
Abstract
Anostomidae species have conserved diploid numbers (2n = 54), although comparative cytogenetic studies have demonstrated chromosomal rearrangements occurrence among them, especially in repetitive DNA rich regions. The location and distribution of ribosomal DNA (rDNA) and small nuclear RNAs (snRNAs) multigene families are highly dynamic in the genomes of several organisms. In this study, we in situ located the rDNA and snRNA sites in two populations of Megaleporinus obtusidens and a sample of Megaleporinus reinhardti to infer their chromosomal changes in the evolutionary lineages. Both species of Megaleporinus shared 2n = 54 chromosomes with the presence of ZZ/ZW sex chromosome system, but they diverged in relationship to the location of 5S and 45S rDNAs as well as the distribution of snRNAs sites. The characterization of the analyzed sequences revealed the presence of complete rDNA and snRNAs sequences as well as snRNAs containing transposable elements (TEs) and microsatellite repeats. After chromosomal mapping, the sequences encompassing TEs proved to be dispersed through autosomes and accumulated on sex chromosomes. The data demonstrate that intra- and interspecific chromosomal changes occurred involving the multigene family's sites in Megaleporinus karyotypes. Furthermore, we detected TE-like sequences in the differentiation of sex chromosome systems in M. obtusidens and M. reinhardti.
Introduction
Anostomidae is one of the most speciose families of Characiformes, comprising 158 valid species 1 from 15 genera, widespread throughout South American hydrographic basins. 2 Despite their remarkable morphological variation, the cytogenetic data in Anostomidae have reported a conserved diploid number (2n) of 54 chromosomes, predominantly metacentric and submetacentric.3–7 Nonetheless, some studies reported a karyotype diversification in this fish group in relationship to the distribution and location of repetitive DNA sequences7–12 and the presence of heteromorphic sex chromosomes.4–6,8,9,11,12
Recently, a new genus of Anostomidae, named Megaleporinus, was validated based on molecular analyses combined with morphological and chromosomal markers. 2 Megaleporinus was recovered as a monophyletic group, 2 in which all species of this genus share sex chromosome systems (ZZ/ZW or Z1Z1Z2Z2/Z1W1Z2W2).5,6,8,9,11–13 The W chromosome in the representatives of Megaleporinus is invariably subtelocentric (st) bearing heterochromatic long arms (q), while the Z chromosome is a middle-sized submetacentric element with small amounts of heterochromatin at distal region of short arms (p).5,6,8 The heterochromatin on the W chromosome of Megaleporinus is derived from the accumulation of repetitive sequences that have probably been responsible for their large size and morphological differentiation.5,6,8,11,12,14,15 For instance, 22 types of satellite DNAs were reported in the repetitive region of the W chromosome in M. macrocephalus, 11 while the sex chromosome of Megaleporinus elongatus encompassed 12 satellite DNAs, 7 of them being shared with M. macrocephalus. 12
Indeed, repetitive sequences represent most of the eukaryote genome and they are organized either in tandem repeats or dispersed through the genome.16–18 Multigene families related to in tandem repetitive DNAs play a key role in the maintenance of the genomes. 19 Despite their evolutionary conservativeness, paralogous copies of a single unit in these multigene families generate intragenomic variation, as well as, lineages diversification, being useful as chromosomal markers.20–24
Ribosomal DNA (rDNA) sequences have been widely used in cytogenetics to infer chromosomal evolutionary pathways in several fish groups.21,25,26 In Anostomidae, most species bear a single 45S rDNA site and one to two clusters of 5S rDNAs on karyotypes.7,27–30 However, in situ mapping of U1 and U2 small nuclear RNA (snRNA) genes revealed that this multigene family is highly variable in both number and location among several fish species.23,31,32 The snRNAs might be clustered into one or several chromosomal pairs23,33,34 or even present on sex chromosomes.32,34
Therefore, the mapping of repetitive DNAs on chromosomes are relevant to the understanding about the interspecific karyotypic diversification of fishes.35,36 In the case of Megaleporinus, repetitive DNAs are also useful to infer the differentiation of sex chromosome systems. Thus, the primary goal of this study was to perform in situ localization and molecular characterization of rDNA and snRNA multigene families isolated from the Megaleporinus obtusidens genome to evaluate their possible involvement in the chromosomal changes in this species group. Furthermore, the data permitted to infer the role of the transposable element (TE)-like sequences in W-specific chromosome region differentiation in M. obtusidens and Megaleporinus reinhardti.
