
Abstract
Hyperglycemia, a primary symptom in diabetes mellitus, is associated with difficulties in wound healing and regeneration. This condition is due to the length of the inflammatory phase and free radicals. Furthermore, there is evidence that molecular pathogenesis is involved in impaired wound healing in diabetics. As an animal model, zebrafish have many shared orthologous genes with human that are involved in protein regulation of wound healing and regeneration. Little is known about natural drugs that may be used to treat complications of wound healing in diabetes. Propolis, however, is known to consist of various organic compounds such as phenols and flavonoids with antioxidant and anti-inflammatory activities. This research aims to study propolis' effect on caudal fin regeneration and relative expression of several genes belonging to Hedgehog, bone morphogenetic protein (BMP), and Wnt signaling hyperglycemic (HG) zebrafish. GC-MS analysis and antioxidant activity testing were performed on ethanolic extract of propolis (EEP). Caudal fin regeneration was analyzed using ImageJ; blood glucose levels were measured; and relative gene expression analysis of shha, igf2a, bmp2b, and col1a2 was performed by the real-time polymerase chain reaction method with the β-actin housekeeping gene. Impairment of caudal fin regeneration in zebrafish hyperglycemia was characterized by a low percentage of regeneration and decreased relative gene expression. EEP at 15 ppm could increase the percentage of caudal fin regeneration and the expression of shha, igf2a, bmp2b, and col1a2. Based on the results, it appears that phenols and flavonoids from the EEP can improve the caudal fin regeneration of HG zebrafish.
Introduction
Diabetes mellitus (DM) belongs to the noncommunicable metabolic disease group characterized by elevated glucose levels in the blood (hyperglycemia) caused by reduced insulin hormone secretion, insulin resistance, or both. 1 This disease is also accompanied by adverse complications that include retinopathy, nephropathy, neuropathy, macrovascular, heart attack and stroke, and diabetic foot ulcer. 2 One of the long-term complications that produces the significantly degraded quality of life caused by hyperglycemia is impaired wound healing.3,4 Impaired wound healing can lead to secondary complications, such as an inflammatory phase around the wound, increased risk of infection, decreased cell number and growth factor response resulting in decreased local angiogenesis, and inhibition of blood flow in peripheral blood vessels to chronic wounds and amputations. 5 Studies showed that DM might cause the disruption of marker genes for regeneration, especially genes that are involved in the wound healing process6–8 due to endothelial cell damage, decreased or disrupted growth factor production as a result of angiogenic response (formation of new blood vessels), macrophage function, collagen accumulation, epidermal barrier function, number of granulation tissues, migration and proliferation of fibroblasts, the number of epidermal nerves, as well as the balance between the accumulation of extracellular matrix components and their remodeling by the metalloproteinase matrix. 5
Some of the essential molecular mechanisms causing endothelial cell damage are activated by (1) increased intracellular activity of the intracellular sorbitol (polyol) pathway,9–13 (2) improved advanced glycation end products11,12,14 and (3) improved receptor expression,15,16 (4) activation of protein isoform kinase C, and (5) overactivity of the hexosamine line.17,18 These processes are induced by hyperglycemia, which produces free radicals that cause oxidative stress. 18
Based on a previous study, molecular pathogenesis involving impaired wound healing can be observed in an animal model system such as zebrafish.4,19 Studies showed that diabetic zebrafish exhibit complications such as diabetic retinopathy and nephropathy and show disruption in the wound healing process and caudal fin regeneration.4,20 Moreover, zebrafish have many orthologous signaling pathways that play essential roles in regulatory pathways and are associated with wound healing, including Hedgehog (HH), BMP, and Wnt signaling pathway.21–26 The pathways listed above are involved in regeneration stages, including wound healing (inflammation, reepithelialization, and remodeling), blastema formation, regeneration growth, and proliferation.27–31
The administration of synthetic chemical drugs and physical therapies to overcome the wound healing and tissue regeneration deficiencies during hyperglycemia is considered less effective if administrated in the long term.32–34 Natural drugs based on natural compounds to minimize the side effects of synthetic drugs have been developed as an alternative,. Some natural remedies are known to have antioxidant activity and low side effects.32,35,36 These antioxidant compounds have neutralized free radicals effectively37,38 and may decrease complications in the pathogenesis of DM. 39 One natural product known for its antioxidant activities is propolis. 40 The role of antioxidants from propolis and its mechanism on wound healing and tissue regeneration in hyperglycemia conditions have not been widely studied. Therefore, research on the potency of ethanolic extract of propolis (EEP) in inducing wound healing and regeneration that analyzes the expression of several marker genes involved in both processes during hyperglycemia still needs to be done. This study may provide new knowledge as a first step in discovering natural drug candidates from propolis with safer application to wound healing in the conditions of hyperglycemia.
