
Abstract
Abstract
We investigated the chromosomal distribution of 18S ribosomal DNA (rDNA) in different populations of 11 species of Hypostomus collected in important Brazilian basins, namely South Atlantic, Upper Paraná, and Paraguay applying the fluorescence in situ hybridization (FISH). Hypostomus cochliodon, Hypostomus commersoni, Hypostomus hermanni, Hypostomus regani, Hypostomus albopunctatus, Hypostomus paulinus, Hypostomus aff. paulinus, Hypostomus iheringii, and Hypostomus mutucae presented multiple 18S rDNA sites while Hypostomus strigaticeps and Hypostomus nigromaculatus exhibited a single pair of chromosomes with 18S rDNA sites. The studied species presented variations in the number and position of these sites. The results accomplished were similar to those obtained by the analysis of AgNORs, revealing the same interspecific variability. Each species exhibited distinctive patterns of AgNOR and 18S rDNA distribution, which can be considered cytogenetic markers in each species of the genus and help improve the discussions on the phylogeny of the group.
Introduction
I
In fish, the location of cistrons of 45S rDNA constitutes a major cytogenetic marker, revealing groups that typically have simple NORs (Curimatidae, Anostomidae, Parodontidae, Prochilodontidae, and Cichlidae), and others that possess multiple NORs (Sternopygidae, Characidae, Lebiasinidae, Erythrinidae, and Callichthyidae).2–4 Studies on the chromosomal location of NORs in Loricariidae have shown a single chromosome pair carrier of NORs in most of the analyzed species. Nevertheless, the tribe Hypostomini exhibits multiple sites in most species. 5 Data on the distribution of ribosomal sites in Hypostomus are available for some species, viz. H. ancistroides, H. aff. ancistroides, H. commersoni, H. derbyi, H. paulinus, H. regani, H. strigaticeps, and Hypostomus aff. unae with localization of 18S rDNA sites.6–8 Data are also available for H. affinis, H. albopunctatus, H. ancistroides, H. cochliodon, H. commersoni, H. faveolus, H. hermanni, H. iheringii, H. nigromaculatus, H. paulinus, H. cf. plecostomus, H. regani, H. strigaticeps, H. tapijara, and H. topavae utilizing 18S and 5S rRNA gene probes.9–18
Considering the potential of the ribosomal DNA as a cytogenetic marker in fish, this study focused on the mapping of 18S RNA ribosomal genes by FISH in different species of Hypostomus and in some of their topotypes, that is, collected in their type locality, such as H. albopunctatus, H. hermanni, H. iheringii, and H. paulinus. Thus, it is possible to provide new data on the chromosomes of this group of fish, besides using silver nitrate impregnation to analyze the variation in number and location of activity of the existing Ag-NORs among the species investigated. Through fluorescent in situ hybridization and impregnation with silver nitrate the construction of a suitable ideogram with location of 18S rRNA genes was possible, to detect intraspecific polymorphisms of this sites and, additionally, transcriptionally inactive AgNORs.
Materials and Methods
This study comprised 11 species of the genus Hypostomus collected at different locations in three major river basins in Brazil. Some of them were captured in their type locality. The specimens were deposited in the collection of NUPELIA (Núcleo de Pesquisas em Ictiologia, Limnologia e Aquicultura) at the Universidade Estadual de Maringá. The collection sites, species deposit number, and number of analyzed specimens are shown in Table 1.
Núcleo de Pesquisa em Limnologia, Ictiologia e Aquicultura (NUP).
