species), even when the fish had been exposed to this virus and in vitro neutralization assays tested low dilutions (20-fold) of their sera. 9 Other fish species have VHSV inhibitory substances other than antibodies that were detectable when the fish sera was tested shortly after infection and at low dilutions. 39 The absence of VHSV neutralizing activity in nonimmunized zebrafish suggests that unspecific inhibitors were not present in their plasma, at least at the lowest dilutions (∼150-fold) that could be tested from individual specimens in this assay. In most salmonids, in vitro neutralization titers obtained at 40–80-fold dilutions are considered positive. 40 Given that similar sera dilutions in the amounts required to perform several replicates were difficult to obtain from individual zebrafish, we used plasma as an alternative. The use of this blood fraction allows for simpler harvesting; however, only >∼30-fold dilution of blood could be obtained in anticoagulant media, as measured by the amount of protein (zebrafish serum contains ∼60 mg of protein/mL, while plasma harvested by dilution had ∼2 mg/mL). Therefore, to allow for an EPC-cell-compatible media to be assayed in 96-well plates, it was not possible to assay individual zebrafish plasma at <150-fold dilutions (30-fold diluted plasma in anticoagulant media plus at least a 5-fold dilution in cell culture media). Alternative methods to obtain zebrafish plasma (i.e., bleeding into anticoagulant liquid from the tail vein) yielded a similar amount of protein plasma (data not shown).
Evidence of the presence of anti-VHSV protein G Abs in VHSV-immunized fish was suggested by using an adaptation of a similar micro-neutralization assay that detects anti-VHSV neutralization antibodies in trout 16 and by inhibition of neutralization by removing zebrafish IgM as mentioned above (Table 1). Plasma from survivors of VHSV infection neutralized VHSV in vitro with the highest titers while plasma from fish immunized against VHSV protein G by other protocols showed lower degrees of neutralization. In addition, plasma from survivors of VHSV infection showed anti-VHSV protein G activity for at least 2 to 3 months, compared to rainbow trout peaks obtained ∼6 weeks after infection. 8
Our results showed that an in vitro VHSV-neutralizing activity was present in the plasma from zebrafish surviving VHSV infections (Fig. 3), which correlated with the level of in vivo protection against VHSV challenge (Fig. 4). Nevertheless, the possibility of using recombinant protein G fragments to estimate the presence of anti-VHSV protein G Abs, independently of their neutralizing or non-neutralizing activity, would also be desirable since protection might not only be due to the presence of neutralizing Abs. Thus, in natural VHSV/trout infections it is known that: i) the number of fish showing anti-VHSV Abs by ELISA is higher than those showing neutralizing Abs any time after VHSV infection; 13 ii) non-neutralizing ELISA Abs persist longer than neutralizing Abs in survivors, 40 and iii) at least one in vitro non-neutralizing MAb provides in vivo passive protection. 43 Therefore, while VHSV-neutralizing Abs are an essential component of the protective immune response of fish against rhabdoviruses,43,44 it is not the only host response. For instance, it is known that VHSV also induces cell-mediated and other humoral defenses, most probably including non-neutralizing Abs.13,22 The notion of in vivo protective, yet in vitro non-neutralizing, Abs could also explain why neutralizing Abs are detected in only ∼50% of VHSV survivors.43,45 Despite the relatively high dilutions required for the assay, here we did not find any zebrafish surviving VHSV infections that had no detectable in vitro neutralizing activity; however, we examined a small sample of fish.
Some degree of plasma neutralizing activity (Fig. 3) and protection against VHSV challenge (Fig. 4) were also achieved in zebrafish orally immunized with pT2MCV1.4-G. Although these results appear to be promising, they need to be confirmed to increase confidence. Nevertheless, to our knowledge, this is the first report of fish immunization against rhabdoviruses using oral delivery of plasmids coding for their protein G, although previous immunization using a similar oral approach has been reported, for instance, for the infectious pancreatic necrosis virus (IPNV) of salmonids,34,46 a nonrhabdoviral virus.
The use of the immunoperoxidase technique for the micro-neutralization assay has several advantages over other possible methods to detect VHSV-positive foci, such as the absence of background staining, the use of ordinary light microscopy, and the possibility to store results for a long time. Due to its high titer, the anti-N MAb 2C9 could also be used for a simpler direct immunoperoxidase assay by conjugating peroxidase to the MAb rather than by using it as a second Ab. Large numbers of plasma samples from zebrafish immunized with VHSV can now be assayed with a high degree of specificity and in a short time (about 1 day) using this micro-method. An even simpler procedure, more reliable, and capable of being automated might be developed by using cytofluorometry to estimate the level of VHSV infection with higher accuracy by measuring fluorescence of 5000–10,000 cells per well (work in progress). The micro-neutralization assay described in the present study may be useful for following up vaccination attempts in the zebrafish/VHSV model, as well as for similar research involving other in vitro neutralizable zebrafish pathogens. Given that difficulties are encountered when using ELISA to differentiate negative and positive plasma by the anti-zebrafish immunoglobulin reagents currently available (data not shown), the micro-method described here could also serve to develop alternative ELISA detection assays for zebrafish Abs.
Footnotes
Acknowledgments
B.C. holds a FPI doctoral fellowship from the Ministerio de Economía y Competitividad (MINECO), Spain. This work was supported by CSIC project 2010-20E084, and CICYT project AGL2011-28921-CO3 and CSD07-00002 of the Ministerio de Economía y Competitividad of Spain.
Disclosure Statement
No competing financial interests exist.