Materials and Methods
The experimental protocols were approved by the appropriate Institutional Review Committee and meet the guidelines of the responsible governmental agency. Ethical exemption no. 281/UN6.KEP/EC/2021.
Zebrafish maintenance
Three- to 6-month-old wild-type adult zebrafish (Danio rerio) were used in this research. All the fish were obtained from a local breeder from Bogor, West Java, Indonesia. They were maintained in a terrarium at the School of Life Sciences and Technology, Institut Teknologi Bandung (ITB), under standard laboratory conditions (14 h light period/10 h dark period at 28.5°C). Zebrafish were fed with Tetramin™ twice a day and supplemented with artemia.
Propolis extraction
Crude propolis of Trigona laeviceps was obtained from a local stingless bee farm in Subang, West Java, Indonesia. Seventy percent ethanol was added to clean propolis in a 1:20 ratio. Propolis was blended and shaken at 200 rpm in a dark condition for 1 week. EEP was harvested using Whatman No.1 filter paper.41,42 The maceration process was repeated 1 × 24 h until the EEP became colorless. The collection of EEP was evaporated with rotavapor and dried with a steam cup at temperature 40°C–50°C to form EEP powder. The extraction was conducted at the Laboratory of Pharmaceutical Natural Product, Research Center for Biosciences and Biotechnology, ITB.
Preliminary study
GC-MS and 2,2-diphenyl-1-picrylhydrazyl analysis
GC-MS analysis was performed to examine the chemical compounds in the EEP.41,43 GC-MS Analysis was done in Laboratory Testing Forest Products, Forest Products Research, and Development, Ministry of Forestry, Bogor, West Java, Indonesia. Before application on the zebrafish, the antioxidant activity of EEP was determined by 2,2-diphenyl-1-picrylhydrazyl (DPPH) analysis44,45 at Biopharmaca Research Center, Bogor Agricultural University, Bogor, West Java, Indonesia.
Induction of hyperglycemia
We used a combination of alloxan (Sigma-Aldrich®) 0.4% (400 mg alloxan in 100 mL NaCl) and glucose (E-Merck®) % solution in various concentrations to induce hyperglycemia in the zebrafish. 44 Twenty fish (divided into two containers containing 10 fish) swam in 100 mL alloxan solution with the duration of 1 h per day for 5 days, then were transferred into 2 L of 2% glucose solution for 6 days with the duration of 24 h per day with medium replacement every 24 h. 46
Treatment with EEP
DPPH-analysis showed antioxidant activity of propolis with IC50 at a concentration of 15 ppm. EEP powder was dissolved in propylene glycol (a nontoxic solvent, 1% of the tank's total water volume) before being mixed into a 2-L water tank at the container. 46
Treatment with metformin
Metformin is an antidiabetic drug used in this study as a positive control. Metformin concentration was chosen at 10 μM, according to Capiotti et al. 47 who reported that this concentration lowered blood glucose levels of hyperglycemic (HG) zebrafish after 2% glucose solution exposure.
Blood glucose measurement
Measurement of blood glucose levels was performed at 96 hpa (hours postamputation) using the OneTouch® UltraEasy® blood glucose meter. Five fish from each treatment group fasted for 24 h before blood collection. The fish were euthanized using the rapid freezing method.48,49 The blood was obtained by cutting the head of the fish near the heart area toward the gills.4,20,50
Caudal fin regeneration analysis
Zebrafish were anesthetized using 100 mL Tricaine (MS-222) 0.6 mM for ±1–2 min. Five fishes for each treatment group were used for caudal fin regeneration analysis. The fish were placed in a Petri dish with a transverse orientation so that the caudal fin was located on the right-hand side. The observation of the fin was carried under a microscope. Caudal fin was stretched carefully to the visible bony ray (segmented bone in zebrafish caudal fin) and amputated using a lancet (Aesculap Scalpel Handle No. 3 and Aesculap Scalpel Blade No. 10 of B BRAUN) at the first or second segment below the level of the first ray bifurcation. Either bony ray no. 3 or no. 4 was selected as as a reference.51,52 Postamputation fish were returned to the tank according to treatment group in its related solutions alongside alloxan/glucose and allowed to regenerate. Caudal fin regeneration was observed at 0, 24, 48, 72, and 96 hpa with a stereomicroscope (Nikon HFX DX) and camera (Canon Power Shot A650IS PC 1252 DC 4.3V; Carl Zeiss 426126). The area of regeneration was traced using ImageJ 1.48v software to determine the percentage of regeneration.
Relative gene expression analysis
Gene expression analysis of shha, igf2a, bmp2b, and col1a2 with housekeeping gene β-actin was performed using reverse transcription-quantitative polymerase chain reaction (RT-qPCR). The shha, igf2a, bmp2b, col1a2, and β-actin (actb1) primers were ordered and purchased from IDT (Integrated DNA Technologies®) with specific primersequences shown in Table 1.
