
Abstract
Abstract
The Amazon has abundant rivers, streams, and floodplains in both polluted and nonpolluted environments, which show great adaptability. Thus, the goal of this study was to map repetitive DNA sequences in both mitotic chromosomes and erythrocyte micronuclei of tamoatás from polluted and nonpolluted environments and to assess the possible genotoxic effects of these environments. Individuals were collected in Manaus, Amazonas (AM), and submitted to classical and molecular cytogenetic techniques, as well as to a blood micronucleus test. Diploid number equal to 60 chromosomes are present in all individuals, with 18S ribosomal DNA sites present in one chromosome pair and no interstitial telomeric sites on chromosomes. The micronucleus test showed no significant differences in pairwise comparisons between environments or collection sites, but the Rex3 retroelement was dispersed on the chromosomes of individuals from unpolluted environments and compartmentalized in individuals from polluted environments. Divergent numbers of 5S rDNA sites are present in individuals from unpolluted and polluted environments. The mapping of repetitive sequences revealed that micronuclei have different compositions both intra- and interindividually that suggests different regions are lost in the formation of micronuclei, and no single fragile region undergoes breaks, although repetitive DNA elements are involved in this process.
Introduction
H
Repetitive DNA sequences play key roles in adaptive processes in many organisms because they are less vulnerable to selective pressures compared to single-copy sequences.4,5,8–12 Such adaptive processes may be related to genetic variability, which enables a population to adapt to an environment.13,14 However, the environment has experienced severe damage resulting from anthropic activity, 15 and aquatic biota is constantly exposed to numerous toxic substances released daily into the water. Many substances resulting from human activity have great genotoxic potential due to their cumulative effects, which may lead to serious risks for aquatic biota.16,17
Anthropogenic activities resulting in the contamination of bodies of water also occur in the Amazon region, especially in the city of Manaus, the capital of Amazonas state. The city is situated at the confluence of the Negro and Solimões Rivers and is crossed by two Rio Negro tributary sub-basins: the Mindu and Quarenta streams. 18 Due to the precariousness of dwellings on the banks of these streams, a portion of the garbage, domestic sewage, and industrial wastewater is routinely dumped in these bodies of water, decreasing the water quality.19,20
The interaction between living organisms and the environment may promote genomic changes, which, when positive, result in organism adaptations for improved habitat exploitation.21–23 However, genomic changes are not always positive. For example, the action of genotoxic agents tends to promote chemical modifications in the DNA, which are often harmful to cells, despite the sophisticated systems of identification and repair of DNA damaged by cells. Chromosomal abnormalities and significant increases in micronuclei in blood cells have been associated with exposure to genotoxic agents.24–26
The micronucleus test (MN)27–29 has been recommended for environmental biomonitoring studies,17,30,31 mainly due to its capacity to qualitatively assess the effects of clastogenic and aneugenic agents.16,32–35 Fish have received special attention because they are efficient bioindicators of environmental quality and are capable of metabolizing xenobiotics and pollutants accumulated in the aquatic environment.36–39 Additionally, they play different roles in the food chain and bioaccumulate contaminants dissolved in water.40,41
However, few fish species manage to survive in both polluted urban streams and the different types of Amazonian waters, which are classified as white, clear, and black waters, each with different physical and chemical characteristics. 42 The species Hoplosternum littorale, considered as a monophyletic group and commonly known as tamoatá, survive in all amazonic aquatic environments, and is widely distributed in the rivers composing the local basin of Manaus, including the streams affected by human eutrophication caused by substances derived from domestic and industrial wastewater.
Tamoatá does not have migratory habits43,44 and is also considered a cryptic species found in different basins of South America with a high genetic diversity. 45 Furthermore, these fish have great commercial value because the riverside population commonly uses them as a food source.46–48
The adaptability of the tamoatá may be partly related to its heterochromatin composition and location, which may be differentially organized according to the environment in which the individual lives. Three patterns of heterochromatin distribution from different regions of Brazil and Venezuela are found: (a) centromeric and interstitial blocks44,49,50; (b) subterminal and interstitial blocks 51 ; and (c) centromeric, pericentromeric, and interstitial blocks. 52 However, cytogenetic data available for the species have shown no differences in the karyotype macrostructure between individuals collected from natural and polluted environments, although several questions regarding their genomic organization remain unanswered.
