
Abstract
Abstract
The electric eel, Electrophorus electricus, the only species of its genus, has a wide distribution in the Amazon and Orinoco drainages. There is little previous information regarding the population variation in E. electricus, with only basic karyotype data from two populations (Amazon and Araguaia Rivers). Karyotypic description and analysis of CO1 barcode sequences were performed for E. electricus from three localities (Caripetuba, Irituia, and Maicuru Rivers). All samples share the 2n=52 (42 m-sm [meta-submetacentric] +10 st-a [subtelo-acrocentric]) with previously studied material. However, the Maicuru River samples differ from the other populations, as they have B chromosomes. The distribution of noncentromeric constitutive heterochromatin between samples is relatively divergent. All samples analyzed present the Nucleolar Organizer Region (NOR) located in a single chromosome pair. In the samples from Caripetuba, NORs were colocalized with a heterochromatin block, whereas the NOR was flanked by heterochromatin in Maicuru River samples and pericentromeric heterochromatin adjacent NOR was found in Irituia River samples. Alignment of CO1 barcode sequences indicated no significant differentiation between the samples analyzed. Results suggest that karyotypic differences between samples from the Caripetuba, Irituia, and Amazon Rivers represent chromosome polymorphisms. However, differences between the samples from the Maicuru and Araguaia Rivers and the remaining populations could represent interpopulation differentiation, which has not had time to accrue divergence at the CO1 gene level.
Introduction
T
Along with the genus Gymnotus, Electrophorus forms a monophyletic clade, the family Gymnotidae, which is a sister group to the remaining Gymnotiformes. 2 Gymnotus also presents a large geographic distribution, being represented by 39 recognized species.1,2,5,6 Gymnotus is relatively well known from a cytogenetic point of view, with various karyotypes described from different species and populations,6–14 even including comparative genomic mapping, using chromosome painting.15,16 Such studies are important for understanding the diversity of the group and diagnosing species and populations, as well as helping to elucidate diversification processes in the genus. On the other hand, there is very little information regarding population variation in E. electricus, with only basic data about the karyotype of two females, one from the lower Amazon River and one from the Araguaia River. 17 The fact that Electrophorus produce strong electrical discharges explains the low number of cytogenetic studies and the difficulty in obtaining large sample sizes, as such studies require manipulation of the live animal to obtain chromosomes.
In the present study the karyotypic description and analysis of the CO1 barcode sequence were performed for samples of E. electricus from three new localities. We compare the results obtained with information in the literature and present discussions with respect to the mechanisms, which have resulted in karyotypic diversification.
Materials and Methods
Samples were collected from three localities in the lower Amazon basin (Table 1; Fig. 1). Voucher specimens were deposited at the Museu Paraense Emílio Goeldi. Metaphasic chromosomes were obtained from the anterior portion of the kidney 18 and analyzed using conventional staining (Giemsa), C-banding, 19 staining using the fluorochromes Chromomycin A3 and DAPI (4′6-diamidino-2-phenylindole), 20 and fluorescent in situ hybridization (FISH) using probes for 18S ribosomal DNA and telomeric sequences (TTAGGG) (Oncor©). The probes were marked with biotin or digoxigenin using nick translation and detected using avidin-Cy3 or anti-digoxigenin-FITC (fluorescein isothiocyanate). The chromosomes were organized in two groups: meta-submetacentric (m-sm) and subtelo-acrocentric (st-a). 21 At least 30 metaphases were analyzed by each method for every individual.
