
Abstract
Abstract
Fibrodysplasia Ossificans Progressiva (FOP) is a rare, autosomal dominant genetic disorder in humans characterized by the gradual ossification of fibrous tissues, including skeletal muscle, tendons, and ligaments. In humans, mutations in the Type I BMP/TGFβ family member receptor gene, ACVR1, are associated with FOP. Zebrafish acvr1l, previously known as alk8, is the functional ortholog of human ACVR1. We previously created and characterized the first adult zebrafish model for FOP by generating animals harboring heat shock-inducible mCherry-tagged constitutively active Acvr1l (Q204D). Since injury is a known trigger for heterotopic ossification (HO) development in human FOP patients, in this study, we investigated several injury models in Acvr1lQ204D-expressing zebrafish and the subsequent formation of HO. We performed studies of Activin A injection, cardiotoxin (CTX) injection, and caudal fin clip injury. We found that none of these methods resulted in HO formation at the site of injury. However, some of the cardiotoxin-injected and caudal fin-clipped animals did exhibit HO at distant sites, including the body cavity and along the spine. We describe these results in the context of new and exciting reports on FOP, and discuss future studies to better understand the etiology and progression of this disease.
Introduction
F
In general, the progression of an FOP lesion is currently thought to follow a two-phase process. 7 In the first phase, inflammation results from a triggering event, which can include a physical injury, a surgical procedure, or a systemic infection.4,8,9 Inflammatory cells overwhelm the lesion site and promote muscle tissue degradation, angiogenesis, and fibroproliferation.10–12 In the second phase, a recruited population of fibro/adipogenic progenitor cells (FAPs) then undergoes the process of normal endochondral ossification to generate HO at the site of the FOP lesion.7,13–15
Given the intriguing finding that immunosuppression was able to mitigate the development of HO in a human FOP patient, 16 it is possible that the immune system may play a greater role in driving early stage FOP lesion and HO formation than currently understood. Thus, a better understanding of the inflammatory response in FOP patients is needed and may help in developing therapeutic strategies to reduce and/or abolish FOP lesion formation. To address this and other questions in the FOP field, researchers have developed a number of FOP animal injury models that induce HO in a consistent and site-directed manner.13–15,17–24 These valuable models have provided significant insight into the mechanism of FOP-induced HO, including the important finding that ACVR1R206H is activated by activin A rather than inhibited by it, as in the wild-type ACVR1,20,22 that ACVR1R206H expressing FAPs are cells contributing to HO in FOP patients,13–15 and that enhanced mTOR signaling promotes endochondral ossification in FOP. 21 These models have also been invaluable in validating possible new therapeutic strategies to treat FOP.17,18,21 In light of these advances, we felt that the generation of an adult zebrafish model of FOP, with all of the advantages that the zebrafish system provides (reviewed in Ref. 25 ), would be an excellent tool in which to further probe open questions in the field.
In our prior work, we developed the first adult zebrafish model for FOP. 26 We used Tol2 transgenesis to generate transgenic zebrafish carrying a heat shock-inducible, mCherry-tagged, constitutively active form of acvr1l, the functional zebrafish ortholog of human ACVR1. We exposed these Acvr1lQ204D-expressing animals to several months of daily heat shock to investigate whether constitutively active Acvr1l would promote the formation of FOP-like phenotypes, including HO. We found that adult zebrafish ubiquitously express Acvr1lQ204D when exposed to long-term daily heat shock and develop FOP-like characteristics, including fully penetrant abnormal spinal curvature, and partially penetrant vertebral fusions, pelvic fin malformations, and HO.
In this study, we expand upon our work with the Tg(acvr1l_Q204D) zebrafish using a series of injury models to investigate whether Acvr1lQ204D-expressing zebrafish develop reliable site-directed HO lesions. We found that injury of Acvr1lQ204D-expressing animals by any one of three methods, (1) injection of recombinant human (rh) activin A, (2) injection of cardiotoxin (CTX), or (3) clipping of the caudal fin, resulted in early tissue damage that resolved normally and did not result in HO at the site of injury. Interestingly, several Acvr1lQ204D-expressing animals, injured either by CTX injection or caudal fin clip, did develop HO lesions at sites distal to the injury site, including the body cavity or along the spine. In this study, we describe the development and characterization of each of these injury models and discuss our future plans to create more robust zebrafish models for human FOP.
