
Abstract
Abstract
Rhamdia quelen, a species of Heptapteridae, is considered to be a complex because of taxonomic and phylogenetic inconsistencies. Determining the physical location of repetitive DNA sequences on the chromosomes and the DNA barcode might increase our understanding of these inconsistencies within different groups of fish. To this end, we analyzed R. quelen populations from two river basins in Brazil, Paraguay and Parana, using DNA barcoding and different chromosomal markers, including U2 snDNA, which has never been analyzed for any Rhamdia species. Cytochrome c oxidase I gene sequence analysis revealed a significant differentiation among populations from the Miranda and Quexada rivers, with genetic distances compatible to those found among different species in neotropical fishes. Our results, in general, revealed a conservative chromosomal evolution in R. quelen and a differential distribution of some markers, such as 5S rDNA and U2 snDNA, in different populations. We suggest that R. quelen must undergo a major revision in its morphological, genetic, and cytogenetic molecular and taxonomic structure to elucidate possible operational taxonomic units.
Introduction
Rhamdia species are found in nearly all South American drainage basins and are the most specious genus in the Heptapteridae family. Rhamdia's body shape, color pattern, and habitat are very similar among species, 1 which makes it difficult to establish their taxonomic status. In a systematic review of species morphology conducted by Silfvergrip, 1 more than 100 species of Rhamdia were reclassified into 11 species and, later, Ferraris 2 reclassified these same species into 17 species, 50 species names of which were synonyms of Rhamdia quelen, which have often been analyzed from morphological, genetic, and cytological points of view. Most studies disagree with the above classifications, for example, genetic and morphometric evidence suggest that Rhamdia guatemalensis, Rhamdia saijaensis, and Rhamdia cinerascens, considered synonyms of R. quelen, should be considered valid species. 3 In addition, the morphological and karyotypic characteristics of Rhamdia voulezi and Rhamdia branneri have suggested that these are also valid species,4–6 and analyses of DNA barcoding, a genetic marker that is widely used to identify groups with taxonomic inconsistencies, confirmed the occurrence of these two species. 7
Chromosomal mapping of repetitive DNA, such as multigene families, satellites, and transposable elements, can also help resolve taxonomic problems, 8 phylogenetic problems, 9 populational differentiations, 10 sexual chromosome evolution, 11 and B chromosomes.12,13 As with a majority of other fish species, 18S rDNA is the most studied repetitive DNA class in R. quelen, and its distribution in a unique chromosome pair has proved to be well preserved at the species level (Table 1). 5S rDNA sequences also appear to be distributed within a unique chromosomal pair, in the interstitial position, in the majority of populations of R. quelen that have already been analyzed.5,10
Karyotypic Data in Rhamdia quelen
2n, diploid number; a, acrocentric chromosomes; Ar, Argentina; Bs, supernumerary chromosome; CMA3, fluorochromes chromomycin A3; FISH 5S, in situ hybridization with 5S rDNA probe; FISH 18S, in situ hybridization with 18S rDNA probe; Lg, lagoon; m, metacentric chromosomes; MS, Mato Grosso do Sul; FN, fundamental number; NOR, nucleolus organizer regions; PR, Paraná; R, river; Rb, stream; RJ, Rio de Janeiro; RS, Rio Grande do Sul; Rv, reservoir; SC, Santa Catarina; sm, submetacentric chromosomes; SP, São Paulo; st, subtelocentric chromosomes.
Little is known about the other repetitive DNA sequences in this group of fish, such as the snRNAs, noncoding RNAs that are part of the spliceosome machinery, can be subdivided into U1, U2, U4, U5, and U6 snRNAs, and play a crucial role in the splicing process of precursor mRNA. 14 Although scarce, data on the physical mapping of U2 snRNA genes in fish have shown a strong conservation in the number of sites per genome and suggested that they tend to accumulate exclusively within one chromosomal pair. 15
In this study, we associated chromosomal mapping of ribosomal and U2 snDNA genes with an analysis of the cytochrome oxidase I (COI) gene sequence in different populations of R. quelen from two hydrographic basins in Brazil. Because the taxonomic status of R. quelen is controversial, the objectives of these analyses were to observe a possible difference and recognize genetic divergence between these populations.
Materials and Methods
Origin of the samples
We analyzed six populations of R. quelen collected from different hydrographic basins in Brazil (Table 2). The samples were collected with the permission of the Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (IBAMA), protocol number 11399-1. We also obtained permission from the research ethics committee of the University Estadual of Londrina (Animal Ethical Use No.: CEUA 28520.2012.03).