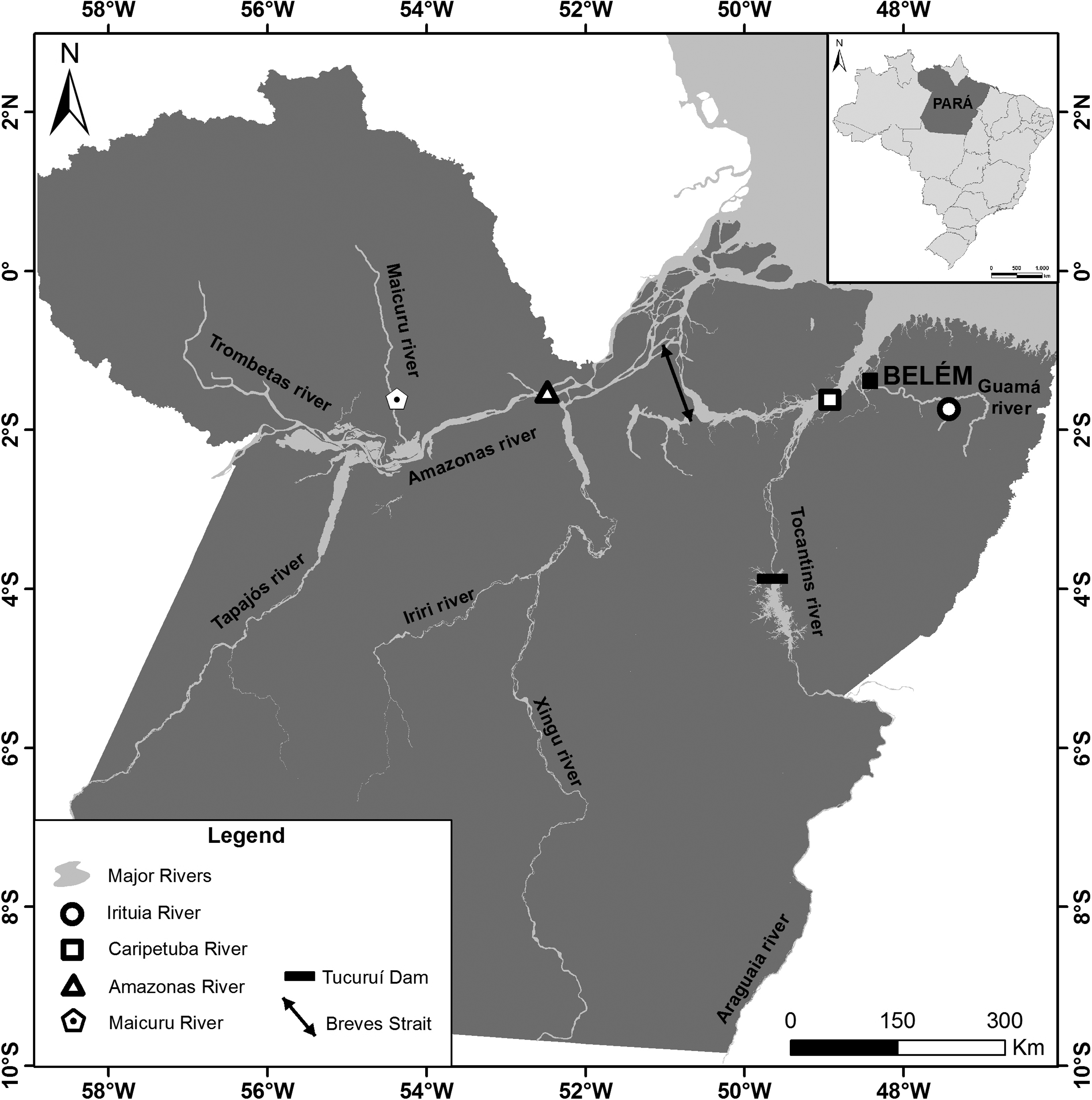
Localities of the samples of Electrophorus electricus analyzed in the present study and by Fonteles et al. 17
The last column at right shows the frequency of B chromosomes on 30 metaphases from each individual. The entire Maicuru sample has both one and two B chromosomes.