Materials and Methods
Zebrafish husbandry
Zebrafish (Danio rerio) were raised in the Tufts Yelick Lab Zebrafish Facility at 28.5°C in a controlled, automated recirculating environment, with 14/10 h light/dark cycle, as previously described. 27 Zebrafish of both sexes were used in this study, at ages indicated in the article. We used the following mutant and transgenic strains: Tg(Bre:GFP) 28 and Tg(Bre:GFP); Tg(hsp70l:acvr1l_Q204D-mCherry). 26 The following shorthand names are used in this article: Tg(hsp70l:acvr1l_Q204D-mCherry) is referred to as Tg(acvr1l_Q204D), protein product as Acvr1lQ204D.
Ethics statement
All experimental procedures on zebrafish embryos, larvae, juveniles, and adults were approved by the Tufts University Institutional Animal Care and Use Committee (IACUC) and Ethics Committee.
Heat shock procedures
Adult Tg(Bre:GFP) and Tg(Bre:GFP); Tg(acvr1l_Q204D) zebrafish were heat-shocked once daily for 1 h at 38°C, as previously described. 26 Animals were maintained on heat shock systems throughout the duration of each injury experiment. At collection time points, animals were euthanized using 250 mg/L tricaine methanesulfonate (Argent Aquaculture, Redmond, WA) and fixed before microcomputed tomography (Micro-CT) analysis or processing for histological and/or immunohistochemical analyses.
Injury methods
Zebrafish were injured using one of the following three methods: (1) rh activin A injection; (2) CTX injection; or (3) caudal fin clip. For the rh activin A injury experiments, zebrafish were anesthetized with 150 mg/L tricaine methanesulfonate. Blunt forceps were used to gently remove scales at the injury site (on the right side, above the spine, just anterior to the dorsal fin). The rh activin A (0.1 mg/mL in dimethyl sulfoxide [DMSO], 2 μL per injection; 338-AC, R&D Systems, Minneapolis, MN) was then injected at the site of scale removal using a 100 μm diameter microcapillary needle and FemtoJet microinjection station (Eppendorf, Hamburg, Germany). Control animals were injected with DMSO alone. After injection, animals were allowed to recover in fresh system water and then returned to their respective heat-shocked tanks for continued daily heat shock. Tg(Bre:GFP) and Tg(Bre:GFP); Tg(acvr1l_Q204D) zebrafish were heat-shocked starting at 14 days post-fertilization (dpf) for 8 months before rh activin A injections and harvested at 1 h postinjury (hpi), 2 days postinjury (dpi), and 4 weeks postinjury (wpi). All specimens were prepared and analyzed using histology and immunohistochemistry (IHC).
The CTX injury experiments were carried out essentially as described above for the rh activin A injury experiments. The injury site was descaled and injected with CTX from naja pallida (0.3 mg/mL in phosphate-buffered saline [PBS], 1 μL per injection; 217503, Millipore, Burlington, MA) as per prior published reports in zebrafish. 29 Control animals were injected with PBS alone. Tg(Bre:GFP) and Tg(Bre:GFP); Tg(acvr1l_Q204D) zebrafish were heat-shocked starting at 14 dpf for 10 months before CTX injections. CTX-injected zebrafish were collected at 2 dpi, and at 4 and 8 wpi. Collected specimens were analyzed by micro-CT (8 wpi only), histology, and IHC.
For caudal fin clip injury, ∼50% of the caudal fin was removed from anesthetized zebrafish using a sterile razor blade. Bright field images before and after fin clip tails were captured using an Axiocam 503 color camera (Zeiss, Oberkochen, Germany) attached to an M2-Bio dissecting microscope (Zeiss) and processed using AxioVision SE64 microscopy software (Zeiss). Tg(Bre:GFP) and Tg(Bre:GFP); Tg(acvr1l_Q204D) zebrafish were heat-shocked starting at 14 dpf for 7 months before fin clip injury. Fin clip-injured zebrafish were all collected at 2 wpi and analyzed by Micro-CT, histology, and IHC.