Collection Sites and Hydrographic Basins of Rhamdia quelen Specimens Analyzed
Total of individuals: 52.
MS, Mato Grosso do Sul; PR, Paraná; ?, unidentified sex.
Conventional cytogenetic analysis and chromosome banding
Mitosis was stimulated by injection of a yeast suspension in the animals, as described by Lee and Elder. 16 Mitotic chromosomes were obtained by direct preparation by removing the anterior kidney, according to the methodology proposed by Bertollo et al. 17 Conventional analysis slides were stained with Giemsa 5% in phosphate-buffered solution (PBS) at pH 6.8. The chromosomes were classified according to the protocol of Levan et al. 18 and to determine their fundamental number (FN), the metacentric chromosomes (m), submetacentric chromosomes (sm), and subtelocentric chromosomes (st) were considered having two arms; the acrocentric chromosomes (a) were considered to have one arm. The nucleolus organizer regions (AgNORs) were detected using the silver nitrate impregnation technique according to the protocol of Howell and Black. 19 To determine the relative abundance of G–C and A–T base pairs, the chromosomes were stained with chromomycin A3 (CMA3) and 4,5-diamino-phenylindole (DAPI) fluorochromes, respectively, according to the methodology by Schweizer. 20
Preparation and labeling of probes for fluorescence in situ hybridization
Probes for 18S rDNA of Prochilodus argenteus (Hatanaka and Galetti 21 ) and U2 snRNA of Eigenmannia virescens used in the double fluorescence in situ hybridization (FISH) were labeled by polymerase chain reaction (PCR) with biotin-16-dUTP and the signal detected with avidin-FITC (Life Technologies, Carlsbad, CA), and were labeled with digoxigenin-11-dUTP and the signal detected with anti-digoxigenin-rhodamine (Roche Applied Science, Indianapolis, IN), respectively. The 5S rDNA probe in Imparfinis schubarti (Gouveia et al. 22 ) was labeled by PCR with the nucleotide digoxigenin-11-dUTP 1 mM, and the signal was detected with anti-digoxigenin-rhodamine (Roche Applied Science). FISH was conducted according to the methodology by Pinkel et al. 23
Fiber stretch for fiber-FISH
Cytogenetic preparations using a large number of interphase nuclei were conducted as described by de Barros et al. 24 Cell suspensions were dripped onto glass slides and washed in 1 × PBS for 2 min at room temperature (25°C). The slides were positioned horizontally, and 200 μL barium hydroxide solution (0.15 M NaOH) diluted in 30% alcohol (preprepared) was dripped onto the upper one. The slides were placed on a slope of 30°–40°, and a smear was created by sliding a clean blade over the slides, spreading the entire NaOH solution. Next, 500 μL 100% ethanol was spread over the entire slide, which remained in an inclined position until complete evaporation. The images were captured with a Moticam Pro 282B digital camera coupled with a fluorescence photomicroscope Leica DM 4500 B or an Olympus DP70 coupled with an Olympus BX61 photomicroscope.
Genetic analyses
Five individual fish from the Miranda River (MS) and three from the Quexada River (PR) were used for amplification of COI gene sequence (∼650 bp) using PCR with primers FishF1 and FishR2 (Ward et al. 25 ). PCR was conducted using the following cycle: initial denaturation at 94°C for 5 min; 35 cycles at 94°C for 30 s, 54°C for 30 s, and 72°C for 1 min, with a final extension at 72°C for 10 min in the MJ Research PTC-100. Each reaction was conducted in a final volume of 10 μL 1 × PCR Master Mix (Promega Corporation, Madison, WI) and 0.4 μM primers FishF1 and FishR1 with ∼15 ng DNA and water to yield a total volume. After amplification, the products were visualized in 1% agarose gel (2 μL sample loading buffer) and stained with SYBR® Safe DNA gel stain (Life Technologies).
Following gel analysis, the remaining amplified product was purified by adding 0.5 μL Illustra™ ExoProStar 1-Step™ (GE Healthcare, Little Chalfont, United Kingdom) following the manufacturer's instructions. The product was sent to sequencing, and the two strands were sequenced in independent reactions following the protocol recommendations described by Hajibabaei 26 for molecular identification. Sequencing was performed on an ABI 3500 automated sequencer XL (Applied Biosystems, Inc., CA).
The sequences obtained for both strands—forward and reverse—of ∼600 bp each were combined to form a consensus using the online application Electropherogram Quality Analysis (Togawa and Brigido 27 ). The strands were then submitted to the National Center for Biotechnology Information database (www.ncbi.nlm.nih.gov) under accession numbers KU845687–KU845694. BLAST software was used for similarity searching of the COI gene sequences in GenBank. Correspondences with similarities >98% with interplant distances of <2% were considered to be significant.