Sequencing of the DNA barcode (part of the mitochondrial cytochrome oxidase 1 gene, CO1) was carried out following protocols established by the Consortium for the Barcode of Life.22,23 Total genomic DNA was isolated from muscle tissue of each specimen using the DNeasy Tissue Kit (Qiagen), according to the manufacturer's instructions. A portion (661 bp) of the 5′ end of the mitochondrial CO1 gene was amplified by polymerase chain reaction (PCR) using the primers LIICO1F GATTTTTCTCAACTAACCAYAAAGA and LIICO1R TTYTGRTTYTTCGGACACCCAGAAGT. The 12.5 μL PCR mixes include 6.25 μL of 10% trehalose, 2 μL ultrapure water, 1.25 μL of 10×PCR buffer, 0.625 μL MgCl2 (50 mM), 0.125 μL each primer (0.01 mM), 0.0625 μL each dNTP (0.05 mM), 0.0625 μL Taq polymerase, and 2.0 μL DNA template. PCR was carried out on a Veriti 96-Well Thermal Cycler (Applied Biosystems, Inc.), under the following conditions: 3 min at 94°C; 40 cycles of 25 s at 94°C, 40 s at 52°C, and 45 s at 72°C; 5 min at 72°C. Amplified products were checked on 1% agarose gels. PCR products were labeled with the BigDye Terminator v3.1 Cycle Sequencing Ready Reaction kit (ABI) using standard methods, 24 and were bidirectionally sequenced on an ABI 3500 DNA Analyzer capillary sequencer following the manufacturer's instructions. Sequence data and trace files for specimens are available within the project Barcoding and Cytogenetics of Gymnotiformes (Samples BCG00001–BCG00011) on BOLD (The Barcode of Life Data System, www.barcodinglife.org).
Barcode sequences were aligned using the Sequencher 4.8 (Genecodes, Inc.) and analyzed using the MEGA 5.025 to provide values of pairwise sequence divergence.
Results
Samples of E. electricus from the Caripetuba, Irituia, and Maicuru Rivers present a diploid complement (2n) of 52 chromosomes and karyotypic formula (KF) with 21 meta-submetacentric pairs and 5 subtelo-acrocentric pairs (Fig. 2a–c). Moreover, five of six specimens from the Maicuru River present an additional chromosome, the microchromosomes (very small type of chromosome that have undiscriminated morphology) (Fig. 2c, f), ranging from zero to two between cells of the same organism.