Materials and Methods
Sampling
The specimens of M. obtusidens were collected in Iguaçu River (Iguaçu River basin) in União da Vitória, Paraná (−26°13′5.22″/−51°16′17.40″) and Piumhi River (São Francisco River) in Piumhi, Minas Gerais (−20°16′15″/−45°55′39″). Individuals of M. reinhardti were collected in Gongogi River (Contas River basin) in Dario Meira, Bahia (−14°28′13.1″/−39°57′33.1″). All procedures were carried out with authorization of the Ethics Committee of Animal Usage of Universidade de Ponta Grossa (CEUA/UEPG 011/2017) and Universidade Estadual do Sudoeste da Bahia (CEUA/UESB 32/2013). M. obtusidens was identified and deposited in the ichthyological collection of the “Núcleo de Pesquisas de Limnologia, Ictiologia e Aquicultura” (NUPÉLIA) from Universidade Estadual de Maringá, while M. reinhardti was identified by experts from the Universidade Estadual Paulista, Campus Júlio Mesquita Filho.
Conventional cytogenetic analysis
Mitotic chromosomes were obtained from kidney cells according to Bertollo et al. 37 with modifications. 38 The chromosomal spreads were stained with 5% Giemsa solution in phosphate buffer pH = 6.8. Heterochromatic regions were detected as described by Sumner. 39
Obtaining and characterization of rDNA and snRNA sequences
Genomic DNA was isolated from the liver tissue of M. obtusidens using “Illustra Cell Genomic Prep Mini Spin Kit” (GE Healthcare, Chicago, IL) following the manufacturer's instructions. The 5S and 18S rDNAs and the U1 and U2 snRNAs sequences were amplified via polymerase chain reaction (PCR) as follows: 100 ng of template DNA, 1 × reaction buffer (200 mM Tris, pH 8.4, 500 mM KCl); 1.5 mM MgCl2, 0.2 mM dNTPs, 0.2 mM of each primer, and 1 U Taq DNA Polymerase (5 U/μL). The amplification reaction encompassed a first denaturation step at 95°C for 5 min, 35 cycles at 95°C for 30 s, 56°C for 45 s, 72°C for 2 min, plus a final extension at 72°C for 7 min. The primers used in PCR reactions are described in Supplementary Table S1.
PCR products (5S rDNA, 18S rDNA, U1 snRNA, and U2 snRNA) were purified using the “GenElute™ PCR Clean-Up Kit” (Sigma-Aldrich), then cloned with the “pGEM®-T Easy Vector Systems Kit” (Promega, Madison, WI) in Escherichia coli DH5α and sequenced in ABI-PRISM Genetic Analyzer (Carlsbad, CA). The sequences were analyzed using the software GENEIOUS 7.1.9 (Ref. 40 ) and characterized using the Nucleotide Basic Local Alignment Search Tool (BLASTn), 41 CENSOR software, 42 and Rfam 43 tools. To check the occurrence of protein domains, the sequences were submitted to Pfam database. 44 All obtained sequences were submitted to Dotplot in the software Geneious for comparative analysis using the Dottup tools for the search of in tandem repeats.
Probe labeling
18S rDNA and U1 snRNA (clone possessing TEs) PCR-based probes were labeled with digoxigenin 11-dUTP using the “Digoxigenin NT Labeling Kit” (Jena Bioscience, Dortmund, Germany), according to the manufacturer's instructions. U2 snRNA (clone possessing TEs) probe was labeled with biotin 16-dUTP using the “Biotin16 NT Labeling Kit” (Jena Bioscience). 5S rDNA probe was labeled via PCR using biotin 16-dUTP (Jena Bioscience). After confirming the identity of the amplified sequences, specific probes for both U1 and U2 snRNAs (without TEs segments) were labeled with digoxigenin 11-dUTP (Jena Bioscience) via PCR.