In addition, the COI gene sequences of R. quelen from adjacent basins, as well as that from Rhamdia laticauda (included as an outgroup), were retrieved from GenBank (Supplementary Table S1) and incorporated into the analysis of intraspecific and interspecific genetic distance based on Kimura's two-parameter model (K2P; Kimura 28 ). The confidence level of the branch configuration was assessed using the bootstrap method (1000 pseudoreplicates). The same samples were used to model and construct a genetic distance tree grouped by using the neighbor-joining (NJ) method to create a graphical representation of the distribution of genetic distances among populations. Analyses of the distance and NJ tree were conducted using MEGA v 6.06 (Tamura et al. 29 ).
Results
Classic cytogenetic analysis
All of the populations presented a diploid number equal to 58 chromosomes, with variations in the karyotypic formula as follows: 40m+10sm+4st+4a and FN equals 112 for the Quexada River (Fig. 1a), 40m+12sm +6st with FN equals 116 for the Penacho stream (Fig. 1b), 32m+8sm +18st with FN equals 116 for the Cambé River (Fig. 1c), and 34m+16sm +8 st with FN equals 116 for the Miranda River (Fig. 1d). The Taquari River and the Lindóia stream populations had been previously described by Moraes et al. 30 and Tsuda et al., 31 respectively, by means of conventional staining, silver nitrate impregnation, and fluorochrome staining.
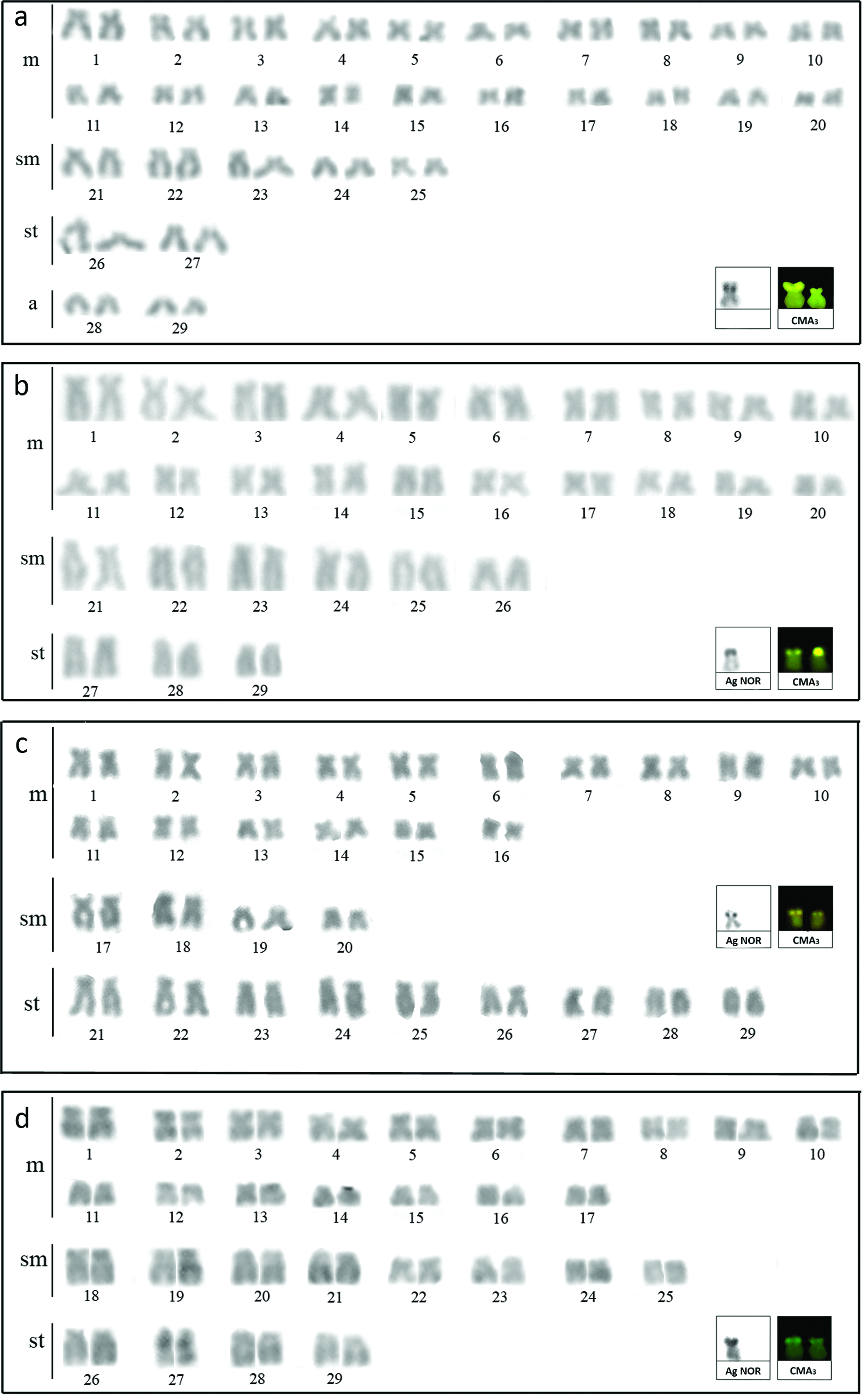
Karyotypes of Rhamdia quelen:
Silver nitrate impregnation revealed AgNORs at the terminal region of the short arm on only one homologous submetacentric chromosome of the four R. quelen populations (Fig. 1a–d, box). The CMA3 fluorochrome treatment revealed a positive signal in a submetacentric pair, most likely corresponding to AgNORs (Fig. 1a–d, box). We found a size heteromorphism among homologous chromosomes in the population from the Quexada River (Fig. 1a, box). We did not observe any DAPI markings.