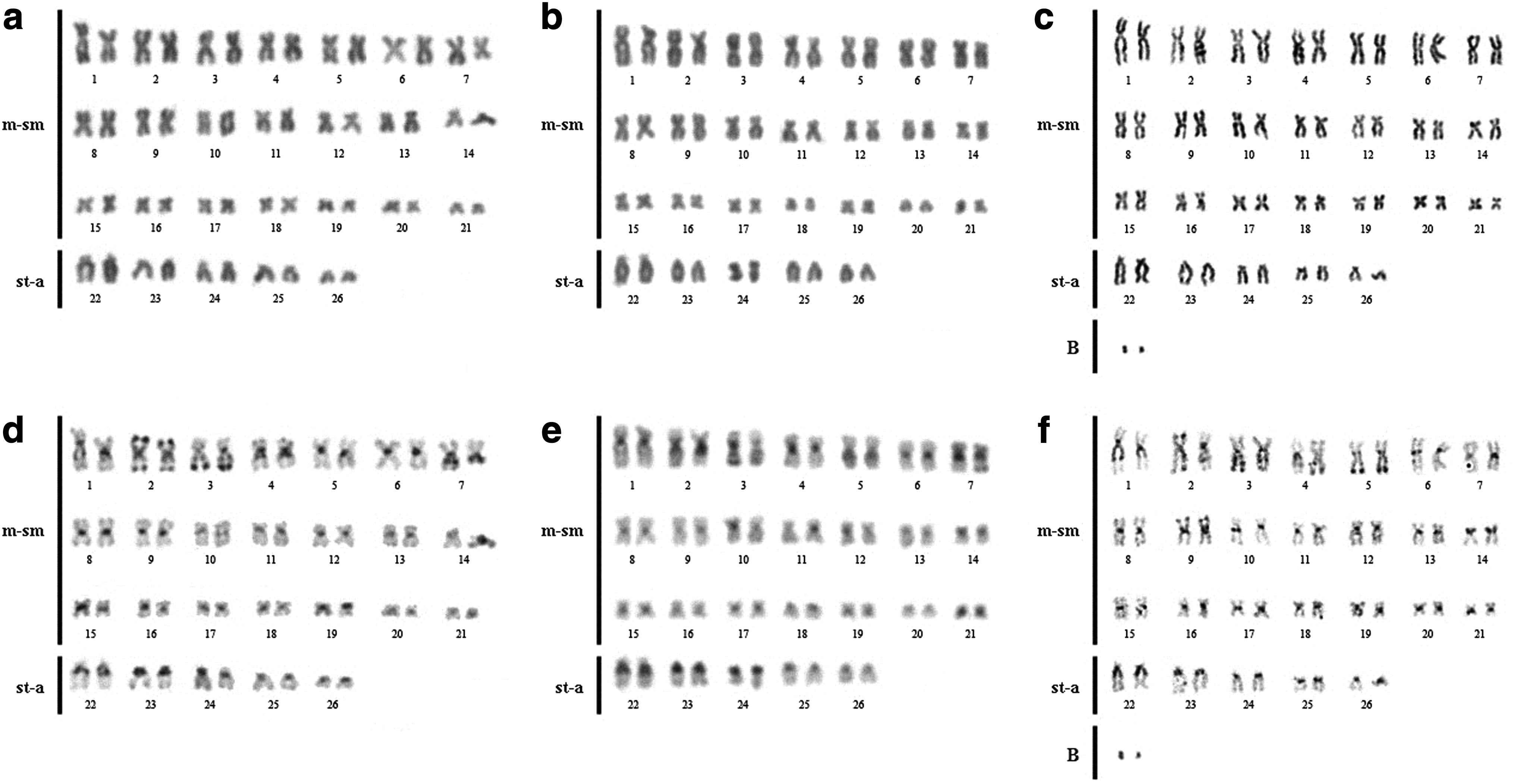
Karyotype of E. electricus from the Caripetuba, Irituia, and Maicuru rivers, respectively
In all the samples the constitutive heterochromatin (CH) was observed in the pericentromeric region of all chromosomes. Additional CH (in distal and/or interstitial regions) was also detected in pairs 1, 2, 3, 4, 7, 14, 22, and 23 of the samples from the Caripetuba River (Fig. 2d), in pairs 2, 3, 5, 7, 14, 22, 23, and 24 of the samples from the Irituia River (Fig. 2e) and in pairs 1, 2, 3, 4, 5, 9, 14, 22, and 23 of the samples from the Maicuru River (Fig. 2f). The B microchromosomes of the karyotype of the samples from the Maicuru River were found to be completely heterochromatic (Fig. 2f).
The Nucleolar Organizer Region (NOR) was localized in the proximal region of the long arm of pair 1 in samples from the Irituia, Maicuru, and Caripetuba rivers (Fig. 3). These NORs were confirmed by the FISH technique using the probes for 18S rDNA. The NOR showed positive signal for CMA3 and negative signal for DAPI. Heteromorphisms in the size of NORs were identified in all individuals analyzed. In the samples from the Caripetuba River the NOR is flanked by CH blocks, whereas in the samples from the Maicuru River the NOR is colocalized with a block of CH and in samples from the Irituia River the NOR is adjacent to pericentromeric CH (Fig. 3).
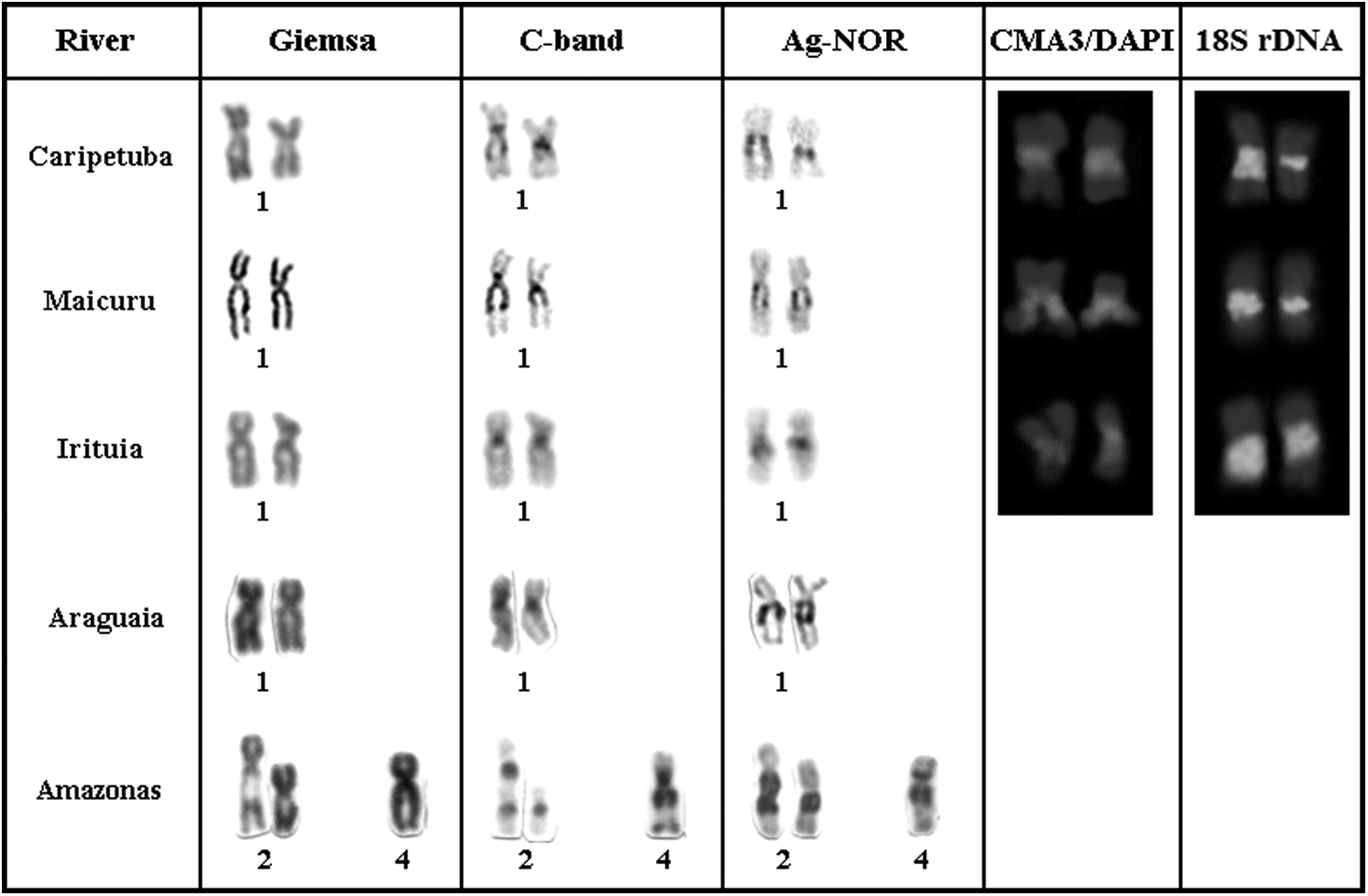
NOR-carrying chromosomes in the samples of E. electricus analyzed by: Giemsa, C-band, Ag-NOR, CMA3/DAPI, and 18S rDNA. Also shown: samples analyzed by Fonteles et al. 17
The alignment of CO1 barcode sequences indicated no significant differentiation between the samples analyzed, with only two haplotypes encountered: one encountered at all localities sampled and one from an individual from the Caripetuba River, which presented a mere 0.76% pairwise sequence divergence in relation to the other samples (five transitions at the third base codon position, with no amino acid change).