Fluorescence in situ hybridization
Fluorescence in situ hybridization (FISH) was performed as described by Pinkel et al. 45 using the following stringency conditions: 2.5 ng/μL of each probe, 50% formamide, 2 × saline-sodium citrate (SSC), 10% dextran sulfate, and hybridization at 42°C for 16 h. Fluorescent signals were detected using Streptavidin Alexa Fluor 488 (Molecular Probes, Carlsbad, CA) and anti-digoxigenin rhodamine Fab fragments (Roche Applied Science, Penzberg, Germany). Chromosomes were counterstained with 0.2 μg/mL of 4′6-diamidino-2-phenylindole in VETASHIELD mounting medium (Vector, Burlingame, CA).
Karyotypic analyses
Chromosomal preparations were analyzed in light and epifluorescence microscope (Zeiss Axio Lab 1) equipped with the software Zen Lite and a Zeiss AxioCam ICM1 camera with resolution of 1.4 megapixels (Carl Zeiss, Oberkochen, Germany). Chromosomes were classified based on the arm ratio (r) according to Levan et al. 46 as follows: metacentric (m), r from 1 to 1.7; submetacentric (sm), r from 1.7 to 3; subtelocentric (st) r from 3 to 7; acrocentric (a) r more than 7; and were arranged in decreasing size in the karyotypes.
Results
Sequence analyses
5S rDNA sequence of M. obtusidens comprised 205 base pairs (bp), 97.7% of similarity with the 5S rDNA of Leporinus obtusidens (AF284732.1) and E-value 7.7 e-25 in Rfam (Supplementary Table S2). This sequence encompasses a transcribed region of 123 bp and a nontranscribed spacer (NTS) of 82 bp (Supplementary Fig. S1a). Conserved regions related to the internal promoter Box A, intermediary element, Box C, poli-T, TATA-Like region, Box GC, and Cytosine-1 were also observed in the 5S rDNA of M. obtusidens (Supplementary Fig. S1a). The partial sequence of the 18S rDNA of M. obtusidens comprised 1078 bp, 98% of similarity with the 18S rDNA of Clarias sp. (KF650737.1) and E-value of 2.2 e-295 in Rfam. When submitted to CENSOR, this sequence was characterized by the presence of an internal segment of 85 bp (segment 471–555 bp) with 65.88% of similarity in relationship to the non-long terminal repeat (non-LTR) R2-N1_Gav retrotransposon (RTE), but TE protein domain were not detected (Supplementary Table S2; Supplementary Fig. S1b).
Four amplicons were recovered and sequenced from the PCR products using the U1 snRNA primers: 160, 563, 796, and 916 bp. The 160-bp sequences presented a similarity of 82.91% in relationship to the U1 snRNA of Xiphophorus hellerii (LOC116716575) with E-value of 1.1 e-21 in Rfam (Supplementary Table S2; Supplementary Fig. S1c). When analyzed using CENSOR, this fragment was 78.85% similar to the satellite DNA SAT-17_DR (Supplementary Table S2; Supplementary Fig. S1c). The sequences with 563, 796, and 916 bp had no similarities with the U1 snRNA, but they encompassed segments similar to TEs. The sequence with 563 bp presented 82.50% and 86.36% of similarity with the DNA transposon hAT-N45C_DR, between the 399–438 and 501–543 segments, respectively (Supplementary Table S2; Supplementary Fig. S1d). The 796-bp sequence was similar to the following TEs: RTE non-LTR L2–2 EL (71.57% of similarity between 25 and 128 bp), RTE non-LTR L2–41_Dre (87.50% of similarity from 239 to 278 bp position), RTE LTR Copia-9_GAr-LTR (78.33% in the segment 175–234 bp), and DNA transposon Mariner TIGGER6A (77.05% in the segment 486–602) (Supplementary Table S2; Supplementary Fig. S1e). On the contrary, the 916-pb sequence was 70.99% similar to the RTE short interspersed nuclear element (SINE) SINE2-1C_CCa (147–274 bp), 72.81% similar to the RTE Penelope-1_CGi (577–698 bp), and 71.23% similar to the DNA transposon hAT-N64B_DR (segment 403–558 bp) (Supplementary Table S2; Supplementary Fig. S1f).