Chromosome preparations using conventional staining and banding techniques
The metaphase chromosomes were obtained through the air-drying technique using kidney tissue in direct chromosome preparations. 19 After thirty metaphases counts per specimen, the diploid number and karyotype were determined. The chromosomes were classified as metacentric (m), submetacentric (sm), subtelocentric (st), and acrocentric (a), as commonly used in fish.20,21 The subtelo-acrocentric chromosomes were grouped according to the previous cytogenetic studies in Loricariidae.5,6,22,23 The AgNORs were detected by silver nitrate impregnation. 24
Fluorescence in situ hybridization
We employed the FISH as proposed by Pinkel et al., 25 with some modifications, using 18S rDNA probes of Prochilodus argenteus. 26 The probes were labeled with biotin-14-dATP by nick translation according to the manufacturer's specifications (Bionick - Invitrogen).
The slides with the material were incubated with RNase (10 μg/mL) 0.04% (RNase/2×SSC) in a humidified chamber at 37°C. Denaturation of chromosomes was performed in 70% formamide/2 × SSC incubation for 4 min at 70°C. Then, we prepared 50 μL of the hybridization mixture (100 ng of labeled probe, 50% formamide, 10% dextran sulfate, 2 × SSC) per slide. The preparation was left overnight at 37° in a humid chamber. Hybridization was detected with avidin-FITC (1: 100). The chromosomes were counterstained with propidium iodide (50 μg/mL) in 50 mM MgCl2 + DABCO mounting medium.
Results
Hypostomus cochliodon, H. commersoni, H. hermanni, H. regani, H. albopunctatus, H. paulinus, H. aff. paulinus, H. iheringii, and H. mutucae exhibited multiple 18S rDNA sites, which presented variations in the number and position. Conversely, H. strigaticeps and H. nigromaculatus were the only species presenting only one chromosome pair with 18S rDNA sites. The location of these sites in all species analyzed concentrates on the short and/or long arm of the st-a chromosome type (Figs. 1 and 2). Only Hypostomus mutucae presented the 18S rDNA sites located beyond the terminal region of the long arm of the st-a pair with a size heteromorphism compared to its homolog, a marking in the interstitial region of an st-a chromosome, without displaying this marking on its homolog, confirming the same pattern observed with the impregnation with silver nitrate (Fig. 2f, f′).