Histology and immunohistochemistry
Two different methods of fixation, tissue processing, and paraffin embedding were used for zebrafish injury experiments. For the rh-activin A and CTX injury experiments, zebrafish were processed as previously described by us. 25
For the tail clip injury experiments, animals were processed as previously described with the following modifications 29 : briefly, zebrafish were fixed in 10% formalin in PBS for 48 h at room temperature (RT), and then rinsed and stored in PBS at 4°C. For paraffin embedding, samples were decalcified in CalEx (Fisher Scientific, Pittsburgh, PA) for 24 h and rinsed in PBS for 2 h, before dehydration in EtOH series. Samples were transferred to 65°C molten Paraplast Plus paraffin (McCormick Scientific, St. Louis, MO) overnight, transferred to fresh molten paraffin, and allowed to harden at RT. Paraffin blocks were serially sectioned at 7 μm and mounted on SuperFrost Plus charged glass microscope slides (Fisher Scientific).
For histological analyses, mounted sections were stained with H&E or with Safranin O and Fast Green counter stain. For Safranin O staining, slides were submerged in 0.02% Fast Green in 0.2% acetic acid for 15 s, followed directly by submersion in 1% Safranin O for 30 s. IHC analyses were performed as previously described 26 using the following antibodies: 1:200 goat anti-activin A (AF338; R&D Systems); 1:100 goat anti-Collagen I (AB758; Millipore); 1:200 rabbit anti-Collagen II (ab34712; Abcam, Cambridge, MA); 1:250 rabbit anti-pSmad2 (AB3849-I; Millipore); 1:50 rabbit anti-von Willebrand factor (vWF) (HPA001815; Sigma-Aldrich, St. Louis, MO); 1:500 donkey anti-rabbit and 1:500 donkey anti-goat (Jackson ImmunoResearch Laboratories, West Grove, PA). Stained sections were analyzed and imaged using a Zeiss AxioPhot epifluorescence microscope (Zeiss) and Axiocam 503 color camera (Zeiss) and processed using AxioVision SE64 microscopy software (Zeiss).
Microcomputed tomography
Euthanized zebrafish were imaged using a Skyscan 1176 high-resolution micro-CT Scanner (Bruker, Allentown, PA) as previously described by us. 26 Scans were performed at 9 μm resolution, every 0.5° over 180°. Reconstructions were performed using NRecon (Bruker) and 3D renderings were generating using CTVox (Bruker). All reconstruction and rendering settings were kept constant between samples to allow for comparison.
Methodology and statistics
All animals established on a given heat shock system that were part of injury experiments were included in analyses described below. Animal numbers are indicated as per figure legends. Animals that died of natural causes before the desired collection times were excluded from analyses.
Results
rh activin A injection of heat-shocked Tg(acvr1l_Q204D) zebrafish does not induce HO at the injury site
To study the progression of HO formation in the context of FOP, researchers have created a variety of FOP mouse injury models.13,14,17–22,24,30 Based on these reports, we used similar approaches to create Acvr1lQ204D zebrafish injury models, to induce and study HO formation in these animals.
Activin A was recently identified as a critical ligand for ACVR1R206H-driven HO formation in an FOP mouse model. 20 We therefore chose to test whether an injection injury model that also delivers rh activin A resulted in HO formation in our Acvr1lQ204D-expressing zebrafish. As there are no existing protocols for injecting into adult zebrafish, we developed the following approach.
Briefly, adult zebrafish were anesthetized and scales were gently removed from the injection site (Fig. 1A). We then injected 2 μL of either 0.1 mg/mL rh activin A or DMSO vehicle control into the muscle tissue using a microcapillary needle (Fig. 1B). Successful delivery of rh activin A was confirmed by IHC analyses of injected animals at 1 hpi (Fig. 1C–H), which showed rh activin A protein in the muscle tissue of rh activin A-injected animals (Fig. 1E, H), but not in DMSO-injected animals (Fig. 1D, G), or no primary antibody negative controls (Fig. 1C, F).