In the case of the U2 snRNA, two amplicons were obtained and sequenced, with 177 and 1464 bp, respectively. The 177-bp sequence presented a similarity of 94.25% with the U2 snRNA of Characidium gomesi (KY566214.1) and E-value of 9.9 e-36 in Rfam (Supplementary Table S2; Supplementary Fig. S1g). Based on CENSOR analysis, this sequence was 90% similar to the RTE non-LTR Keno-1_SSa in the segment from 37 to 177 bp (Supplementary Table S2; Supplementary Fig. S1h). The 1464-bp sequence encompassed a complete copy of the U2 snRNA (segment 599–789 bp) with 94.25% of similarity with the U2 snRNA of C. gomesi (KY566214.1) and E-value of 1 e-39 in Rfam (Supplementary Table S2; Supplementary Fig. S1h). The analysis of this amplicon in CENSOR revealed segments with 89.86% of similarity with the RTE non-LTR Keno-1_SSa (segment 100–248 bp) and the DNA transposon hAT-2_DK (segment 810–839 bp) (Supplementary Table S2; Supplementary Fig. S1h). The graphic representation using Dotplot also revealed in tandem microsatellite repeats within sequence 1464-bp long, being the (CTTT)n microsatellite located in the segment from 896 to 942 bp (Supplementary Fig. S1i). Sequences bearing specific TE protein domain segments were not identified in Pfam analysis (Supplementary Table S2; Supplementary Fig. S1h). All obtained sequences were stored in GenBank (MH697559.1; MH701851.1; MT563073–MT563075).
Karyotyping
The specimens of M. obtusidens (Iguaçu and Piumhi Rivers) and M. reinhardti (Gongogi River) shared a karyotype with 2n = 54 chromosomes distributed into 28m+26sm in males and 28m+25sm +1st in females with a fundamental arm number of 108 (Fig. 1). The difference in the karyotype formulae between males and females refer to the presence of a ZZ/ZW sex chromosome system, in which the Z corresponds to a middle-sized sm chromosome and the W is a large st chromosome (Fig. 1a, c, e).

Karyotypes of Megaleporinus obtusidens from Iguaçu River
C-banding revealed small amounts of heterochromatic at pericentromeric region of some autosomes in both populations of M. obtusidens (Fig. 1b, d) and in M. reinhardti (Fig. 1f). In addition, terminal C-bands on p arm of one homologous of the pair 17 was observed in M. reinhardti (Fig. 1f). The W chromosome presented a large heterochromatic block comprising nearly the entire q arm in all populations and species herein analyzed (Fig. 1b, d, f). On the contrary, the Z chromosome was characterized by a small heterochromatin segment from the middle to distal position on q arms in the studied populations and species (Fig. 1b, d, f).
In situ localization
In M. obtusidens from Iguaçu River, the double-FISH using rDNA probes revealed a single p terminal 5S rDNA cluster in the m pair 5 and a single q terminal 18S rDNA site in the sm pair 15 (Fig. 2a). In this population, the species-specific U1 snRNA probe was observed at terminal q region in the m pair 1, as well as at proximal p region in the pair 15 (Fig. 2b). The specific U2 snRNA probe revealed clusters at the p proximal region of the pair sm 17 in M. obtusidens from Iguaçu River (Fig. 2c).