Thus, this study is aimed to assess the presence of damage in individual genomes and to map repetitive DNA sequences in mitotic chromosomes and micronuclei to determine whether different genomic organizations occur between individuals living in natural and anthropized environments and whether the repetitive DNA sequences are lost in micronuclei formation.
Materials and Methods
Seventy-two H. littorale individuals were collected for the micronucleus test, including 16 from Catalão Lake, located at the confluence of Rio Negro and Rio Solimões (3°09′25″S 59°54′49″W), and 16 from Marchantaria Lake, part of Rio Solimões (3°13′22.30″S 59°55′14.59″W). These environments are considered chemically unpolluted due to their large volume of water. 53 Furthermore, 25 individuals were collected from the Mindu stream (3°04′53.27″S 60°00′25.21″W) and 15 were collected from the Quarenta stream (3°07′49.48″S 60°00′25.21″W), which have been indicated as the most critical sites of environmental pollution in Manaus, AM, Brazil. 53 The mitotic chromosomes of 40 of the 72 individuals, five males and five females from each location were analyzed and 30 metaphases were scored for each of the individuals.
No specific permissions were required for these locations, and the individuals were collected with permission from the Chico Mendes Institute for Biodiversity Conservation (Instituto Chico Mendes de Conservação da Biodiversidade - ICMBIO - Permanent license No. 28095-1/2011). The fish were euthanized following the recommendations of the National Council for the Control of Animal Experimentation (Conselho Nacional de Controle de Experimentação Animal - Concea) Guidelines for the Practice of Euthanasia (Concea, 2013). Samples were collected from the hematopoietic organ of each individual following euthanasia to obtain chromosome preparations, muscle tissue for DNA extraction, and blood samples to perform the micronucleus tests. All procedures were previously approved by the Ethics Committee on Animal Use (Comissão de Ética no Uso de Animais - CEUA) of the National Institute for Amazonian Research (Instituto Nacional de Pesquisas da Amazônia, No. 017/2014).
Preparation of mitotic chromosomes, detection of nucleolar organizing region, and detection of constitutive heterochromatin (C-bands)
Mitotic chromosomes were obtained from kidney cells using the protocol described by Bertollo et al. 54 The method described by Howell and Black 55 was used to detect the nucleolar organizing regions (NORs), and the C-band was detected using the method described by Sumner. 56
Micronucleus test
The protocol described by Fenech et al. 57 was used to prepare the blood smear slides. The number of micronuclei from fish collected in polluted and unpolluted waters was compared using analysis of variance (ANOVA). In this model, the fish samples from both polluted environments (the Mindu and Quarenta streams) were compared with fish samples collected from both unpolluted environments (the Catalão and Marchantaria lakes).
To control the effect of site, the fish samples were nested within their respective collection sites, and the sites were compared within water quality (polluted or unpolluted). Furthermore, we used ANOVA to compare the number of micronuclei between the two polluted streams and between the two unpolluted water bodies. Residual analyses were used to examine the distribution of model residuals and to ensure that the test assumptions were not violated. All analyses were performed using the statistical software R. 58
DNA extraction, preparation of repetitive DNA sequences using polymerase chain reaction probes, and nick translation combined with counterstaining
The genomic DNA of H. littorale was extracted from muscle tissue using the basic protocol of Sambrook and Russel. 59 The following primers were used for polymerase chain reaction (PCR) amplification of 18S and 5S rDNA: 18Sf (5′-CCG CTT TGG TGA CTC TTG AT-3′) and 18Sr (5′-CCG AGGACC TCA CTA AAC CA-3′), 60 as well as 5SF (5′ -CGA GGT CGG GCC TGG TTA GTA −3′) and 5SR (5′-CTT CYG AGA TCA GAC GAG ATC −3′). 61 The primer set RTX3-F3 (5′- CGG TGA YAA AGG GCA GCC CTG) and RTX3-R3 (5′ TGG CAG ACN GGG GTG GTG GT) was used to amplify the Rex3 retrotransposon.62,63 The PCRs were performed at a final volume of 15 μL with 1 μL of genomic DNA (100 ng), 1.5 μL of 10X buffer with magnesium chloride (1.5 mM), 0.15 μL of Taq DNA polymerase (5 U/μL), 3.0 μL of dNTP (1 mM), 0.6 μL of each primer (5 mM), and Milli-Q water to complete the volume.