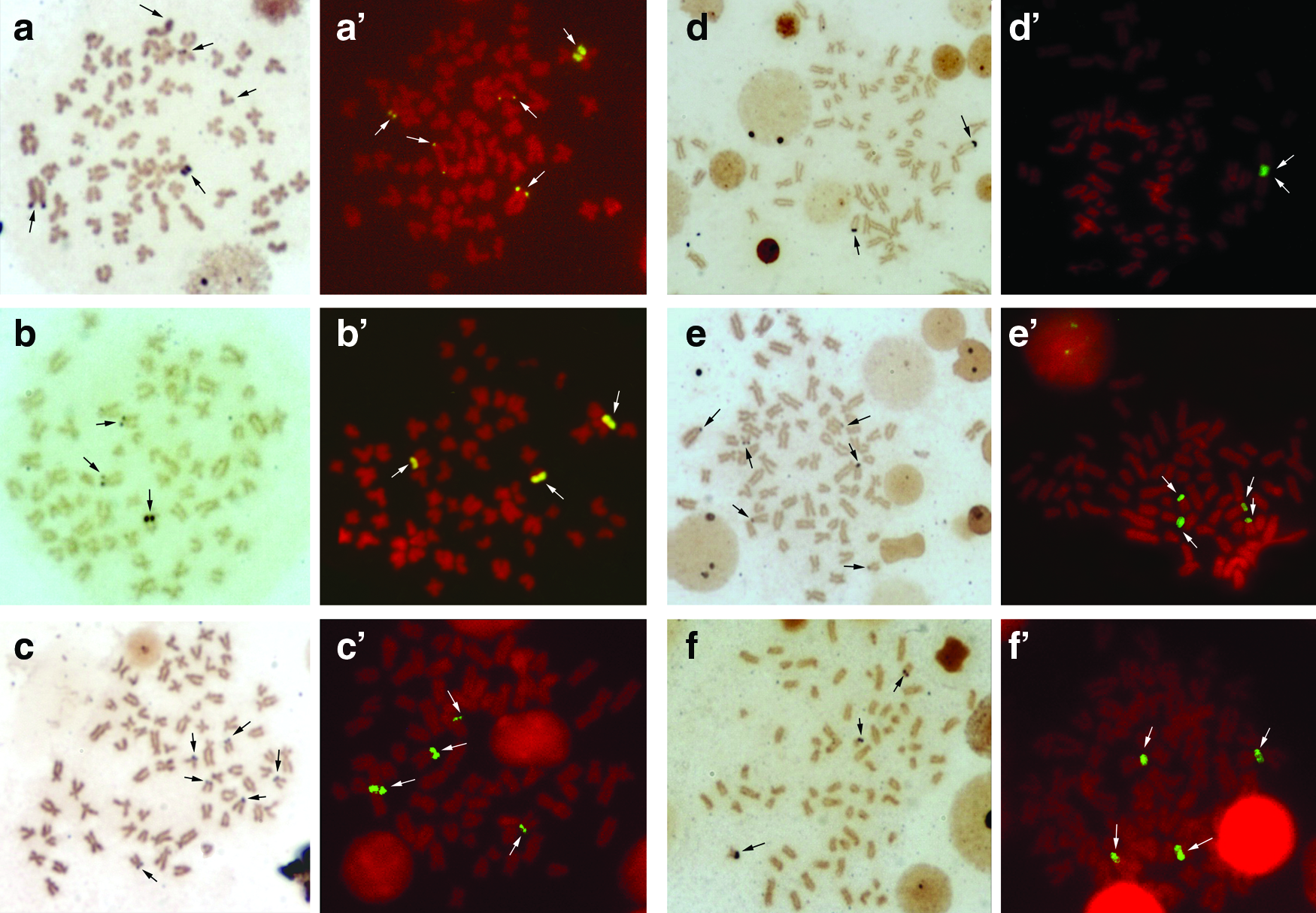
Metaphases with silver nitrate impregnation (left) and FISH (right):
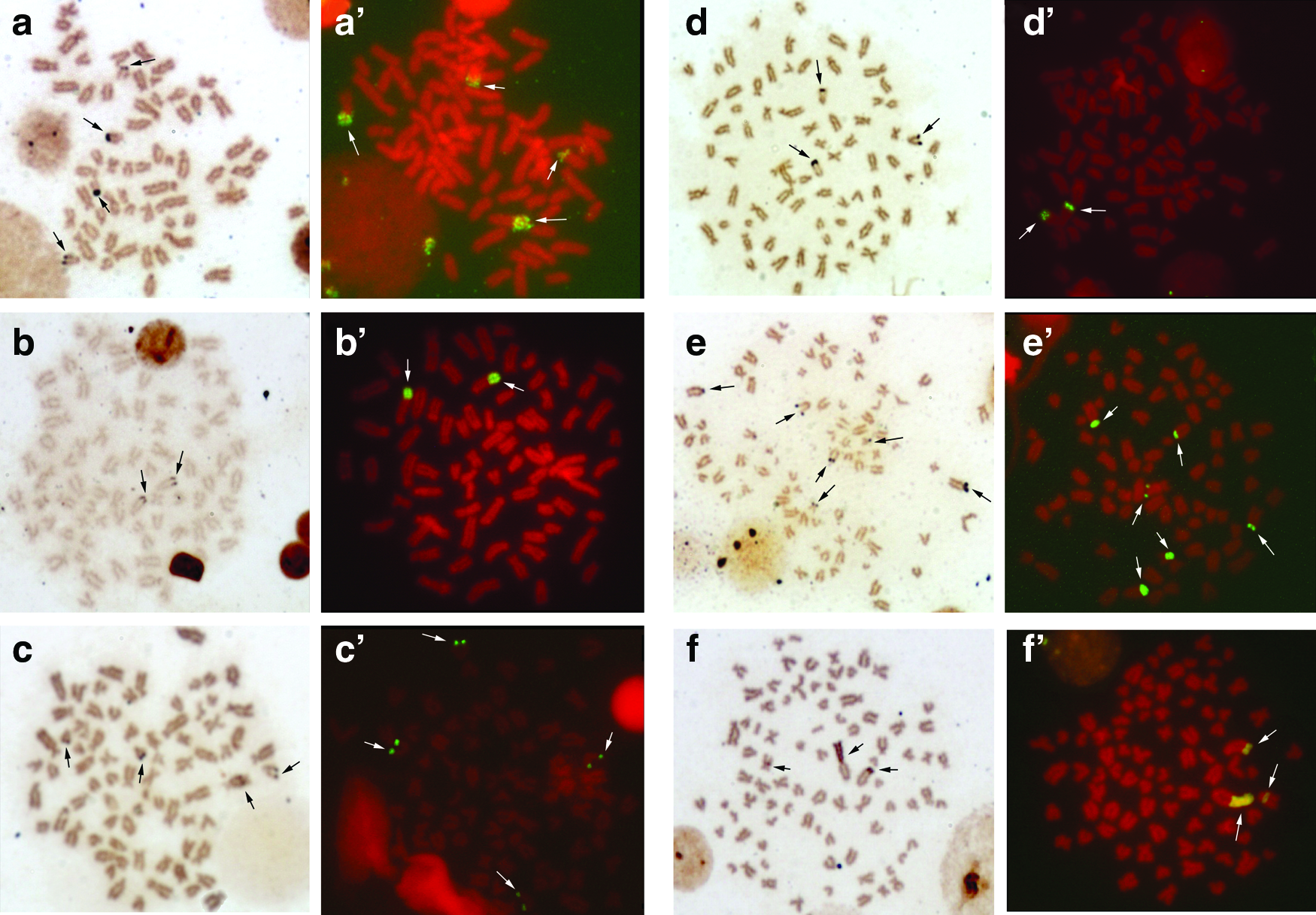
Metaphases with silver nitrate impregnation (left) and FISH (right):
Comparing the results of the silver nitrate impregnation and FISH in H. commersoni (Fig. 1b, b′), H. strigaticeps (Fig. 1d, d′), H. regani (Pirapitinga river) (Fig. 1e, e′), H. regani (Mogi Guaçu river) (Fig. 1f, f′), H. nigromaculatus (Fig. 2b, b′), H. paulinus (Fig. 2c, c′), H. iheringii (Fig. 2e, e′), and H. mutucae, those two techniques evidenced the same chromosomes in all specimens of the species analyzed. Notwithstanding, H. cochliodon (Fig. 1a, a′), H. hermanni (Fig. 1c, c′), H. albopunctatus (Fig. 2a, a′), and H. aff. paulinus (Fig. 2d, d′) exhibited intraindividual variations in the number of 18S rDNA sites. Regarding the number of 18S rDNA sites, H. cochliodon presented five phenotypes and H. albopunctatus three, as shown in Figure 3.
