
Abstract
Abstract
Microsatellites show great abundance in eukaryotic genomes, although distinct chromosomal distribution patterns might be observed, from small dispersed signals to strong clustered motifs. In Neotropical fishes, the chromosome mapping of distinct microsatellites was employed several times to uncover the origin and evolution of sex and supernumerary chromosomes, whereas a detailed comparative analysis considering different motifs at the chromosomal level is scarce. Here, we report the chromosomal location of several simple sequence repeats (SSRs) in distinct electric knife fishes showing variable diploid chromosome numbers to unveil the structural organization of several microsatellite motifs in distinct Gymnotus species. Our results showed that some SSRs are scattered throughout the genomes, whereas others are particularly clustered displaying intense genomic compartmentalization. Interestingly, the motifs CA, GA, and GAG exhibited a band-like pattern of hybridization, useful for the identification of homologous chromosomes. Finally, the colocalization of SSRs with multigene families is probably related to the association of microsatellites with gene spacers in this case.
Introduction
G
Previous cytogenetic studies developed in this group evidenced that interspecific diversification is followed by extensive karyotype changes, with diploid chromosome numbers ranging from 2n = 34 in Gymnotus capanema 3 to 2n = 54 in Gymnotus paraguensis, Gymnotus carapo, Gymnotus Mamiraua, and Gymnotus inaequilabiatus.4–7 Remarkably, even those species that apparently differ from the others by few chromosomal rearrangements might possess high rates of reorganization in their karyotypes. 8 In addition, differential evolutionary dynamics of multigene families (e.g., ribosomal genes and U2 snRNA) might be observed among Gymnotus species. For example, although the chromosomal location of 5S rDNA is highly variable between species, the 18S rDNA and U2 snDNA are quite conserved, with a few exceptions.6,7,9–11
As largely known, repetitive DNA sequences constitute a significant fraction of eukaryotic genomes, including transposable elements (TEs), multigene families, and tandemly repeated sequences. 12 Microsatellites or simple sequence repeats (SSRs) are tandemly organized short DNA motifs of one to six nucleotides that exhibit the fastest evolution rates among DNA sequences. 13 Previous studies indicated that microsatellites distribution is nonrandom, frequently associated with heterochromatin, although examples of clustering on the euchromatin were also described.14,15 Although these sequences constitute a substantial fraction of eukaryotic genomes, their chromosomal distribution in several closely related species has been addressed in limited fish groups, mainly on Characiformes species, to assess the evolution of sex or B chromosomes.16–21
In this study, we described and compared the physical location of eight different microsatellite motifs in five Gymnotus species belonging to the G. carapo clade.1,3 Our main purpose was to better understand the evolutionary dynamics of different microsatellite sequences in distinct species, which could allow insights into the genome organization and repetitive DNA evolution in fish.
Material and Methods
Materials
Five species of Gymnotus were analyzed, including Gymnotus sylvius, G. cuia, G. pantanal, G. capanema, and G. carapo (Table 1). Cell suspensions for chromosome preparations from all species were already available in our laboratory and their karyotypes were previously described.3,6,11
Probe preparation and fluorescence in situ hybridization
Oligonucleotide probes containing microsatellite sequences (A)30, (C)30, (CA)15, (GA)15, (GC)15, (TTA)10, (CAA)10, and (GAG)10 were directly labeled with Cy3 during synthesis, as described by Kubat et al. 22 fluorescence in situ hybridization (FISH) was performed on one male of each species, except for G. capanema. The hybridization conditions were the same for every analyzed sample and applied the following protocol, modified from Pinkel et al. 23 Slides were incubated with 0.005% pepsin/10 mM HCl for 10 min at 37°C and chromosomal DNA was denatured in 70% formamide/2 × saline-sodium citrate (SSC) for 5 min at 70°C. For each slide, 30 μL hybridization solution containing 200 ng labeled probe, 50% formamide, 2 × SSC, and 10% dextran sulfate was denatured for 10 min at 95°C, dropped onto the slides, and hybridized overnight at 37°C in a 2 × SSC moist chamber. After hybridization, slides were washed in 2 × SSC for 5 min, followed by a second wash in 1 × phosphate-buffered saline (PBS) for 1 min. Chromosomes were counterstained with 4′,6-diamidino-2-phenylindole (DAPI) (Vector Laboratories, Burlingame, CA) and analyzed using an optical photomicroscope (Olympus BX61). Images were acquired using Image Pro Plus 6.0 software (Media Cybernetics, Rockville, MD).