The amplification cycles included the following steps: (a) 18S: 2 min at 94°C (initial denaturation); 35 cycles of 1 min at 95°C (denaturation), 1 min at 55°C (primer annealing), and 1 min 40 s at 72°C (extension); and 7 min at 72°C (final extension); (b) 5S: 1 min at 94°C (initial denaturation); 30 cycles of 1 min at 94°C (denaturation), 1 min at 59°C (primer annealing), and 1 min 30 s at 72°C (extension); and 5 min at 72°C (final extension); and (c) Rex3: 2 min at 95°C (initial denaturation); 35 cycles of 1 min at 95°C (denaturation), 40 s at 55°C (primer annealing), and 2 min at 72°C (extension); and 5 min at 72°C (final extension). The primers (TTAGGG)5 and (CCCTAA)5 64 were used to amplify the telomeric sequences (TTAGGG)n at a final volume of 15 μL, with 1.5 μL of 10X buffer with magnesium chloride (1.5 mM), 0.15 μL of Taq DNA polymerase (5 U/μL), 3.0 μL of dNTP (1 mM), 0.6 μL of primer (TTAGGG)5, 0.6 μL of primer (CCCTAA)5 (5 mM), and Milli-Q water to complete the volume.
The first part of the amplification was performed at low stringency: 4 min at 94°C (initial denaturation); 12 cycles of 1 min at 94°C (denaturation), 45 s at 52°C (primer annealing), and 1 min 30 s at 72°C (extension); this was followed by 35 high-stringency cycles of 1 min at 94°C (denaturation), 1 min 30 s at 60°C (primer annealing), and 1 min 30 s at 72°C (extension); and 7 min at 72°C (final extension). The PCR products were used as probes, which were labeled using the nick translation method using 14-dATP biotin (BioNick Labeling System, Invitrogen) and/or digoxigenin-11-dUTP (Dig-Nick Translation mix, Roche), according to the manufacturer's instructions. 5S rDNA probes were labeled using 14-dATP biotin, whereas the 18S rDNA, Rex3, and telomeric sequence probes were labeled using digoxigenin-11-dUTP.
Fluorescence in situ hybridization
Hybridizations were performed according to the protocol described by Pinkel et al. 65 with 77% stringency (2.5 ng/μL 18S rDNA, 5S rDNA, Rex3 or telomeric probes, 50% formamide, 10% dextran sulfate, and 2x SSC at 37°C overnight). Avidin–Fluorescein Isothiocyanate (FITC; Sigma-Aldrich), biotinylated anti-avidin (Sigma-Aldrich), and anti-digoxigenin-rhodamine (Roche) antibodies were used to detect and amplify the probe signal, and the chromosomes were counterstained using 4′,6-diamidino-2-phenylindole (DAPI; 2 μg/mL) in VECTASHIELD mounting media (Vector).
Karyotype analysis
The chromosomes were analyzed under an Olympus BX51 epifluorescence microscope, and the images were recorded using a digital camera (Olympus DP71) and Image-Pro MC 6.3 software. The metaphases were edited using Adobe Photoshop version CS5 software. The chromosomes were measured using ImageJ software and classified according to the nomenclature proposed by Levan et al. 66 Chromosomes with both arms (metacentric and submetacentric) and with one arm (acrocentric) were analyzed to determine the fundamental number.
Results
Chromosomal characterization
Diploid number equal to 2n = 60 chromosomes, without the presence of differentiated sex chromosomes, was observed in individuals from the four collection sites, including four metacentric pairs, four submetacentric pairs, and 52 acrocentric pairs (4m + 4sm + 52a), and the fundamental number (FN) was equal to 68.
Subtle differences in the constitutive heterochromatin distribution were observed between the locations analyzed. Heterochromatic staining was predominantly found in the centromeric region of most mitotic chromosomes and in the pericentromeric and terminal regions of some chromosome pairs (Fig. 1a–c). However, the heterochromatic blocks are more conspicuous in individuals from polluted environments (Fig. 1b, c) than in individuals from environments considered unpolluted (Fig. 1a). Furthermore, the heterochromatic staining of the homologous chromosome pair 3 of individuals from the Quarenta stream differed from that of the other individuals because it exhibited bitelomeric and centromeric staining; most chromosomes had clear terminal staining, which was not found in individuals from the other sites (Fig. 1a, b).