Discussion
Cytogenetic aspects
Existing phylogenies for the Gymnotiformes place the clade E. electricus+Gymnotus (Gymnotidae) as a basal lineage in this order.2,3,26 As such, knowledge of the karyotypic diversity of the Gymnotidae is essential to clarify the chromosomal changes that have occurred during the evolution of the Gymnotiformes. Karyotypic data have also proved useful in determining the biological diversity by characterizing populations and species. In this context cytogenetic contributions have been made for various species of Gymnotus,8,13,15 but are limited to only two individuals, one from each of the two populations of E. electricus in which some differences were encountered related to the distribution of CH and the NOR. 17
All samples of E. electricus analyzed in the present work share the 2n=52 complement and KF=42 m-sm+10 st-a with the specimens previously analyzed. 17 This conservative pattern of 2n and KF is in opposition to what has been found in species of the genus Gymnotus, which generally display divergent chromosome numbers between populations. 13 However, numerical chromosome variations have been found in the population from the Maicuru River due to the occurrence of microchromosomes in some individuals that also show different frequencies of these elements (0–2) in their cells. This pattern indicates that the extra microchomosomes in this population are B chromosomes. These elements are additional to the normal karyotype. They are dispensable, do not have a Mendelian pattern of transmission, and their occurrence is relatively common in fishes. 27 In Gymnotiformes the first case of B chromosomes was described in a population of Apteronotus cf. albifrons from the upper Paraná River floodplain 28 and these elements represent a karyotaxonomic character for this population, since B chromosomes had not been found in a population from Marajó Island, Amazon River delta.28,29 Occurrence of B chromosomes in unrelated species (E. electricus and A. albifrons) indicates independent origins of these elements in Gymnotiformes.