rh activin A is detectable in zebrafish muscle tissue at 1 hpi. Before injection injury, adult zebrafish were anesthetized, and scales were gently removed from the right, dorsal side, just anterior to the dorsal fin
After confirming the effective delivery of rh activin A to zebrafish muscle tissue, we proceeded to perform rh activin A injections on adult heat-shocked Tg(Bre:GFP) and Tg(Bre:GFP); Tg(acvr1l_Q204D) zebrafish. Animals were collected at 2 dpi and 4 wpi and each injection site was analyzed using H&E staining and IHC (Fig. 2). The 2 dpi time point was chosen to assess muscle tissue damage at the site of injury, while the 4 wpi time point was intended to assess HO formation, based on the fact that HO typically develops within 3 weeks of injury in FOP mouse models.20,31,32 At 2 dpi, all animals displayed clear tissue damage at the injury site in response to either rh activin A or DMSO injection (Fig. 2A–D, damage denoted by dashed lines). Nuclear pSmad2 expression was used to demonstrate activated TGFβ signaling in all injected zebrafish (Fig. 2E–H), as expected in response to injury.33–35 By 4 wpi, heat-shocked Tg(Bre:GFP) and Tg(Bre:GFP); Tg(acvr1l_Q204D) zebrafish showed normal healing and resolution of the tissue damage (Fig. 2I–L), regardless of the injected compound. These results indicated that injury and delivery of rh activin A did not promote HO formation in Acvr1lQ204D-expressing zebrafish, consistent with recently published reports of the lack of responsiveness of the Q204D variant to activin A.

rh activin A injection caused tissue damage apparent at 2 dpi that resolved by 4 wpi. Adult heat-shocked Tg(Bre:GFP) and Tg(Bre:GFP);Tg(HS-acvr1l_Q204D-mCherry) zebrafish were injected with rh Activin A
CTX injection does not result HO formation at the injury site in heat-shocked Tg(acvr1l_Q204D) zebrafish
We next tested whether the CTX injection injury model could induce HO formation in Acvr1lQ204D-expressing zebrafish. CTX injection was previously used to study muscle regeneration in zebrafish 29 and in mouse models of FOP.13,14,18,19,21,24,30 CTX is a myonecrotic agent that induces severe muscle fiber contraction leading to cell death. 36 We performed CTX injections as described for rh activin A injections, in both heat-shocked Tg(Bre:GFP) and Tg(Bre:GFP); Tg(acvr1l_Q204D) zebrafish, using 1 μL of 0.3 mg/mL CTX or PBS vehicle control. Injected zebrafish were collected at 2 dpi and 4 wpi, and each injection site was analyzed using histological and IHC analyses (Fig. 3A–L). At 2 dpi, all injected zebrafish exhibited tissue damage at the injection site (Fig. 3A–D, damage denoted by dashed lines), similar to that observed in rh activin A injection injuries (Fig. 2). Notably, the extent of tissue damage in CTX-injected Tg(Bre:GFP) and Tg(Bre:GFP); Tg(acvr1l_Q204D) zebrafish (Fig. 3B, D) was larger than that observed in PBS control-injected zebrafish (Fig. 3A, C). pSmad2 expression confirmed activation of TGFβ signaling in all injected animals, with no obvious differences in pSmad2 levels between treatment groups (Fig. 3E–H). By 4 wpi, CTX injection-induced tissue damage appeared to be resolved, as normal muscle tissue was observed in injured Tg(Bre:GFP) and Tg(Bre:GFP); Tg(acvr1l_Q204D) zebrafish (Fig. 3I, J). Safranin O staining revealed no indication of cartilaginous proteoglycans, a hallmark of FOP-associated HO lesions, at the injury sites of zebrafish of either genotype (Fig. 3K, L). These results suggested that Acvr1lQ204D-expressing zebrafish did not form HO at the site of CTX injury.

CTX injection caused tissue damage apparent at 2 dpi that resolved by 4 wpi. Adult heat-shocked Tg(Bre:GFP) and Tg(Bre:GFP); Tg(HS-acvr1l_Q204D-mCherry) zebrafish injected with CTX
CTX-injured Acvr1lQ204D-expressing zebrafish exhibited body cavity HO
A cohort of CTX-injured Tg(Bre:GFP) and Tg(Bre:GFP); Tg(acvr1l_Q204D) zebrafish was collected at 8 wpi and analyzed by Micro-CT to detect skeletal defects and HO (Fig. 4A–D). Of the three Acvr1lQ204D-expressing zebrafish analyzed at 8 wpi, one displayed densely mineralized HO lesions within the body cavity (arrows in Fig. 4C, D), similar to the HO lesions observed in uninjured Tg(Bre:GFP); Tg(acvr1l_Q204D) zebrafish. 26 None of the heat-shocked Tg(Bre:GFP) zebrafish examined exhibited any detectable HO (Fig. 4A, B). Histological analyses of the mineralized masses in this Acvr1lQ204D-expressing animal revealed a heterogeneous HO lesion (Fig. 4E, F), and Safranin O staining was used to confirm large areas of cartilage matrix deposition within the lesion (Fig. 4G, H).