Karyotypes of Hoplosternum littorale were subjected to C-band staining to show the heterochromatin distribution patterns in individuals collected from the Catalão and Marchantaria lakes
Regarding the NOR, specimens from both polluted and unpolluted environments had single NORs, with staining in the terminal region of both homologous chromosomes of pair 5 (Fig. 1d), coincident with secondary constriction.
Repetitive sequences in mitotic chromosomes
The 18S rDNA sites were located in the terminal regions of the short arms of both homologous chromosomes of pair 5 in individuals from both polluted and unpolluted environments (Fig. 2). However, a size heteromorphism was observed in all individuals from the polluted environments (Fig. 2b).

H. littorale karyotypes showing the 18S rDNA sites (pair 5) and 5S rDNA sites (pairs 23 and 27).
The 5S rDNA sites were not syntenic to 18S rDNA and were mapped to the pericentromeric region of both homologous chromosomes of pairs 23 and 27 (Fig. 2) However, in all individuals collected from unpolluted environments (the Catalão and Marchantaria lakes), size heteromorphism of the 5S rDNA site was observed in pair 27, and a fifth staining site was observed in the interstitial region of one homologous chromosome of pair 8 (Fig. 2a).
No interstitial telomeric sites (ITSs) were observed in individuals collected from either the unpolluted (Fig. 3a) or polluted (Fig. 3b) environments.

H. littorale karyotypes showing the physical chromosomal locations of telomeric sequences.
The Rex3 retroelement in tamoatás collected from the Catalão and Marchantaria lakes, considered unpolluted environments, exhibited a scattered distribution on chromosomes with weak staining (Fig. 4a).

Physical chromosomal locations of the retroelement Rex3 in H. littorale. Patterns found in tamoatás collected from unpolluted environments (Catalão and Marchantaria lakes)
In individuals from the Mindu stream, Rex3 accumulation was observed in the terminal region of the homologous chromosomes of pair 5 and was colocalized in the 18S rDNA region, with greater distribution of this retroelement in metacentric and submetacentric chromosomes, staining particularly in the centromeric region, including the short and long arms of the pairs (Fig. 4b). Individuals from the Quarenta stream showed Rex3 accumulation in the terminal regions of both homologous chromosomes of pairs 5, 8, and 23, and sites with high signal strength were observed throughout the metacentric and submetacentric chromosomes (pairs 1, 2, 3, and 4) and in acrocentric chromosomes (Fig. 4c).
Incidence of micronuclei
The average number of micronuclei detected per individual was as follows: 2.68 from Catalão Lake, 2.62 from Marchantaria Lake, 2.85 from the Mindu stream, and 3.33 from the Quarenta stream (Fig. 5). The number of micronuclei was similar between polluted and unpolluted environments, totaling on average 2.656 micronuclei per individual collected from unpolluted environments and 3.09 micronuclei per individual collected from polluted environments (r2 = 0.01; F3.68 = 0.456; p = 0.717).