Chromosome mapping with repetitive DNAs
FISH with an 18S rDNA probe also identified only one chromosomic pair with the ribosomal cistron in the terminal area in the six R. quelen populations (Fig. 2a–f). A size heteromorphism among the homologous chromosomes was also observed in the Quexada and Miranda River populations (Fig. 2a, c, respectively).
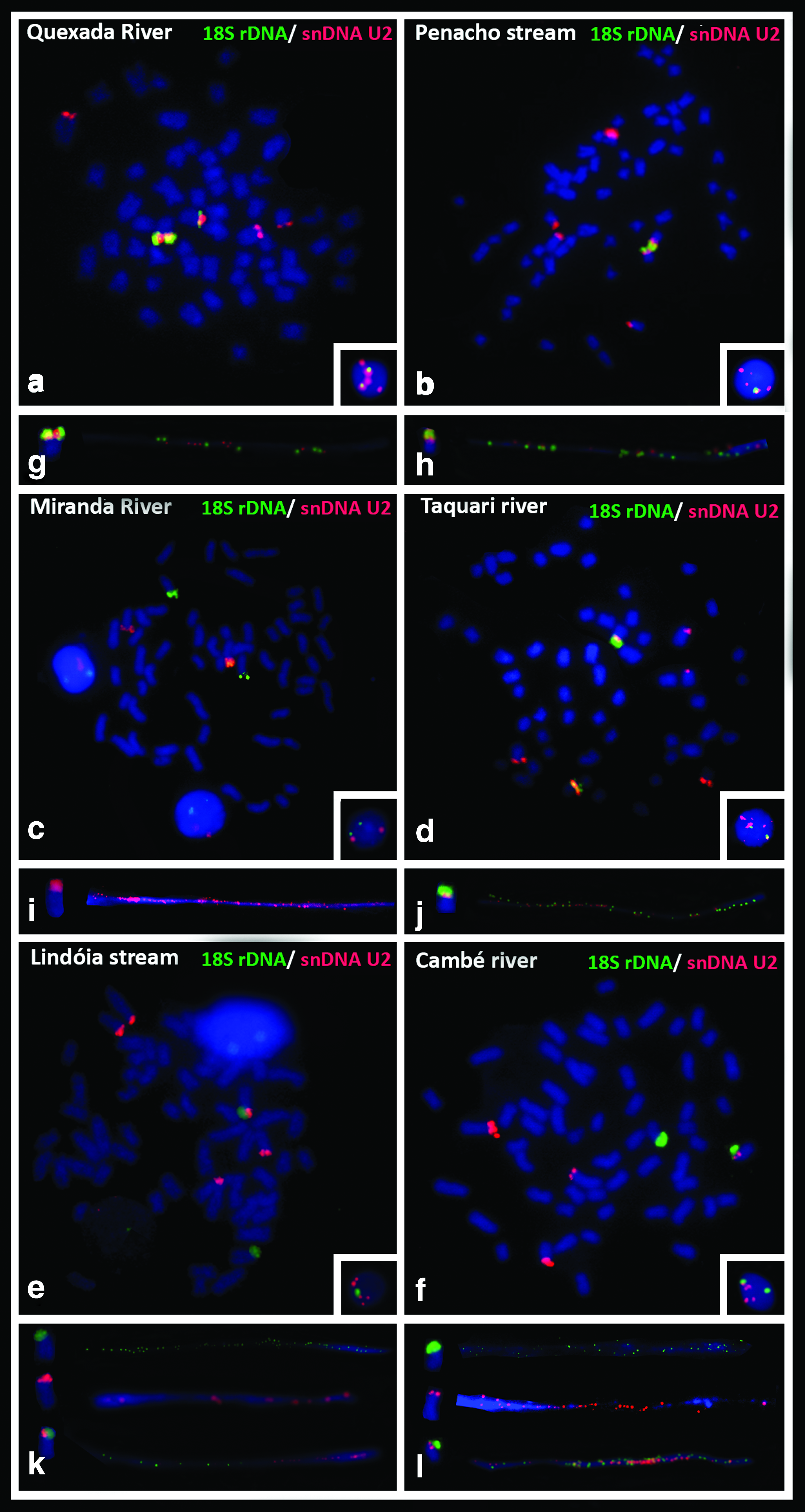
Somatic metaphases of R. quelen after double FISH with probes 18S rDNA (green) and U2 snDNA (red):
The U2 snDNA sequence was observed in four chromosomes in the Cambé River population (Fig. 2f), in five in the Quexada River and Lindóia stream populations (Fig. 2a, e, respectively), in six in the Penacho stream and Taquari River populations (Fig. 2b, d, respectively), and a unique pair in the Miranda River population (Fig. 2c).
The simultaneous detection of 18S rDNA with U2 snDNA that revealed three distinct patterns was confirmed by fiber-FISH in the R. quelen populations as follows: (i) gene synteny on a chromosomic pair in the Quexada River, Penacho stream, and Taquari River populations (Fig. 2a–g, b–h, and d–j, respectively), (ii) synteny on a unique chromosome in the Lindóia stream and Cambé River populations (Fig. 2e–k and f–l), and (iii) nonsyntenic position in the Miranda River population (Fig. 2c–i). Hybridizations of interphase nuclei were observed in the populations, which revealed the syntenic conditions of the 18S rDNA and U2 snDNA probes, and confirmed the number of clusters of each gene (Fig. 2, boxes).
The 5S rDNA sequence is located at the interstitial position in a chromosomic pair in the Quexada, Penacho, Taquari, Lindóia, and Cambé populations (Fig. 3a, b, d, and f). The Miranda River population was the only one to exhibit more than one chromosomic pair that contained this rDNA site (Fig. 3c).
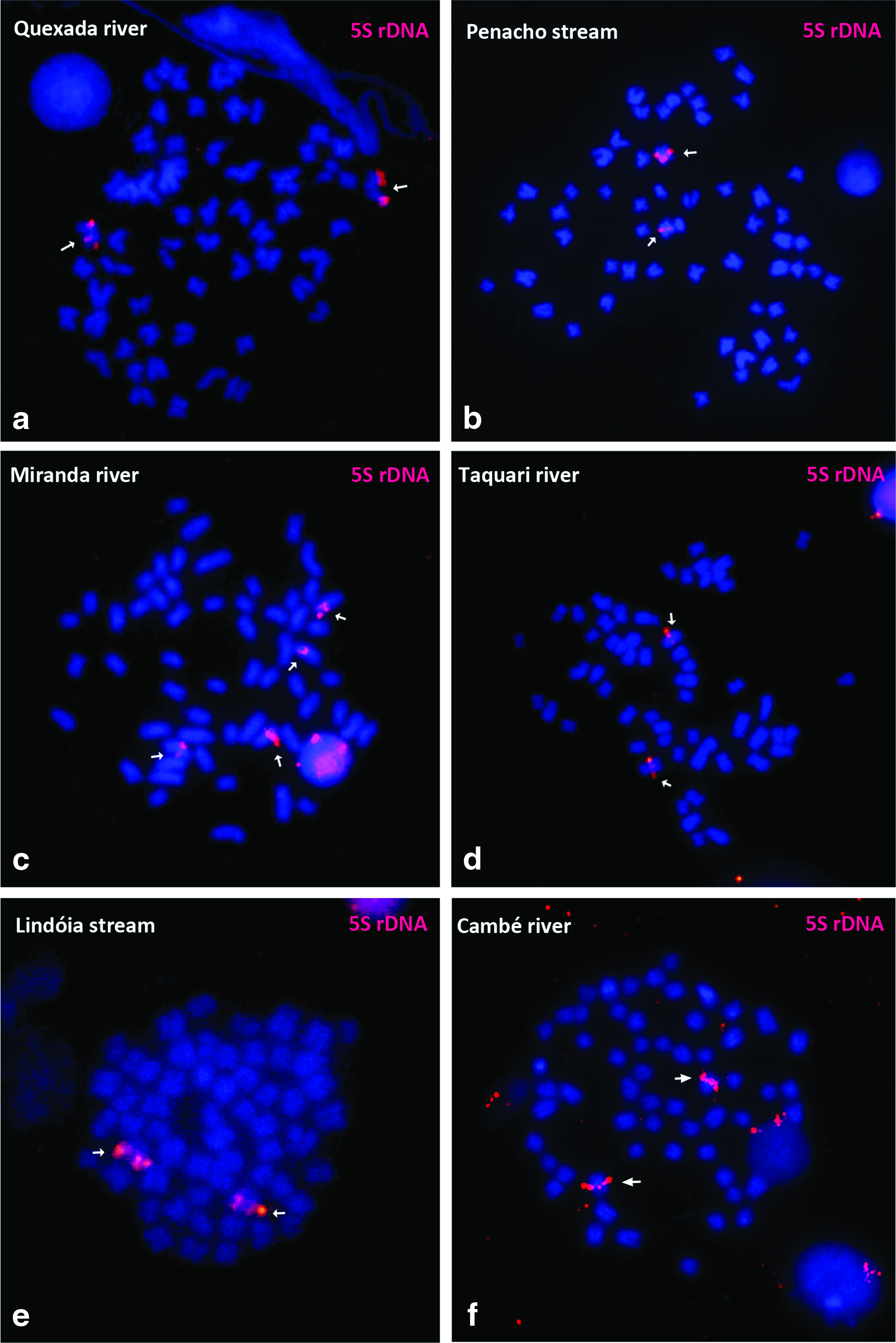
Somatic metaphases of R. quelen after FISH with 5S rDNA probe:
DNA barcoding
Five hundred fifty-one base pairs arose from COI gene sequencing of eight specimens. Including the GenBank sequences, a matrix of 718 bp was analyzed in which 85 bp were variable and 75 were parsimoniously informative, presenting a nucleotide frequency of 23.9% adenine, 29.6% thymine, 18.9% guanine, and 27.7% cytosine. No stop codon or signals of putative pseudogene were found. The K2P genetic distance analysis revealed a substantial genetic difference (5.9%) among the R. quelen samples from the Quexada and Miranda Rivers, which showed a very small