CTX -injected heat-shocked Tg(acvr1l_Q204D) zebrafish exhibited body cavity HO at 8 wpi. Micro-CT imaging of CTX-injected heat-shocked Tg(Bre:GFP); Tg(acvr1l_Q204D-mCherry) zebrafish
Caudal fin clip injury did not induce HO formation at the injury site of Acvr1lQ204D-expressing zebrafish
For our third and final injury model, we turned to the well-established zebrafish injury method, the caudal fin clip. 27 First, anesthetized Tg(Bre:GFP) and Tg(Bre:GFP); Tg(acvr1l_Q204D) zebrafish were imaged using bright field microscopy before caudal fin clip (Fig. 5A, B). Next, a sterile razorblade was used to remove ∼50% of the caudal fin of each animal (Fig. 5C, D). All zebrafish were subsequently monitored for fin regeneration at 1 and 2 wpi (Fig. 5E, F). Gross inspection by bright field imaging showed that all of the tail clipped Tg(Bre:GFP) and Tg(Bre:GFP); Tg(acvr1l_Q204D) zebrafish exhibited normal caudal fin regeneration by 2 wpi (Fig. 5G, H). Micro-CT analyses of zebrafish at 2 wpi revealed no signs of HO formation at the injury site (Fig. 5I, J).

Caudal fin clip did not result in HO formation at injury site in heat-shocked Tg(acvr1l_Q204D) zebrafish by 2 wpi. Caudal fin clip-injured adult heat-shocked Tg(Bre:GFP) and Tg(Bre:GFP); Tg(acvr1l_Q204D-mCherry) zebrafish were imaged with bright field microscopy before fin clip
HO formation was observed along the spine and in the body cavity of caudal fin clip-injured Acvr1lQ204D-expressing zebrafish
Careful inspection of micro-CT reconstructions of caudal fin clip-injured, heat-shocked Tg(Bre:GFP) and Tg(Bre:GFP); Tg(acvr1l_Q204D) zebrafish revealed the presence of numerous HO lesions in two of the three Acvr1lQ204D-expressing zebrafish (Fig. 6D–I). One Acvr1lQ204D-expressing zebrafish exhibited small HO lesions both within the body cavity and in rod-like HO structures that had formed adjacent to the spine (arrows in Fig. 6D–F). The other Acvr1lQ204D-expressing zebrafish exhibited a single large HO lesion within the body cavity (arrows in Fig. 6G–I). None of the Tg(Bre:GFP) zebrafish showed any signs of HO formation (Fig. 6A–C).