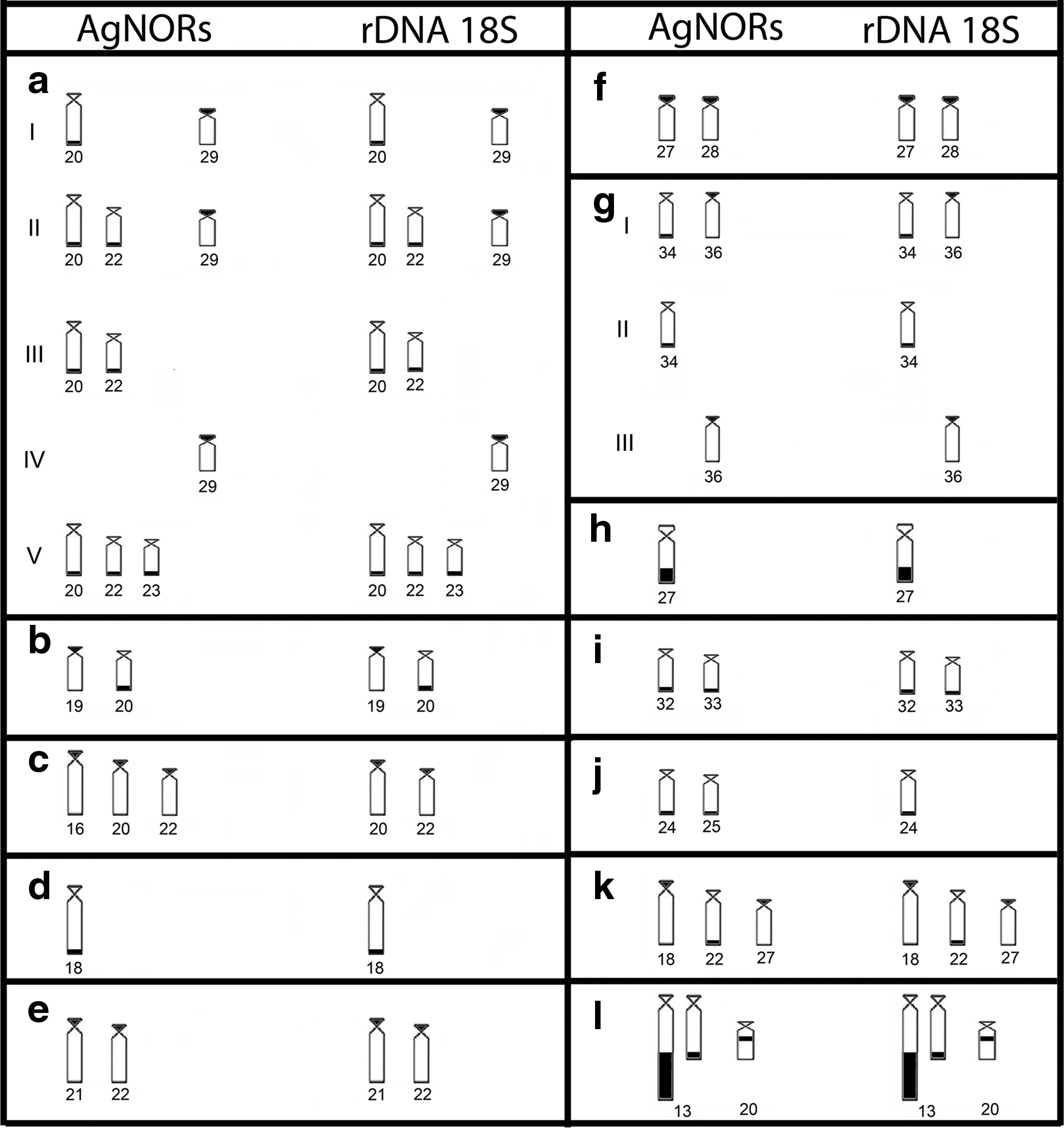
Ideogram of pairs of marker chromosomes identified by impregnation with silver nitrate (left) and fluorescence in situ hybridization (right):
Discussion
Silver nitrate impregnation is more suitable for investigating the expression of NORs, as it detects only those transcriptionally active.24,27,28 Fluorescent in situ hybridization (FISH) with 18S rDNA probe confirmed the results previously obtained by the silver nitrate impregnation, which identified multiple 18S rDNA sites in most species, except for H. strigaticeps, H. nigromaculatus, and Hypostomus aff. paulinus. Multiple NORs were also observed in the previously studied species of Hypostomus, but only with silver nitrate impregnation.5,29 Considering the number of species belonging to the genus Hypostomus, the availability of literature data with the location of ribosomal sites using FISH are scarce, with most studies focused on H. ancistroides, H. regani, and H. strigaticeps. Most of these data are restricted to the species of the Paraná River basin, more focused in the states of Paraná and São Paulo, southern and southeastern Brazil, as summarized in the Table 2 and Figure 4.