intraspecific genetic distance (<0.1%) (Table 3). The same pattern was observed in the intraspecific and interspecific genetic distances among the COI gene sequences of R. quelen from different hydrographic basins, except in samples from the Upper Paraná and São Francisco River basins, which showed small interspecific genetic distances (Table 3). Also, the NJ-K2P tree revealed the placement of samples from the Miranda River into an isolated group and confirmed the differences between those from the Quexada and Miranda Rivers and their respective watersheds with strong bootstrap support (≥70%; Fig. 4).
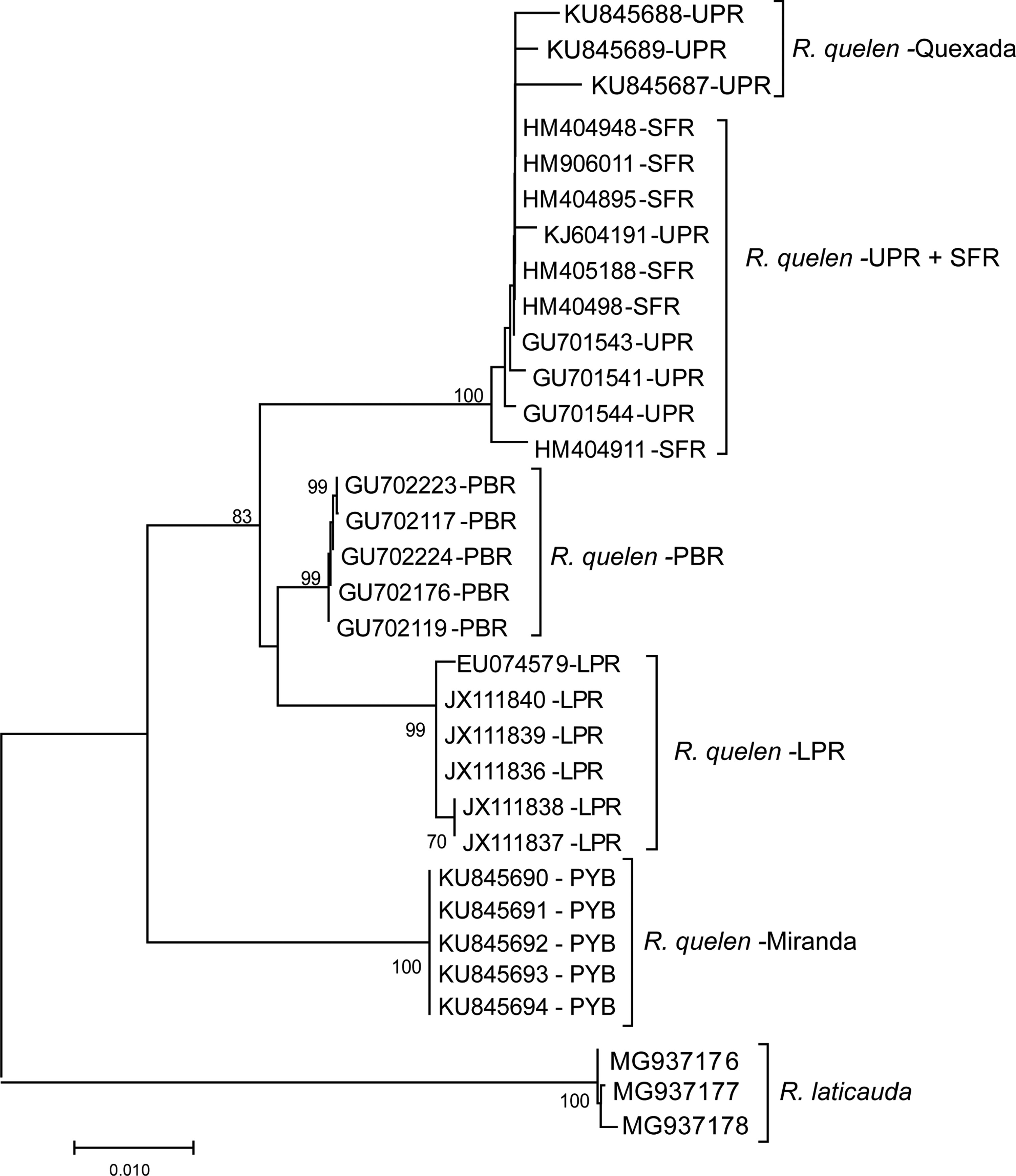
Graphical representation of Kimura's two-parameter genetic distance estimated through neighbor-joining method for the sequences of R. quelen from the PYB (Miranda river—MS), UPR (Quexada river—PR), LPR, SFR, PSR, and Rhamdia laticauda as outgroup. Brach supports were calculated based on 1000 bootstraps and values below 70% were omitted. LPR, Lower Paraná river basin; PSR, Paraíba do Sul river basin; PYB, Paraguay river basin; SFR, São Francisco river basin; UPR, Upper Paraná river basin.
Kimura's Two-Parameter Model Genetic Distance Among Samples of Rhamdia quelen and Rhamdia laticauda from Different South American Basins
Interspecific K2P genetic distance is represented in lower diagonal with respective standard deviation in the upper diagonal. The values of intraspecific genetic distances are reported in bold along the diagonal.
Hydrographic basins are abbreviated as follows: LPR, Lower Paraná river basin; PBR, Paraiba do Sul river basin; PYB, Paraguay river basin; SFR, São Francisco river basin; UPR, Upper Paraná river basin.
K2P, Kimura's two-parameter model.
Discussion
From a cytogenetic point of view, R. quelen is the most studied species in the Heptapteridae family, presenting 2n = 58, well-conserved at genus, a predominance of meta and submetacentric chromosomes, with high FNs, and a distinct karyotypic formula, as evidenced in this study (Table 1). Martinez et al. 10 proposed that different chromosomal rearrangements created this structural variability with a particular geographical distribution. The same authors suggested that this group represents a species complex that requires taxonomic revision. Recently, Rios et al. 32 proposed that R. quelen be split into seven highly differentiated genetic lines, which is consistent with the species complex hypothesis and suggests the existence of cryptic species.