In situ location by FISH of the 5S rDNA
In M. obtusidens from Piumhi River, the 5S rDNA was located at p terminal region of the m pair 8, while the 18S was identified at the p terminal region of the m pair 5 (Fig. 2d). The U1 snRNA sequence was mapped onto the q arm terminal region of the m pair 1 (Fig. 2e) while the U2 snRNA was visualized at the p proximal region of the sm pair 17 (Fig. 2f).
In M. reinhardti, the 5S rDNA was located at the p terminal region of the m pair 8 and the 18S rDNA was located at the p terminal region of the m pair 2 (Fig. 2g). The in situ localization of U1 snRNA on chromosomes revealed signals at the q terminal region of the m pair 1 and at the terminal q region of the sm chromosome 15 (Fig. 2h), whereas the U2 snRNA was in situ located on the p proximal region of the sm pair 20 in this species (Fig. 2i).
FISH procedures using the total PCR products using the snRNA primers, which comprised TEs, evidenced several dispersed and accumulated signals in both species and populations (Fig. 3). In situ location of the four amplicons containing TEs after amplification of the U1 snRNA indicated accumulation of these sites at terminal regions of most autosomes, at pericentromeric regions of a few pairs, and some dispersed signals on the sex chromosomes of the analyzed populations and species (Fig. 3a, c, e). In addition to the specific U2 snRNA signals, the FISH using the U2 snRNA probe encompassing the 1464-bp amplicon also revealed dispersed sites corresponding to Keno and hAT TEs in other autosomes and in both Z and W chromosomes of the analyzed samples of Megaleporinus (Fig. 3b, d, f).
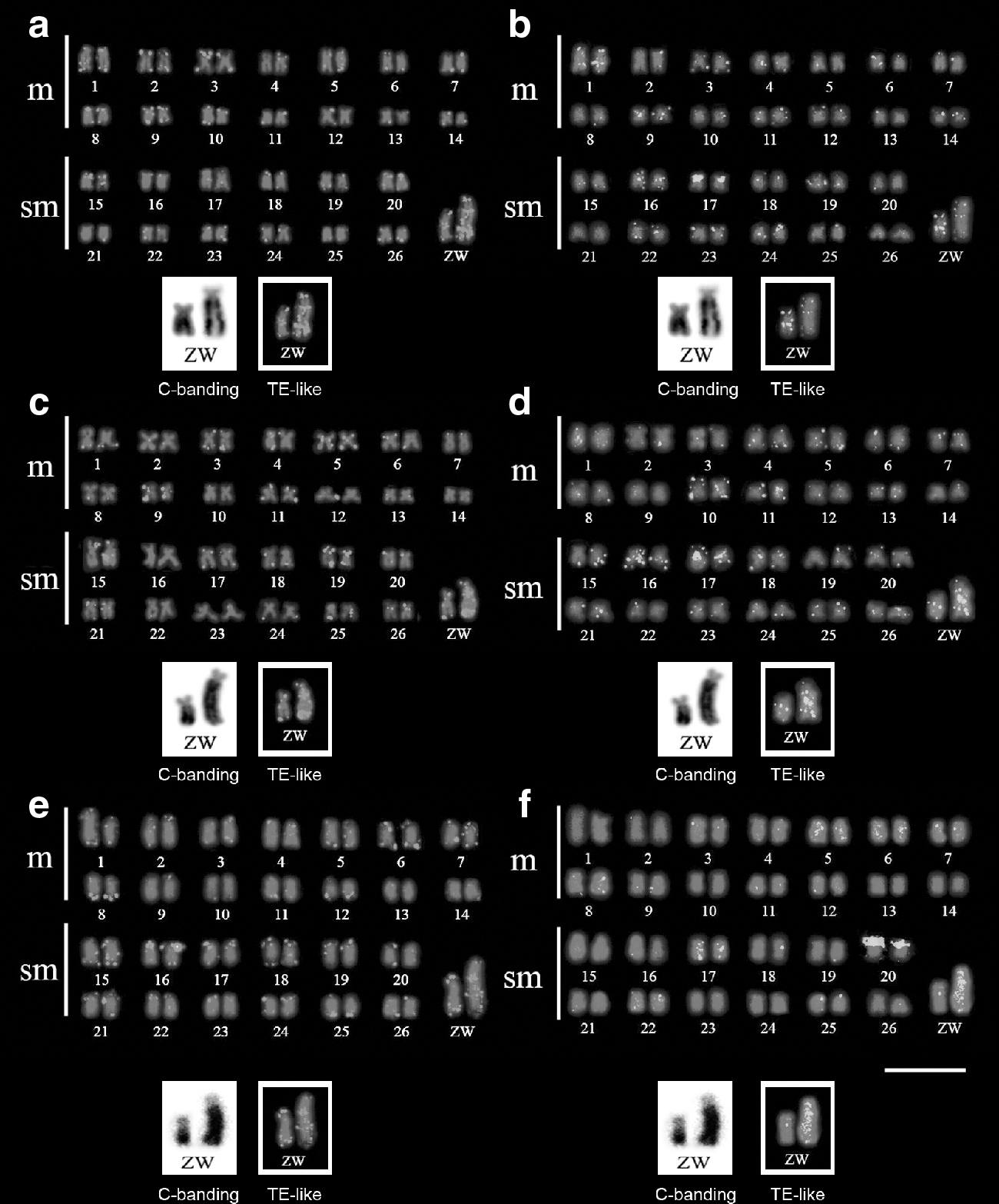
Karyotypes of Megaleporinus obtusidens from Iguaçu River
Discussion
All analyzed specimens of M. obtusidens and M. reinhardti shared the conserved 2n and karyotype macrostructure proposed for Anostomidae. 30 Moreover, both populations M. obtusidens and the samples of M. reinhardti presented a ZZ/ZW sex chromosome system, a cytogenetic feature that corroborates the monophyletism of Megaleporinus, 2 indicating that the occurrence of sex chromosomes is a synapomorphic condition for the Megaleporinus genus.
The heterochromatin is mostly distributed into pericentromeric or terminal segments of a few chromosomes in Anostomidae.4,6,7 On the contrary, the sex chromosomes of Megaleporinus are characterized by conspicuous heterochromatin blocks, particularly on q arms of the W chromosome,5,9,10 reinforcing the role of repetitive DNA in the origin of the heteromorphic sex chromosomes of Megaleporinus.11,12