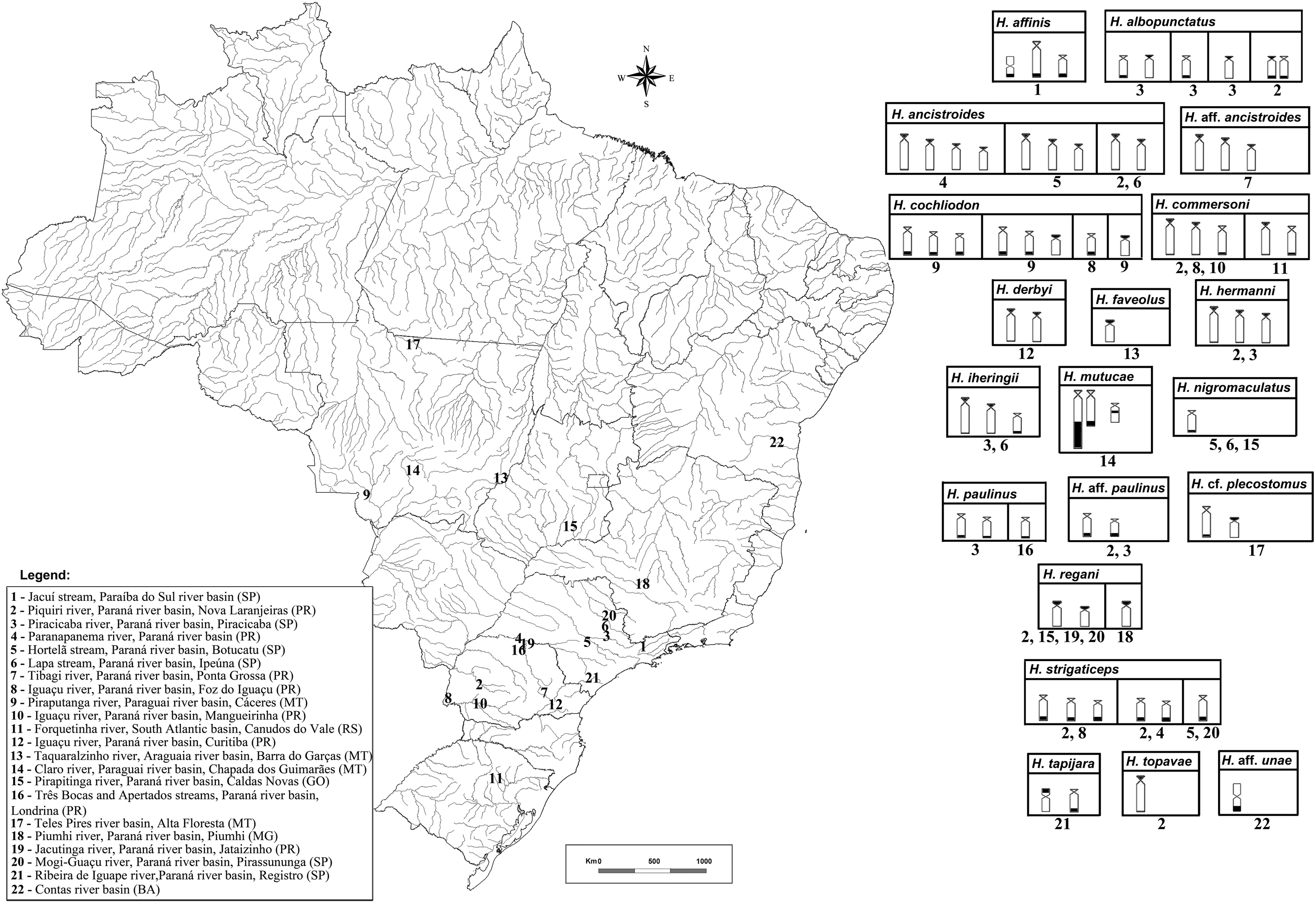
Map showing all collection points of each Hypostomus species analyzed so far, including the present work, the data are related only to works involving fluorescent in situ hybridization with 18S rDNA probes. At right the ideogram briefly shows all defaults location of 18S rDNA sites in Hypostomus species studied to date and their collection points, note that variations are shown in the boxes and the conserved patterns in some species.