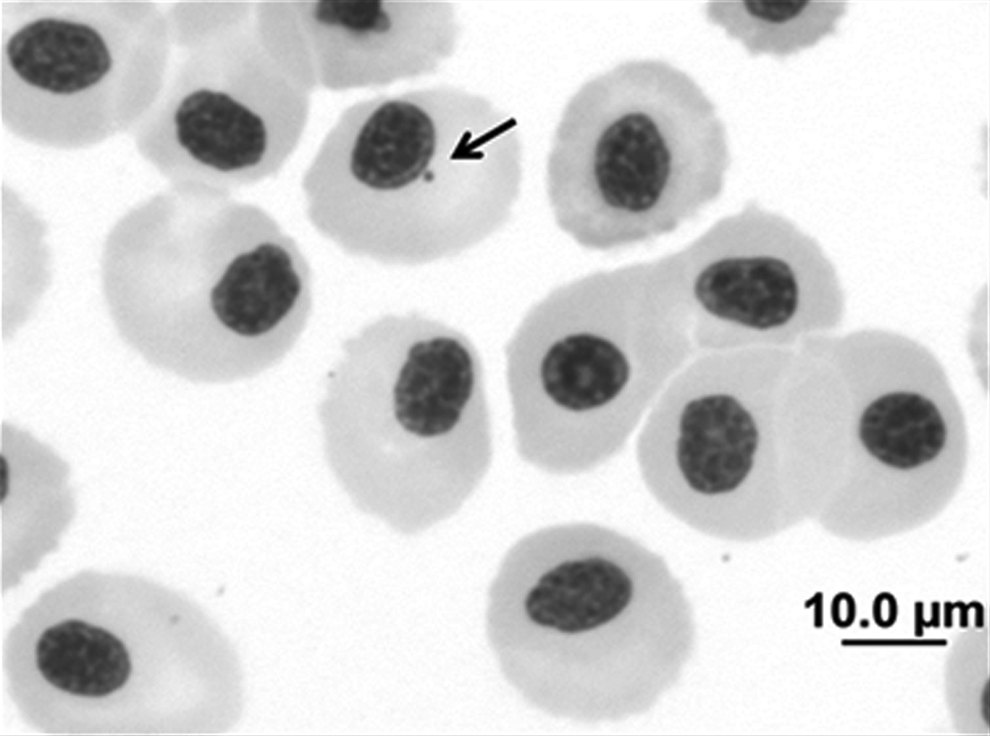
Erythrocyte cells of H. littorale stained with Giemsa stain (5%). The arrow shows the presence of a micronucleus (1000× magnification). The scale bar equals 10 μm.
The number of micronuclei also remained relatively constant between tamoatás collected from the different types of Amazonian waters (Catalão Lake = mixed water x Marchantaria Lake = white water; r2 < 0.01; F1.30 = 0.456; p = 0.923) and between Manaus urban streams subjected to different types of pollution (Mindu stream = domestic sewage x Quarenta stream = industrial wastewater; r2 = 0.02; F1.38 = 0.456; p = 0.434).
Composition of micronuclei
The mapping of repetitive DNA sequences in the chromatin of H. littorale blood cells revealed that the composition of the micronuclei was not constant and that each micronucleus has typical and intra- and interindividually variable genomic composition. The 18S rDNA sites were observed in some micronuclei and in the interphase nuclei of individuals from polluted and unpolluted environments (Fig. 6a–c). Conversely, the 5S rDNA sites, although present in the interphase nuclei, were not observed in any micronucleus (Fig. 6d–f).
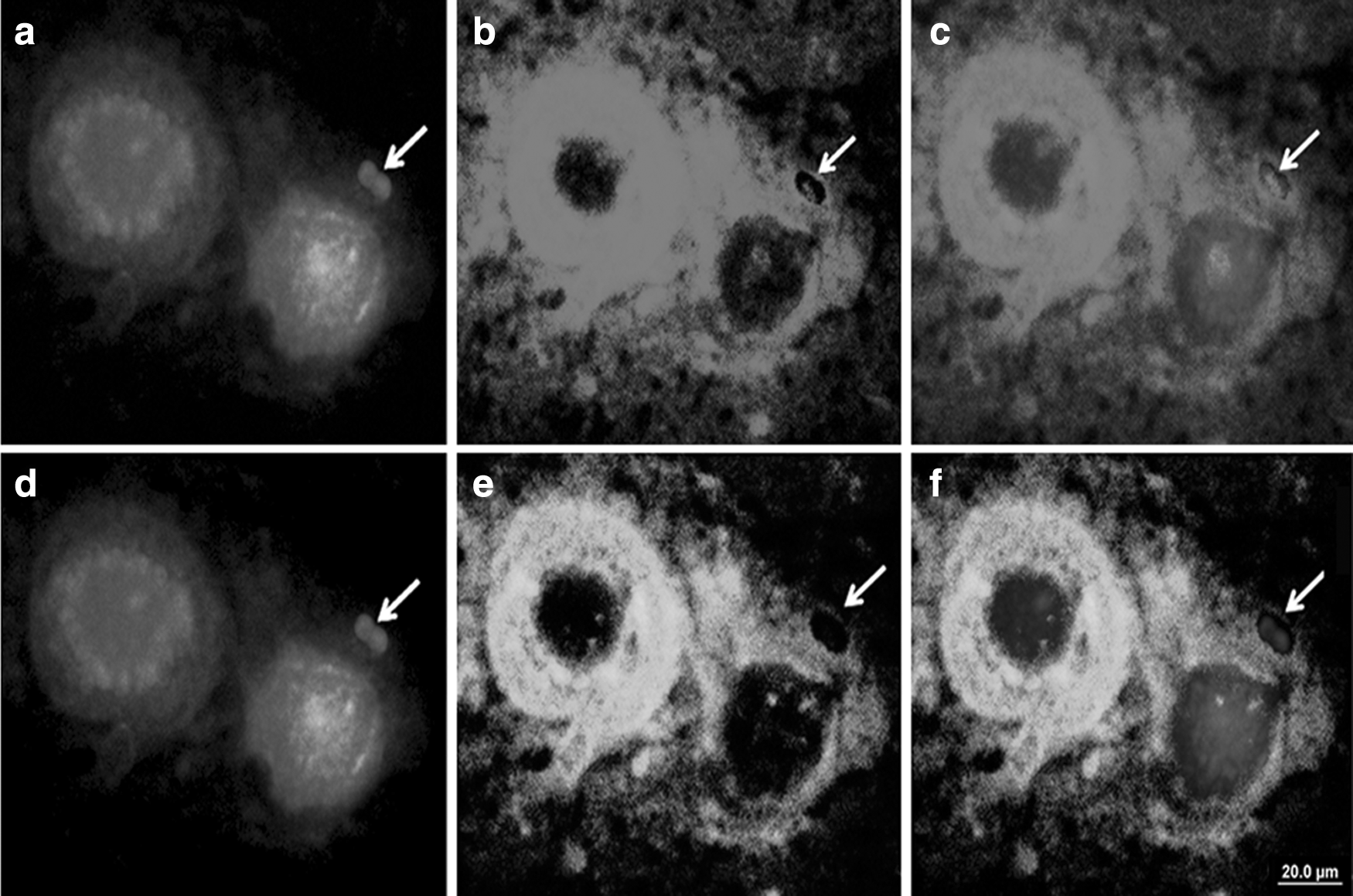
18S rDNA
Furthermore, Rex3 accumulation in some micronuclei and its dispersion in the periphery of the nuclei were observed (Fig. 7a–c). Fluorescence in situ hybridization (FISH) using telomeric sequence probes did not reveal evidence of any hybridization sites in the micronuclei found, although such sites were present in theinterphase nuclei (Fig. 7d–f).

Hybridization sequences of retroelement Rex3
Discussion