Results
The analyzed species exhibited the expected diploid chromosome numbers and all the mapped microsatellite motifs yielded visible signals after FISH, each showing different hybridization patterns: (i) dispersed; (ii) clustered, in which signals occur closely together in a chromosome region yielding a band-like hybridization pattern; and (iii) dispersed + clustered (mixed) (Figs. 1 and 2).
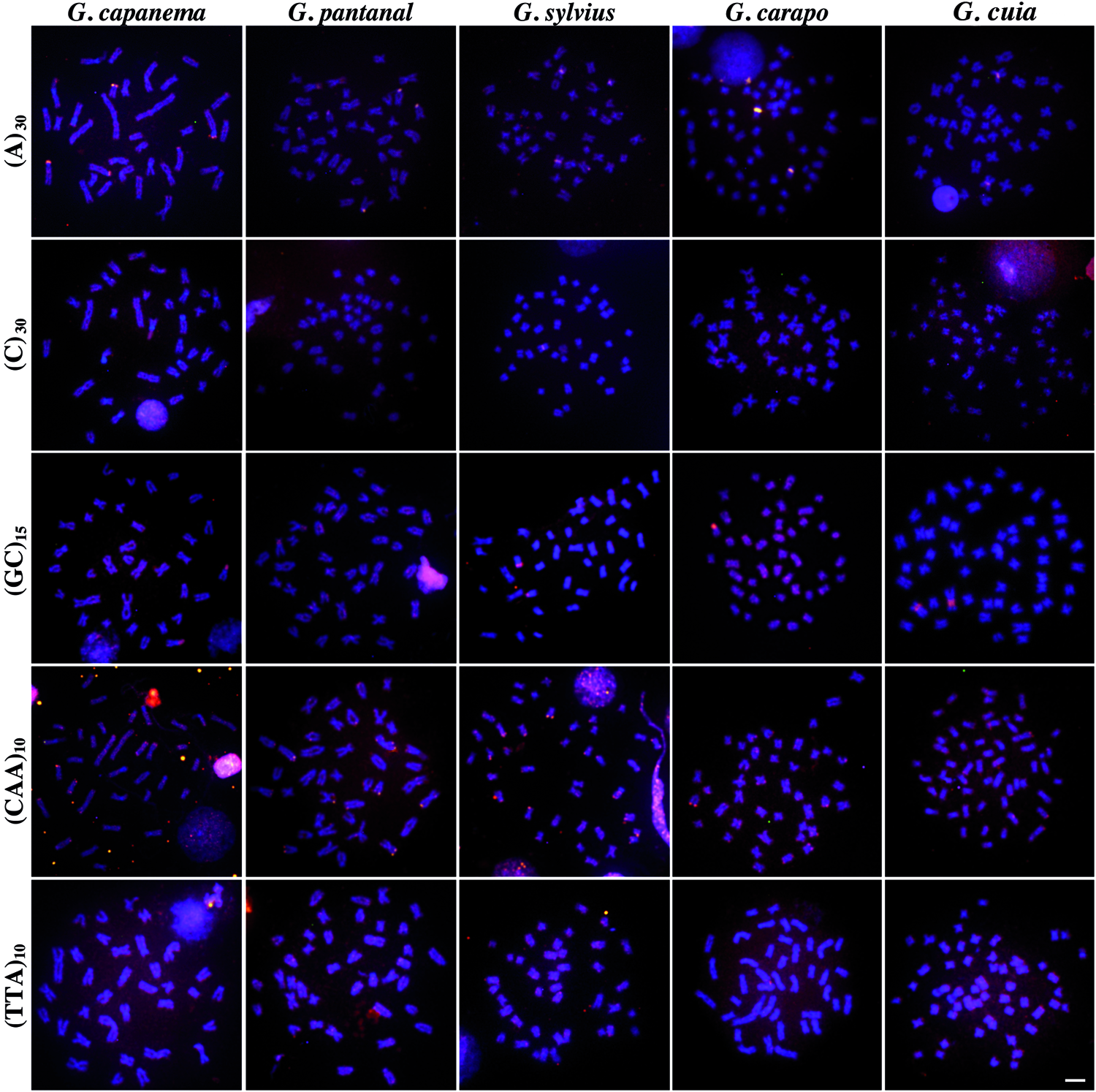
FISH mapping of SSR motifs in different Gymnotus species. Chromosomes were counterstained with 4′,6-diamidino-2-phenylindole (DAPI) and microsatellite probes were directly labeled with Cy3. Scale bar = 10 μm. FISH, fluorescence in situ hybridization; SSR, simple sequence repeat. Color images available online at www.liebertpub.com/zeb
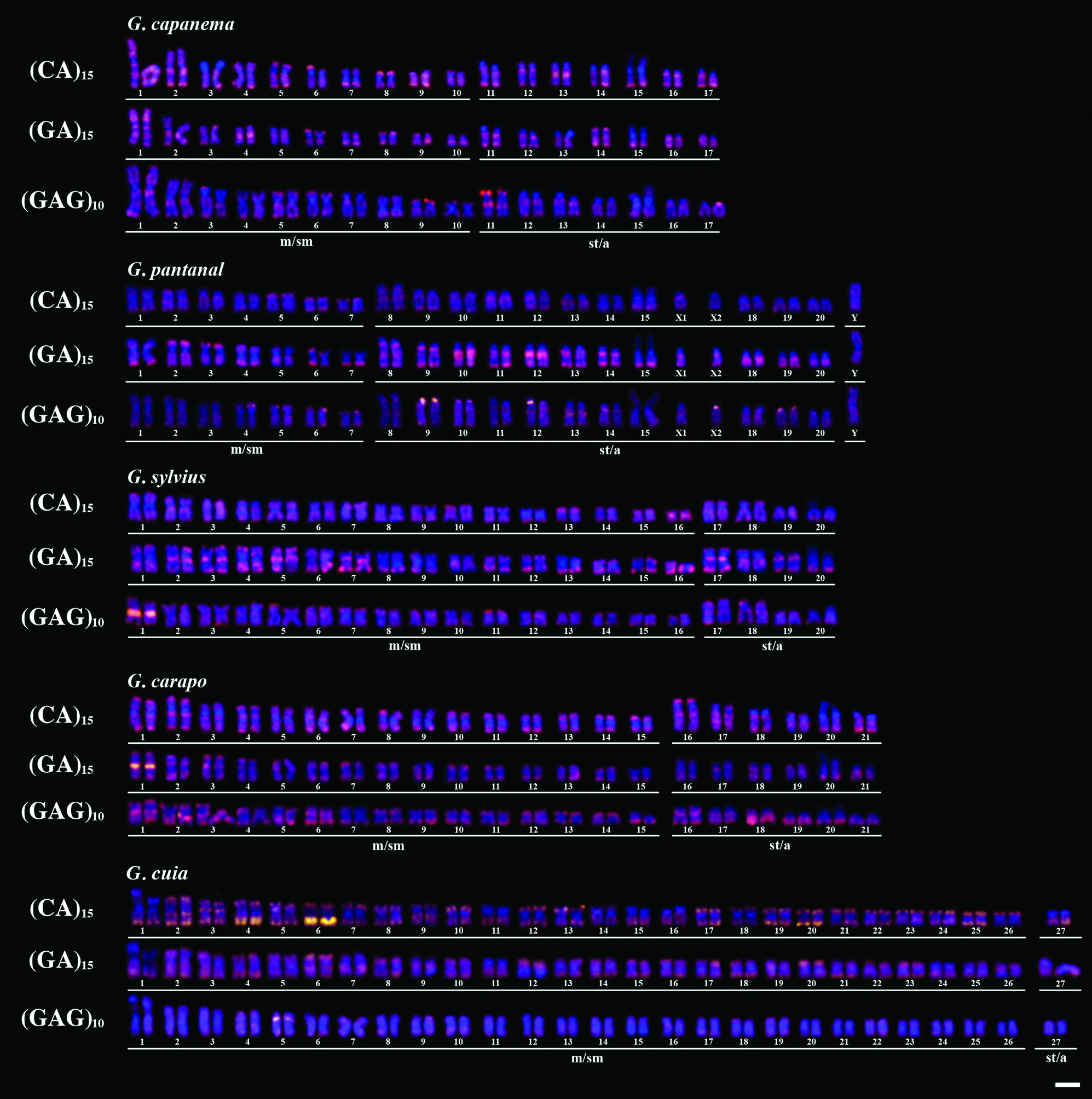
Karyotypes of five Gymnotus species after FISH mapping of compartmentalized SSRs. Scale bar = 10 μm. Color images available online at www.liebertpub.com/zeb
In general, mono-, di-, and trinucleotides were distinctively distributed. For instance, the A and C mononucleotides motifs showed a dispersed pattern throughout the chromosomes with a few clustered sites (Fig. 1), whereas the CA and GA dinucleotides exhibited conspicuous signals over pericentromeric and subtelomeric regions of almost all chromosomes (Fig. 2). Curiously, the centromeres of specific chromosomes of G. pantanal and G. capanema did not exhibite marks (Fig. 2). The GC motif showed different and particular levels of abundance in the analyzed species, although a conserved clusterization pattern involving the secondary constriction related to Nucleolar organizar region (NOR) was observed in all cases (Fig. 1). The trinucleotide GAG was also differentially distributed in terms of abundance; interestingly, this motif was highly distributed on subtelomeric and pericentromeric regions, except in G. capanema, in which the centromeres did not show evident marks (Fig. 2). The CAA motif was also organized in a mixed way, whereas the TTA was the less abundant motif here, showing dispersed hybridization patterns for all species, with small clusters only in G. pantanal (Fig. 1).