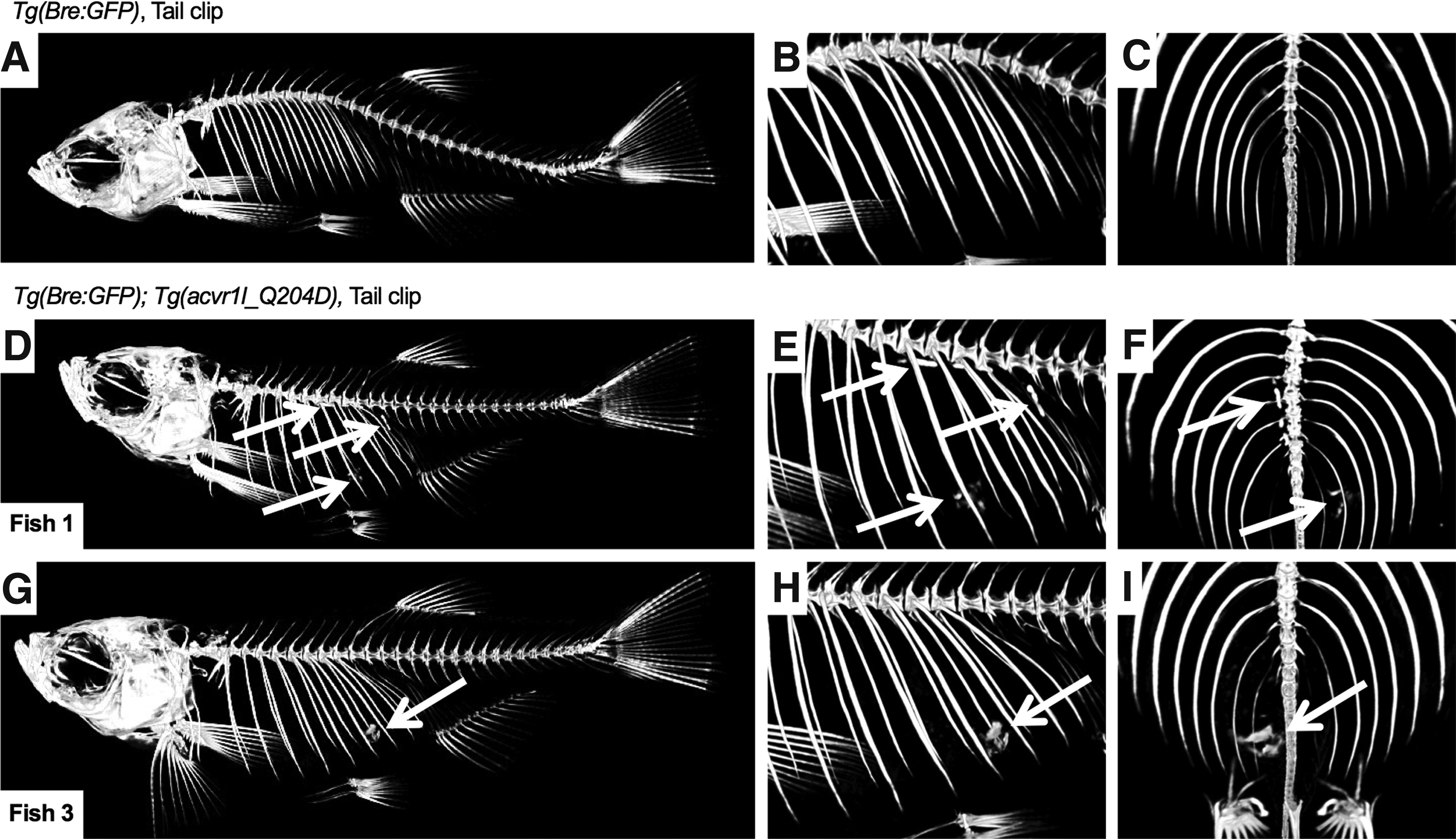
Caudal fin clip-injured heat-shocked Tg(acvr1l_Q204D) zebrafish exhibited spinal and body cavity HO at 2 wpi. Micro-CT image of tail fin clip-injured HS Tg(Bre:GFP); Tg(acvr1l_Q204D-mCherry) zebrafish
The Acvr1lQ204D-expressing zebrafish that displayed HO lesions, which were identified by micro-CT, were subsequently paraffin sectioned and analyzed by H&E staining (Fig. 7A, B, F, G) and IHC (Fig. 7C–E, H–J). H&E staining revealed that the HO lesions in Acvr1lQ204D-expressing animals were quite large (Fig. 7A, F, lesion denoted by dashed lines) and heterogeneous (Fig. 7B, G). IHC analyses of the HO lesions confirmed the presence of several cell types commonly found in HO, including vWF-expressing endothelial cells (Fig. 7C, H), collagen II-expressing cartilage (Fig. 7D, I), and collagen I-expressing osteoblasts (Fig. 7E, J). Interestingly, each marker exhibited different expression patterns throughout the HO lesion, indicative of various stages of HO progression within the lesion.

Cartilage and bone formation in body cavity HO in Acvr1lQ204D-expressing zebrafish. HO lesions in tail fin clip-injured HS Tg(Bre:GFP); Tg(acvr1l_Q204D-mCherry) zebrafish at 2 wpi, paraffin sectioned and H&E stained
Discussion
In this study, we describe the development and characterization of three injury methods used to induce HO in the previously characterized Tg(acvr1l_Q204D) zebrafish FOP model. 26 Prior characterization of this zebrafish FOP model showed that 1 h/day heat shock of Tg(hsp70l:acvr1l_Q204D-mCherry) zebrafish was sufficient to induce ubiquitous expression of Acvr1l Q204D for up to 3 days.