The mapping of 18S rDNA, 5S rDNA, Rex3, and telomeric sequences in erythrocytic micronuclei of tamoatá showed that the composition of micronuclei is not constant, although intraindividual variation does not occur in these sequences of mitotic chromosomes in the species. It suggests that different regions are lost in the formation of micronuclei, and no single fragile region undergoes breaks, although repetitive DNA elements are involved in this process. The variable micronuclei compositions also are observed in barley (Hordeum vulgare) root cells to analyze the effects of Gamma ray irradiation. 67 Furthermore, the presence of 18S rDNA in micronuclei of tamoatás may result in problems in the production of ribosomes, which are responsible for protein synthesis, since the 18S rDNA was shown to be active in micronuclei when subjected to impregnation with silver nitrate.
This combination of FISH technique with MN test, allowed observing the presence of 18S rDNA and Rex3 in MN composition. These repetitive elements are colocalized in the terminal region of the acrocentric chromosomes of pair 5, which are allocated in heterochromatic regions. Therefore, TEs have been reported in several studies as being colocated with ribosomal genes in different groups of organisms, 68 thus it is possible that TEs can influence the regulation of the synthesis of rRNA by recombination events due to their repetitive nature 69 and, therefore, these elements may be acting in the dispersion of ribosomal genes and consequently, causing the loss of sequences in the formation of micronucleus. Moreover, the transposition of these mobile elements is not a random process and may be controlled by host cell signals and external factors.4,70–72
The genomic organization of retroelement Rex3 is different between the streams considered polluted environments in Manaus, AM. These streams have different sources of pollution in their waters. This differential genomic organization can be related to different lineages of tamoatás, but cannot discard the possibility of environmental pollution influencing the genomic stability.