Discussion
Genomic abundance of microsatellite sequences provides an interesting opportunity to address issues such as chromosome structure and evolution. 24 In this study, the performed experiments revealed that, in general, di- and trinucleotides are more abundant than the mononucleotides, although the trinucleotide TTA was the less abundant motif. Remarkably, the different modes of genomic organization of the repetitive sequences at cytological level observed (scattered, clustered, and mixed) were already described to occur in different species and most likely reflect historical events, which might explain the intragenomic and interspecific variability of microsatellite distribution here observed.15,19,25
Although an uneven SSR distribution for Gymnotus species was found, specific patterns of microsatellite location could be observed, like the predominant clustered organization of GA, CA, and GAG, the scattered distribution of A and C, and the colocalization of GC motif with NOR regions, corroborating the hypothesis that microsatellite accumulation in eukaryotic genomes is nonrandom, and closely related species show preferential accumulation of specific SSRs with a particular chromosomal distribution.13–15 Previous studies using genetic maps have evidenced that microsatellites are spread over the entire karyotypes 26 ; however, our data revealed that particular SSRs might be localized in specific genomic regions, such as euchromatin, heterochromatin, or both (Figure S1; Supplementary Data are available online at www.liebertpub.com/zeb). In this sense, the chromosomal distribution of GA, CA, and GAG is remarkable, suggesting a compartmentalization pattern on chromosomes.
Data on different fish orders concerning microsatellites distribution evidenced that a compartmentalized organization of specific SSRs is quite common and highly variable. For instance, although the dinucleotides CA and GA are clustered in the chromosomes of several otophysian fishes (e.g., Characiformes, Siluriformes, and Gymnotiformes),16,18,20,24,27–31 their distribution pattern in Perciformes species is dispersed.32,33 Here, we showed that the clustered SSRs CA, GA, and GAG exhibited high levels of correspondence between homologues, as evidenced by the karyotypes, suggesting that differences on the distribution patterns might be due to structural rearrangements found between species. In this sense, we propose that the chromosomal mapping of different clustered microsatellites might be very useful in the identification of homologues in fish species with homogeneous karyotypes.
As largely known, microsatellites are highly dynamic sequences and distinct mechanisms, such as unequal crossing over, polymerase slippage, extra circular DNA, association with TEs, among others, might play an important role in the differential spreading and/or clustering of such sequences.15,34–36 Thus, apart from structural chromosome rearrangements, these mechanisms might also drive the interspecific dispersion and diversification of microsatellites in Gymnotus genome. Interestingly, the chromosomal mapping of different TEs in Gymnotus species, such as Tc1 and SINES, showed clustered and scattered distribution patterns, respectively, 9 evidencing that TE-mediated dispersion of microsatellites might occur in this fish group.
Examples of colocalization between microsatellites and specific repetitive DNA sequences are rather common, including ribosomal and histone genes.15,19,37 Here, the colocalization between GC motif and 45S rDNA clusters was detected in all species, whereas the A motif distribution corresponds to the U2 snRNA location in G. cuia, G. sylvius, G. carapo, and G. pantanal. 11 Such condition is probably related to the association between microsatellite repeats and multigene family spacers, similar to that observed in grasshopper genomes for histone genes. 15 In fact, the assembly of U2 snRNA from G. sylvius and G. cuia revealed an A-rich spacer for this gene (Utsunomia in preparation).
Taken together, our results indicate that microsatellite distribution in the genomes of Gymnotus species is highly variable, although some conserved features were observed, especially the distribution of those microsatellites linked with multigene families and with a compartmentalized organization (e.g., CA, GA, and GAG). Such a feature enabled an accurate identification of homologues in the analyzed species, pointing to the usefulness of microsatellites mapping for species with homogeneous karyotypes.
Footnotes
Acknowledgments
This study was supported by the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), Coordenadoria de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) from Brazil.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.