Acvr1lQ204D-expressing zebrafish injured by rh activin A injection, CTX injection, or caudal fin clip showed early signs of tissue damage, but none exhibited HO at the site of injury. Acvr1lQ204D-expressing zebrafish did exhibit HO at sites distal to the injury, either within the body cavity or along the spine. These HO lesions observed in injured Acvr1lQ204D-expressing zebrafish resembled those observed in uninjured Acvr1lQ204D-expressing zebrafish, although they were larger in size. It is important to note that our sample sizes for each experiment were small due to the complexity and duration of each experiment. Nonetheless, these results suggest that while Acvr1lQ204D-expressing zebrafish acquire phenotypes consistent with human FOP, they do not exhibit reliable HO formation at the site of injury. Additional animals can be tested in future experiments to provide further support for these findings.
We first determined that Acvr1lQ204D-expressing zebrafish did not form HO at the injury site in response to rh activin A injection. There are several possible explanations for this finding. It is possible that rh activin A cannot bind to the zebrafish Acvr1l receptor as it does to mouse and human ACVR1. Full-length zebrafish Acvr1l is 69% identical to human ACVR1 at the amino acid level, including an intracellular serine/threonine kinase domain that is 85% identical, strongly suggesting the conservation of receptor function in intracellular signaling cascades.25,37 However, the extracellular ligand binding domain of zebrafish Acvr1l is only 30.3% conserved compared to human ACVR1, 37 indicating that human Activin A may not bind to zebrafish Acvr1l (the mouse and human extracellular domains are 96% identical). In addition, the zebrafish inhibin βA subunits that dimerize to form activin A are only 55% identical to the human inhibin βA subunits. Consistent with this, we did not detect any endogenous zebrafish activin A expression when we performed IHC using the anti-rh activin A antibody (Fig. 1D, G). It is therefore possible that zebrafish Acvr1l is not capable of binding and responding to injected rh activin A. Another possible explanation is that zebrafish Acvr1lQ204D is not responsive to activin A in the same manner as is human ACVR1R206H. Previous work has shown that the ACVR1 p.Q207D mutation is ligand independent. 38 Indeed, recent work has suggested that the p.Q207D mutation is unaffected by activin A binding, in that activin A can neither suppress nor stimulate further activity of ACVR1Q207D. 39 Therefore, it is also possible that zebrafish Acvr1lQ204D is similarly unresponsive to rh activin A.
We next observed that neither muscular injection of CTX nor caudal fin clip injury of Tg(acvr1l_Q204D) zebrafish resulted in HO formation at the site of injury. Despite observing clear tissue damage at early time points in each model, none of these injuries resulted in HO at later time points. It is possible that the efficient resolution of the injury and lack of HO formation may reflect unique properties of the regenerative abilities of zebrafish, which belong to the small group of vertebrate organisms capable of regenerating many adult tissues. Adult zebrafish have well-studied capacities to regenerate fins, heart, retina, and spinal cord (reviewed in Ref. 40 ). In contrast, adult mammals, including humans, have very limited regenerative capacity. 41 However, adult mammals do possess a robust capacity to regenerate muscle tissue. Therefore, we believe that there may be additional aspects of tissue regeneration in Acvr1lQ204D-expressing zebrafish, which may protect them from HO. For example, it has been determined that the RA signaling pathway is upregulated in early stages of zebrafish fin regeneration, where it plays a key role in establishing the blastema. 42 In models of HO development, RA has been shown to act through RA receptor gamma (RARγ) to inhibit BMP signaling through Smad1/5/8, so that infiltrating fibroproliferative cells do not undergo chondrogenesis and osteogenesis.24,43,44 In fact, the RARγ agonist palovarotene has been demonstrated to prevent HO formation in FOP mouse models,24,45 and has recently shown potential in clinical trials with human FOP patients (www.prnewswire.com/news-releases/phase-2-part-a-open-label-extension-trial-of-palovarotene-for-treatment-of-patients-with-fibrodysplasia-ossificans-progressiva-continues-positive-trends-300429975.html). Future studies may investigate whether Acvr1lQ204D-expressing zebrafish avoid injury-induced HO formation, as seen in these studies, by activating RA signaling in response to injury.