A higher number of Rex3 sites were detected in individuals from polluted environments than in individuals from unpolluted environments. The composition of Rex3 in tamoatás from the Quarenta stream, which has industrial waste and domestic sewage in its waters,18,73 showed stronger signals than that in tamoatás from the Mindu stream, which has contaminants from domestic sewage and organic residues. 74 Furthermore, detected variations in the increase in pH, high conductivity values, low dissolved oxygen levels, and high concentrations of cations and anions in the waters of these streams, with higher values in the Quarenta stream, suggest that changes in these ecosystems are related to the origins of pollution sources.75–78
Although the Rex3 distribution in the chromosomes of fish collected from polluted waters was different from that of fish collected from unpolluted waters, the DNA damage remained relatively constant (p = 0.717). Similarly, the physical and chemical differences naturally found in unpolluted Amazonian environments with mixed water and white water were not correlated with changes in the number of micronuclei in tamoatás (p = 0.923). The number of micronuclei per individual was similar (p = 0.434), even between environments polluted by wastewaters from different sources (domestic or industrial).
Chromosome break are not observed in tamoatá and diploid number is constant, but different karyotypic formulas from different localities49,50,61,67,68,79,80,present study are observed despite ITSs not being present in H. littorale. These results, combined with nonmigratory habit and different pattern of heterochromatin distribution, suggest occurrence of structural chromosomal rearrangements in isolated populations of tamoatás during their evolutionary lineage. 79
Variation in the heterochromatin pattern is common within Callichthyidae, given the occurrence of heterochromatin dispersion and amplification events within the group. 80 In tamoatá stronger staining of C-band was observed in individuals collected from polluted environments (the Mindu and Quarenta streams). The increase in heterochromatin suggests a heterochromatinization process or the addition of heterochromatin. This difference may have resulted from uneven exchanges during crossing-over processes, duplication of these sequences, or even epigenetic mechanisms, including DNA methylation or chromatin remodeling. 1
Due to the repetitive nature of sequences found in the heterochromatin, DNA polymerase errors during the addition of nucleotides in the duplication of genetic material and uneven crossing-over may cause accumulation. 81 This accumulation could explain the heteromorphism observed in the 5S rDNA and 18S rDNA sites in tamoatás, as previously observed in other fish and mammals.82–84
Association between 18S rDNA and Rex3 are observed in the present study and can be related to dispersion of ribosomal sites, as have previously been observed in other Amazonian fish species.12,85 Thus, probably 5S rDNA sites were duplicated and dispersed in the genome, since TEs are able to remove and insert themselves at other sites in the genome and are often duplicated during this process. 86 They may generate polymorphisms at their insertion site and exhibit intra- and interspecies copy number variation, or this variability observed in 5S rDNA sites in H. littorale individuals of polluted and unpolluted environments may be related to population differences.70,87–89
Thus, the differential genomic organization under different environmental conditions is present in tamoatás, although striking divergence happens in the evolutionary trends in closely related species even if they share a common origin and the repetitive sequences are involved in the micronuclei formation.
Footnotes
Acknowledgments
This work was supported by the Universidade Federal do Amazonas (UFAM), the graduate program of UFAM, Diversidade Biológica, Fundação de Amparo a Pesquisas do Estado do Amazonas (PPP grant number: 10/2011 and PRONEX/FAPEAM/CNPQ grant number: 003/2009, FAPEAM 20/2013), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (Pró-Amazônia 23038.009447/2013-45 grant number: 3295/2013), Conselho Nacional de Pesquisa e Desenvolvimento Tecnológico (grant number: 476292/2013-1), and Center for Studies of Adaptation to Environmental Changes in the Amazon (INCT ADAPTA, FAPEAM/CNPq grant number: 573976/2008-2). Species were collected with a permit issued by the Chico Mendes Institute for Biodiversity Conservation (ICMBio/SISBIO license number 28095-1/2011). FAS received funding from the Fundação de Amparo a Pesquisas do Estado do Amazonas.
Author Contributions
All of the authors performed the experiments, analyzed the data, wrote the article, and read and approved the final article.
Disclosure Statement
No competing financial interests exist.