Brazilian States: Bahia (BA), Goiás (GO), Mato Grosso (MT), Minas Gerais (MG), Paraná (PR), Rio Grande do Sul (RS), São Paulo (SP). Diploid number (2n), Fundamental number (FN), karyotypic formula (KF), metacentric (m), submetacentric (sm), subtelocentric (st), acrocentric (a), number of tags AgNORs, number of 18S rDNA sites, number of 5S rDNA sites. Interstitial (i), terminal (t), short arm (p), long arm (q). n.d., no data.
All species of Hypostomus analyzed to date have AgNORs located terminally on the long and/or short arm of the st-a chromosome type, and most species showed several sites.5–18 Experiments with FISH revealed structural and size heteromorphism of 18S rDNA site between the homologs of H. mutucae, and the presence of an interstitial marking, which is not common among the species of Hypostomus. This size heteromorphism was detected by both silver nitrate and FISH (Fig. 2f, f′) demonstrating that it corresponds to a structural aspect of the chromosomes.
Several phenotypes of AgNORs occur among the different subfamilies of Loricariidae. For example, in Neoplecostominae, one of the subfamilies considered basal, these sites are located predominantly in the interstitial region of a submetacentric chromosome.9,10,30 Such phenotypes are also found in the same position in most Hypoptopomatinae,31,32 while their occurrence in the terminal location is observed in other subfamilies, such as Delturinae, 10 Loricariinae,10,33,34 and Hypostominae.5–18,22,23 However, the multiplicity of NORs, detected by both silver nitrate and FISH, is a characteristic of the representatives of the tribe Hypostomini, more specifically the genus Hypostomus,5–18,22,23 this feature can be seen in Figure 4. These data corroborate the suggestions of some authors, who support that this genus has a karyoevolutionary mechanism entirely distinct from the other loricariids.5,23
Besides the significant interspecific variation in position, location, and number of NORs, showed by both silver nitrate and FISH, we also observed that some species showed an interindividual variation in the number of the NORs sites, as seen in H. cochliodon, H. hermanni, H. albopunctatus, and H. aff. paulinus, but in Figure 3 we can see that the variations always comprise the same chromosome pairs. This variation can probably be attributable to the difference in the activity of AgNORs. Also, in some cases, the silver nitrate may be impregnating acidic protein regions other than AgNORs. Concerning FISH, this may be either due to problems in detecting the sites, most of which are too small, or due to chromosomal rearrangements. Variations in the 18S rRNA genes position during the evolutionary process has often been attributed to chromosomal rearrangements of the inversion or translocation type, 35 or even to the dispersal of rDNA sites by the genome, owing to its association with heterochromatic sequences, creating new rDNA loci. Since in fish rDNA sites are most often associated with heterochromatin, 36 this convergence can make these chromosomal regions most susceptible to an unequal crossing-over. 37 Consequently, during the evolutionary process, the number of repetitive sequences can be modified by duplications and deletions, and other mechanisms, such as transposable elements adjacent to the ribosomal genes.38,39
Many authors consider that a lower diploid number, a higher number of meta-submetacentric chromosomes, and the presence of only one pair of chromosomes bearing AgNORs sites is suggested as an ancestral condition for Loricariidae family.5,29,30
This work shows that species from the South Atlantic basin, such as H. commersoni (2n = 66) from the Forquetinha river (municipality of Canudos do Vale, Rio Grande do Sul state), presents a conservative tendency regarding its chromosome number, although they possess multiple 18S rDNA sites. Conversely, species farther north of these populations, including some from the Upper Paraná and especially some from the Paraguay river basin, present a variation either in both the diploid numbers (from 2n = 64 to 2n = 82) and 18S rDNA sites, as seen in H. cochliodon (2n = 64) and H. mutucae (2n = 82).