The presence of two clusters of 5S rDNA, including a larger and terminal cistron and a minor and interstitial one is a typical trait of some Anostomidae.27–29 However, M. obtusidens and M. reinhardti revealed a single 5S rDNA site, as also reported in other species of this family.7,30 The analysis of the molecular structure of the 5S rRNA gene in M. obtusidens herein carried out is a predictor of its functional organization. In turn, the occurrence of single 45S rDNA site, as presently observed, is commonly reported in most species of Anostomidae.7,27–30 Moreover, a few representatives of this fish family present syntenic 5S and 18S rDNA clusters.7,30 Therefore, these data reveal the dynamics of the chromosomal location of 5S rDNA and 18S rDNA clusters in Anostomidae.
The distinct location of rDNA families on chromosomes might affect the recombination rates between genomes of recently diverged species. 26 Indeed, some groups of Neotropical fishes are characterized by rearrangements involving the distribution of rDNA families among populations or incipient species.36,47,48 Accordingly, the present results in M. obtusidens and M. reinhardti as well as comparative cytogenetic reports based on chromosomal mapping of ribosomal cistrons revealed the high mobility of rDNA on chromosomes of several representatives of Anostomidae.7,27–30
Sequencing of snRNA amplicons in M. obtusidens revealed some degree of unspecific amplification of TEs, and inasmuch as the multigene families might be associated with TEs, their sequences can be duplicated as a consequence of transposed mediated by TEs. 49 Under these circumstances, one portion of multigene family duplicated copies would be composed of interrupted and probably inactive genes. 49 Conversely, some small segments of TE similar within multigene families without an invasion assinature or bearing a specific TE protein domain, as observed in 18S rDNA sequence, could represent a sequence artifact or evolutionary convergence. Other studies reported that some TEs presented snRNA-like internal segments.23,50,51 Nonetheless, the identity values obtained for the partial sequence of the U1 snRNA isolated from M. obtusidens indicate that it corresponds to one copy of U1 snRNA gene family. In addition, the genetic relationship of the U1 snRNA sequence herein studied with SAT-17_DR should reflect only the similarity between the sequences due to origin of this satellite sequence from U2 snRNA defective copies in D. rerio genome. The genetic characterization of three other amplicons (563, 796, and 916 bp) obtained using the U1 snRNA primers revealed that they lack functional units of the multigene family. Instead, these amplicons are enriched with fragments of hAT, L2, Copia, Mariner, SINE2, and Penelope TEs. This result corroborates the inferences from Schemberger et al. 52 demonstrating that different TE-like sequences might be interspersed in the genome of M. obtusidens, especially on sex chromosomes.