The general pattern of distribution of CH is relatively similar among the populations of the present study and those previous analyzed, 17 but there are small differences with respect to the chromosome pairs bearing nonpericentromeric heterochromatin blocks, mainly the CH blocks associated with the NOR. This may result from the occurrence of translocation of short chromosome segments, which can change the chromosome size and, therefore, the classification of the chromosome pairs. In some species of the genus Gymnotus (including cryptic species of Gymnotus carapo) chromosome translocations are very common.15,16 Alternatively, it is possible that these differences may result from different levels of chromosome condensation, which may interfere in the classification of the chromosome pairs. The heterochromatic nature of the microchromosomes of E. electricus reinforces that they are B chromosomes. 27
The localization of the NOR in samples analyzed in the present study coincides with samples from Amazon and Araguaia Rivers, 17 being localized on the proximal region of the long arm of one chromosome pair (pair 1 for samples from the Araguaia, Caripetuba, Irituia, and Maicuru Rivers and pair 2 for the sample from the Amazonas River). The difference in the NOR-bearing chromosome pair among the populations can be explained by the same processes responsible for variations in CH.
In the previous karyotypic study of Electrophorus 17 the occurrence of an additional NOR on the proximal region of the short and long arms of just one of the homologues of pair 4 in the sample from the Amazonas River was also described (Fig. 3). According to those authors, this should be considered a sporadic case. In fact, FISH with 18S rDNA probes confirm the occurrence of these sequences only in one chromosome pair (Fig. 3). However, the additional NOR present on pair 4 in the specimen from the Amazonas River differs from the NOR present on pair 2, which is flanked by CH blocks, by being colocalized with a CH block. The association of the NOR with CH (Fig. 3) could facilitate chromosomal rearrangement processes and transposition events because of the common presence of transposable elements in the heterochromatin.30,31 This could explain the occurrence of the NOR in more than one pair of chromosomes in the sample from the Amazon River. This variation could also represent a population level polymorphism where one chromosome of pair 4 has the NOR and one chromosome does not, constituting a multiple NOR type. As such, the individual analyzed would be a heterozygote. Among Gymnotiformes, multiple NORs have been found in three of the nine karyotyped species of Gymnotus6,9,10 and in the sample of E. electricus from the Amazon River, indicating independent transposition events of major ribosomal DNA in the Gymnotidae family. Another possible explanation could be the occurrence of sexual chromosomes in the population to which this sample belongs, with rDNA involved in the differentiation of these chromosomes. rDNA has been localized in the sexual chromosomes of various organisms, including fish, mammals, insects, and plants.32–40 Analysis of a larger number of individuals of both males and females from the Amazon River and other localities is necessary to resolve this question.
Heteromorphisms in the size of NORs were identified in all individuals analyzed in this study as well as in the sample from the Amazonas River. These heteromorphisms could be explained by duplication events in sequences of rDNA, which has been frequently observed in fishes.31,41–43 The positive signal of the NOR with the fluorochrome CMA3 and the negative signal with DAPI indicate GC-rich DNA is interspersed between the sequences of rDNA, a situation frequently described in the karyotypes of fishes. 41
In the previous karyotypic study of Electrophorus 17 the NOR from pair 2 of the sample from the Amazon River was described as flanked by CH blocks, while the additional NOR (in one homologue of pair 4) was colocalized with a CH block. NOR flanked by CH blocks were also found in the population from the Caripetuba River, whereas NOR colocalized with a CH block was found in the population from the Maicuru River and NOR adjacent to pericentromeric CH was found in samples from the Araguaia and Irituia Rivers. These different types of association between NOR and CH, which are generally constituted by repetitive DNA, could be the result of the dynamism of this class of DNA, resulting in events such as unequal crossing over and transposition.30,31
Biogeographical aspects
The low level of pairwise divergence between the barcode sequences studied is comparable to intraspecific divergence values in barcoding studies of Neotropical fishes,
44
and indicates that all the samples analyzed in this study likely belong to the same species. However, the various karyotypic differences found between the samples analyzed permits the presentation of two hypotheses:
(1) There is gene flow between populations and the karyotypic variants represent chromosome polymorphisms; (2) Gene flow between the populations has ceased or been drastically reduced recently, and the karyotypic variants represent distinct differences between populations that have not yet had time to accrue divergence between the CO1 gene sequences.