A single NOR in submetacentric and subtelocentric chromosomes is the most frequent feature in R. quelen and was confirmed by FISH with the DNAr 18S probe in some populations, including the ones studied here (Table 1). This pattern is an important cytogenetic marker for the species because multiple systems have already been described for other Rhamdia species.4,33 According to Salvadori et al., 34 this region is associated with DNA rich in GC basis pairs, a pattern observed in all the R. quelen populations analyzed (Table 1). A structural polymorphism of the ribosomal site may be the cause of the NOR size polymorphism of the Quexada and Miranda rivers, and the possible causes of this polymorphism include spontaneous duplication events, deletion, and unequal cross-over.35–38
Most studies of the distribution of U2 snDNA sites have suggested that they accumulate in an exclusive chromosomal pair. 15 We demonstrated differences in U2 snDNA accumulation between populations of the same species, and individuals from the Miranda River were the only ones that exhibited only one chromosomal pair with this sequence. The variability observed among the six populations demonstrates that this sequence may be one more important cytogenetic marker in R. quelen because we demonstrated an interpopulational variation never before analyzed in other species. The presence of more sites of U2 snDNA could be related to an association of this sequence with transposons (TEs) and retrotransposons (TREs); however, until now, no study has analyzed this possiblity. TEs and TREs have been observed associated with 18S rDNA, in Sorubim lima 39 and Astyanax scabripinnis, 40 and 5S rDNA, in Eigenmannia.41,42 According to Le Rouzic and Capy, 43 this association may favor the occurence of the recombination of this DNA carrying mutations that may influence not only the genome organization but also the expression of those sequences.
The association of snDNA U2 with other repetitive DNAs was also found in other species; the syntenic location of 5S rDNA and U2 snDNA was observed in 10 species of Characidium, 44 four species of Triportheus, 15 and Eigenmannia aff. trilineata. 45 The co-location of 18S and U2 snDNA has also been observed in Thalassophryne maculosa 46 and Bryconamericus ecai cytotype VI 47 ; the syntenic location of 18S and U2 snDNA was observed only in Triportheus albus, Triportheus auritus, Triportheus nematurus, Triportheus signatus, and Triportheus trifurcatus 15 and in this study.
The absence of U2 snDNA sites in one of the homologous chromosomes in the Quexada and Cambé river and the Lindóia stream populations could be explained due to a size heteromorphism of this site, which is impossible to detect using FISH. However, fiber-FISH, which is a powerful approach to high-resolution physical mapping, confirmed this result. In this way, chromosome orientation according to the Rabl model can explain this situation because this site is located in the terminal area of the chromosomes. Therefore, transfer of genetic material might be occuring from one homologous chromosome to another due to proximity to the interphase nucleus. 48
5S rDNA is excellent tool for studying the evolution of multigene families because it acts as a genetic marker among species and populations. 49 Among the Siluriformes, there is a high variability in the number of 5S ribosomal clusters, 50 and R. quelen is one of the few species in which only one chromosome pair bears this cistron (Table 2). The exception in this study was the Miranda River population. It was the most different of the six, with two bearing pairs of this region, which might be a result of translocation, as proposed by Garcia et al. 5 for R. quelen in the Fortuna River, which also presented multiple 5S rDNA sites.
According to Martins and Galleti, 51 the location of this sequence at the interstitial region may be responsible for protection against transposition and crossing events, which are more frequent in terminal regions. Other phenomena may be acting to maintain this sequence at this site since in R. quelen, as in other species, the 18S rDNA sequence is maintained in the terminal position of a chromosomal pair even though it is more subject to transposition and crossing events.
The pattern of differentiation obrserved for the cytogenetic markers was corroborated by COI analysis. For all comparisons, the samples from Miranda river showed genetic distances fourfold or fivefold higher than the mean of genetic distances registered for the species in the Upper Parana river basin, 52 and 40- or 60-folds higher than intraspecific genetic distances observed for the samples from the hydrographic basins analyzed. Interestingly, each hydrographic basin showed patterns of genetic distance in line with the expected for different species,53–55 which highlight the isolation among these watersheds, excepting the São Francisco and Upper Paraná river basins. Despite us analyzing two populations from Paraguai and Upper Paraná river basins, the comparison among these and those samples from GenBank point out to the taxonomic status of R. quelen throughout different watersheds.
This study showed a karyotypic conservative structure in R. quelen, of diploid number and rDNA number of sites, already identified as a characteristic of the species. However, the inclusion of U2 snDNA data revealed an important variability, pointing out these sequences as suitable markers for populations, in agreement with that observed in COI analyzes. These results demonstrate the power of joint cytogenetic analysis with DNA barcoding for identifying taxonomic distances, which are candidates for new species, reinforcing the idea that R. quelen may be a species complex, as suggested by Martinez et al. 10 and Ríos et al., 32 and corroborating a need for a major revision in this genus based on morphological, genetic, and cytogenetic data.
Footnotes
Acknowledgments
The authors thank Dra. Fernanda Simões de Almeida and Dra. Silvia Helena Sofia of the Genetics and Animal Ecology Laboratory (LAGEA) for allowing us to use their laboratory for DNA barcoding experiments. This study was supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Brazil.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.