As for the U2 snRNA sequences, the 177-bp unit was compatible with the transcribed region of functional copies of this gene. In turn, the 1464-bp sequences comprised a transcribed region of U2 snRNA, fragments of hAT DNA transposon, and microsatellite repeats within the NTS region. The invasion of NTS in multigene families by TEs has been reported in 5S rDNA and histones families.23–25,53 Nonetheless, the identification of Keno TE units in the U2 snRNA sequences might correspond to the similarities between both sequences23,50,51 rather than to the presence of this class of TE in the genome of M. obtusidens. Thus, the present data revealed that the specific and functional sequences of U1 and U2 snRNAs were distributed on a single to two chromosomal sites in M. obtusidens and M. reinhardti karyotypes, which have chromosomal location specific for each evolutionary lineage due to chromosomal rearrangements occurrence. Likewise, the presence of one or a few chromosomal pairs bearing U2 snRNA signals has been observed in several organisms.23,32,54–63
Accumulated copies of U1 snRNA and dispersed signals of U2 snRNA were reported on the W chromosome of Triportheus. 62 In Gymnotus pantanal, the snRNAs were detected in X1 but absent in the Y chromosome. 32 In the present study, the sex chromosomes of both M. obtusidens and M. reinhardti accumulated TE-like sequences instead of snRNA cistrons. Previous reports inferred that TEs should have played a key role in the origin of heteromorphic sex chromosomes in several species.63–67 In situ localization in M. obtusidens and M. reinhardti karyotypes herein reported showed that the invasion of TEs favored the differentiation of the heteromorphic region in the W chromosome. In addition to TEs sequences, sex chromosomes of Megaleporinus species are also enriched with satellite repeats.11,12 At the moment, the differences in TE-like sequences accumulation between Z and W sex chromosomes in Megaleporinus species suggest their participation on sex chromosomes nonrecombining region origin events, and in W-specific chromosome heteromorphism formation.
Therefore, the chromosomal changes in Anostomidae species seem to be highly influenced by several transpositions of multigenic sites, leading to karyotype diversification among evolutionary lineages. This study also demonstrated that U1 and U2 snRNA sequences might encompass parts of TEs and microsatellite units and, therefore, only the transcribed region of these genes should be used for in situ chromosomal mapping. Moreover, sequences containing parts of TEs were dispersed through the karyotypes and accumulated in sex chromosomes, probably leading to the differentiation of sex chromosome system in M. obtusidens and M. reinhardti.
Footnotes
Acknowledgments
The authors are grateful to ICMBio (Instituto Chico Mendes de Conservação da Biodiversidade) for authorizing the collection of samples (license numbers 10538-1, 15117-1, 26752-3).
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Fundação Araucária (Fundação Araucária de Apoio ao Desenvolvimento Científico e Tecnológico do Estado do Paraná), FAPESP (Fundação de Amparo à Pesquisa do Estado de São Paulo—2015/16661-1), FAPESB (Fundação de Amparo à Pesquisa do Estado da Bahia), CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Finance Code 001), and CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico—305142/2019-4).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.