The first hypothesis could be used to explain the karyotypic differences between the populations from the Caripetuba, Irituia, and Amazon Rivers, given that the differences encountered in relation to the CH and NOR. Robust differences related to NOR number and location indicate divergence between populations of Bryconamericus aff. iheringii. 45 Similarly, the interpopulation variation registered in the NOR-bearing chromosome associated with CH among some populations of Sternopygus macrurus have been associated to different species.46,47 However, as found in this study in E. electricus, more subtle differences in the repetitive nature of the NOR and CH, can also be found at the intrapopulation level and may represent polymorphisms,48,49 within a single species. Moreover, populations of E. electricus from the Amazonas, Caripetuba, and Irituia Rivers are all directly connected by the Breves narrows and the Pará River, 50 which makes gene flow between these populations likely. This can be supported by data from studies, including some populations of G. carapo 13 and Eigenmannia virescens 51 from these river systems (Caripetuba and Irituia Rivers), where conserved karyotypic forms for populations of the same species were found.
The second hypothesis may explain the difference between the sample from the Maicuru River and the remaining populations. This river is located on the Guyana shield, an area characterized by sharply increasing elevations, which could contribute to the isolation of populations of this river followed by karyotypic diversification. A study in Astyanax scabripinnis found that B chromosomes do not occur in a population at a lower altitude, but are found in populations from higher altitudes (Sierra Mantiqueira) due to an absence of gene flow between the populations, 52 a pattern consistent with the findings of the present work. Reduced gene flow, together with favorable environmental conditions, could also explain the occurrence of B chromosomes in a population of Parauchenipterus galeatus and their absence in two other populations. 53 Therefore, we consider that B chromosomes represent a marker for the population of E. electricus from the Maicuru River, as in A. albifrons from Paraná River 28 and in P. galeatus from the São Francisco River. 53
The fact that the population from the Araguaia river only differs from the populations from the Amazonas and Caripetuba rivers in terms of subtle chromosomal changes related to CH and NOR, suggests that these characteristics represent intraspecific polymorphisms as mentioned above. Additionally, the shared karyotypic characters between the samples from the Araguaia and Irituia rivers indicate a possible connection between these populations. However, it is likely that the population from the Araguaia River is isolated from the other populations to some extent by the major rapids and the Tucuruí Dam on the lower Tocantins River, 54 leading to ceased or reduced gene flow.
Our results call attention to the need for the revision and further analysis of populations of the genus Electrophorus, whose diversity is almost certainly underestimated, as well as demonstrate the importance of chromosome analysis in the characterization of species and populations.
Footnotes
Acknowledgments
Most of this research was supported by FAPESPA (Fundação Amazônia Paraense de Amparo à Pesquisa) through the National Excellence on Research Program (PRONEX, TO 011/2008) on a project coordinated by J.C.P. Other funding and supports included: FAPESPA (TO 064/2012) on a project coordinated by J.C.P.; CNPq (Edital Universal, Processo 475013/2012-3), on a project coordinated by C.Y.N.; NSF DEB-1146374 (“Aquatic Faunal Survey of the Lower Amazon”) for J.S.R.; UFPA, and CAPES. C.Y.N. (306989/2009-3) and J.C.P. (307071/2009-0) are grateful to CNPq for Productivity Grants. Collection was authorized by ICMBio (Instituto Chico Mendes de Conservação da Biodiversidade) permit 020/2005 (ICMBio Registration: 207419).
Disclosure Statement
No competing financial interests exist.