Although CTX and caudal fin clip injury of Acvr1lQ204D-expressing zebrafish did not result in HO at the injury sites, both of these injury models were found to exhibit HO in areas other than the sites of injury. Since these animals were harvested and analyzed 2–4 weeks after injury, it is likely that the HO lesions had already formed in these animals before the time of injury. However, it is possible that injury may have contributed to a heightened level of systemic inflammation in these zebrafish, thereby promoting increased HO lesion size compared to uninjured Acvr1lQ204D-expressing zebrafish. Although not examined in the study described here, this possibility is supported by recent work describing an increased inflammatory response directly preceding mineralization and that existing HO begets more HO. 31 Since systemic inflammation status was not monitored in these experiments, future injury experiments in zebrafish could incorporate the use of MRI to visualize inflammation and edema to more accurately correlate inflammation and HO formation in Tg(acvr1l_Q204D) zebrafish and other zebrafish FOP models as they are created.
The locations of the HO that formed in Acvr1Q204D-expressing zebrafish are both curious and somewhat puzzling. Since the zebrafish body cavity is not expected to experience greater mechanical stimulation from swimming compared with the tail and skeletal tissues, mechanical stimulation is likely not a stimulating factor for these HO lesions. One possible explanation is that the locations where HO forms in Acvr1Q204D-expressing zebrafish are where FAPs normally reside within the zebrafish. FAPs have been found to be a cell source contributing to HO in mouse FOP models.13–15 Future studies will explore the locations, characteristics, and activities of FAPs in zebrafish FOP models. We do not believe that the Safranin O-stained tissues are degenerated oocytes, for the following reasons: the Safranin O staining color in oocytes is a deep purple, while that of the putative HO masses is bright red, and distinct from the cartilaginous tissues in the zebrafish. In addition, we never observed these HO masses in female BRE-GFP heat-shocked or non-heat-shocked control animals. In addition, the IHC staining patterns observed in these putative HO masses were never observed in the ovaries of BRE-GFP control animals.
While the p.Q204D/Q207D variant has been studied in FOP mouse24,46–49 and zebrafish 26 models, it is not a naturally occurring mutation in human FOP patients. In addition, this mutation results in constitutive activation of ACVR1 in a ligand-independent manner and causes significantly more severe phenotypes that the human FOP-associated mutations. The p.Q207E variant is ligand-dependent, while the p.R206H variant can respond to Activin A with increased signaling, but can also signal independent of BMP ligand or Activin A.36,51,52 Therefore, our future work will focus on generating and characterizing zebrafish expressing a p.R203H variant, which is equivalent to the p.R206H variant present in more than 90% of human FOP patients.6,50 We anticipate that Acvr1lR203H-expressing zebrafish may more faithfully exhibit features observed in FOP patients, including the formation of HO in response to injury.
In summary, this study demonstrated that although Acvr1lQ204D-expressing zebrafish do form HO resembling that observed in FOP patients, they do not do so at the site of injury. These results are consistent with recently characterized functional differences between the artificial p.Q204D Acvr1l variant and FOP patient-associated ACVR1 variants, and may also reflect a fundamental difference in the biology of wound healing between zebrafish and mammals. Future studies will address these and other questions, to determine whether novel mechanisms of healing in zebrafish could be exploited to benefit human FOP patients.
Footnotes
Acknowledgments
The authors sincerely thank John Lyons, Patricia Hare, Shaina Colón, Shirin Shabahang, Caroline Banevicius, Annabel Azziz, Michael Chen, and Andrade Hendricks for expert technical support and helpful discussions. This work was supported by NIH/NIDCR R01DE018043 (PCY) and R21AR065761 (PCY), and NSF GRFP NS9344 (ML). The funders had no role in study design, data collection, and analysis, decision to publish, or preparation of the article.
Authors' Contributions
Conceived and designed the experiments: M.L. and P.C.Y. Performed the experiments: M.L. and N.P. Analyzed the data: M.L., N.P., and P.C.Y. Contributed reagents/materials/analysis tools: M.L. and P.C.Y. Wrote the article: M.L. and P.C.Y.
Disclosure Statement
No competing financial interests exist.