Carvalho and Albert, 40 reported the dispersal of the ichthyofauna of the Amazon and Paraguay river basins, where the expressive similarity between species is due to the migration of populations from the tributaries of the southern Amazon river basin and populations from the Paraguay river basin. Nevertheless, studies on populations belonging to these two basins reporting the position of 18S rDNA sites are scarce (Fig. 4). Recently, Oliveira et al., 16 analyzed Hypostomus cf. plecostomus from the Teles Pires river basin (Alta Floresta, MT), which presented 2n = 68 with multiple sites of 18S rDNA, besides a ZZ/ZW system of sex chromosomes, which is an unusual feature within the group Hypostomini.
Hypostomus cochliodon (2n = 64) analyzed in this work, collected in the Piraputanga river (municipality of Cáceres/MT), a tributary on the left bank of the Paraguay river, whose sites assessed by FISH are multiple besides presenting an interindividual variability in number. This feature is different from that observed by Bueno et al., 11 in H. cochliodon (2n = 64) collected in the Iguaçu river, which showed an ancestral condition with a single 18S rDNA site.
We found that the chromosome behavior and the spread of certain chromosomal rearrangements may be different in each population in different watersheds, owing to geographical, geological, ecological, and environmental conditions. These populations may have undergone different selective pressures and/or experienced distinct evolutionary processes. Such differences may also be due to the fact that it is a very large and variable group of the genus Hypostomus known as the H. cochliodon group. 41
In Table 2, comparing different studies, we can see that Hypostomus species, which have more than one study, possess a conserved 2n and a similar distribution pattern of 18S rDNA sites, that is, predominance of chromosome of the st-a type, and location (short arm and/or long arm), which only differ in chromosomal pair that carries the 18S rDNA site in each population. These differences may be related to variation in chromosomal condensation patterns, or in the measures for chromosomal classification or be generated by structural rearrangements, such as a translocation.
Therefore, if the chromosome pairs are not taken into consideration but only the chromosomal type and position of sites, we can observe a pattern concerning all species that have more than one study, as H. ancistroides, which has 4–8 whose chromosomes were all of the st-a type, all positioned in short arm; H. hermanni (6 st-a chromosomes, short arm); H. nigromaculatus (2 st-a chromosomes, long arm); H. paulinus (2–4 st-a, long arm); H. regani (2–4 st-a, short arm); H. strigaticeps (2–6 st-a, long arm) among others, as can be seen summarized in Figure 4.
However, as the genus Hypostomus is one of the most karyotypically variable among loricariids, we cannot consider only features like 2n and 18S rDNA sites to infer about the phylogeny of the group since it is not a rule applicable to all populations. Thus, most often these characteristics only serve as population markers.
In fact, Hypostomus presents a high diversity concerning not only the diploid number and karyotypic formula,5,6,23 but also the distribution of 18S rDNA sites. This variation shows a distinct evolutionary path from the representatives of the other subfamilies of Loricariidae, 42 which may be attributable to intrinsic factors presented by the group, which have not been disclosed so far.
Considering the data already available on the location of the ribosomal genes for the genus Hypostomus, the information gathered in this work represent a valuable contribution toward a broader view of the evolution of ribosomal DNA in this group.
Footnotes
Acknowledgments
This research was supported by a grant from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). The samples were collected with the permission of Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (IBAMA) License number: 11399. This study was approved by the ethics committee (CEUA 507.2014.96) of our institution and meets all requirements of the Brazilian environmental laws.
Disclosure Statement
No competing financial interests exist.