Sequences of Oligonucleotide Primers
The analysis began with RNA isolation of the regenerating caudal fin blastema at 96 hpa from 20 fish using peqGOLD TriFast™ reagent (PEQLAB Biotechnologie GmbH). After digestion with DNAse I (Thermo Fisher Scientific) to eliminate genomic contamination, cDNAs were synthesized using RevertAid™ First Strand cDNA Synthesis kit (Thermo Fisher Scientific). Real-time qPCR was conducted using SsoFast™ EvaGreen® supermix (Bio-Rad) with CFX96 Touch Real-Time PCR Detection System (Bio-Rad). The housekeeping gene, actb1, was used as a reference for internal standardization. The relative cDNA expression was calculated using the method of 2−ΔΔCt. 53
Statistical analysis
Data analysis was performed in three categories, that is, blood glucose level analysis, caudal fin regeneration, and relative expression of the marker genes. Blood glucose level analysis was performed using a one-way normality test and a one-way analysis of variance (ANOVA) with a Games-Howell post hoc test. In contrast, analysis of the relative expression of the marker genes was performed using a one-way ANOVA test of normality and a post hoc Tukey Honest Significant Difference test to determine significant differences between the treatment groups. Data are presented as mean ± standard deviation.
Result and Discussions
GC-MS and DPPH analysis of EEP
Based on the result of GC-MS analysis, EEP consisted of 57% polyphenols compounds, such as guaiacol, cresol, dimethoxy phenol, ethyl phenol, ethyl guaiacol, methoxy phenol, methyl phenol, pyrocatechol, and phenolic acid (benzoic acid/benzaldehyde) (Fig. 1) that are known to have antioxidant and anti-inflammatory activity.52,54 The antioxidant activity55–60 of phenolic compounds of EEP is due to the aromatic ring structure containing at least one hydroxyl group. 61 Therefore, these compounds can inhibit reactive oxygen species (ROS) derived from alloxan by metabolizing excess glucose by donating an electron or hydrogen atom to the free radicals.52,54,62 The result of our DPPH analysis supported these prior findings by showing that EEP was able to reduce 50% of free radicals DPPH at a concentration of 14.53 ppm (IC50) (compared with Vitamin C that had IC50 at a concentration of 2.54 ppm), with the best result showed on the application of 15 ppm (data not presented).
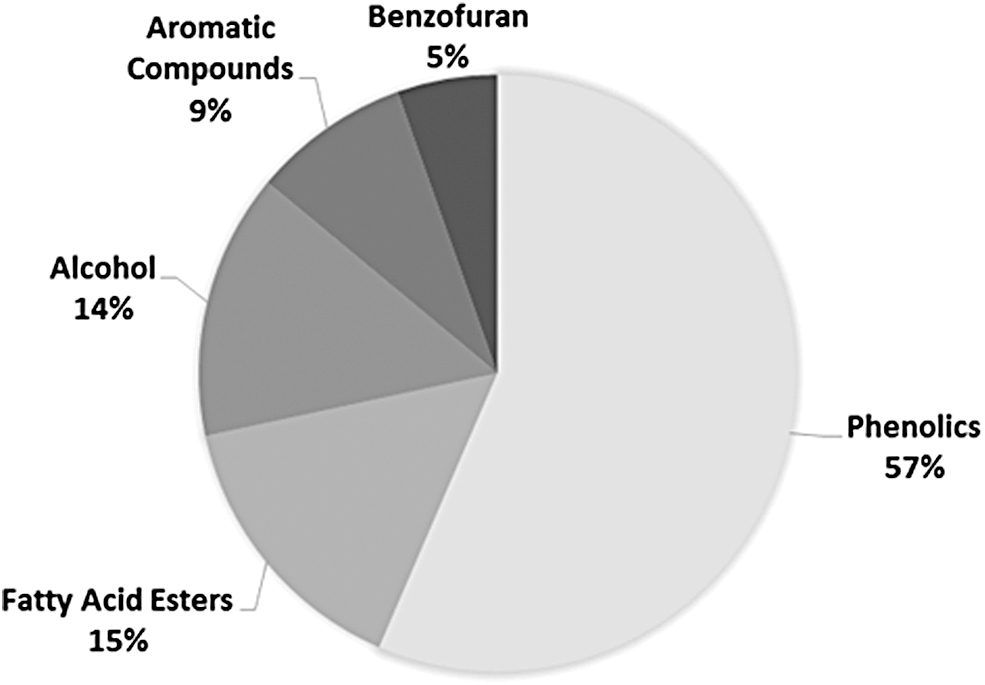
The GC-MS analysis results show the chemical components of the EEP. Polyphenols are known to be the majority compound. EEP, ethanolic extract of propolis.
Effect of EEP on blood glucose levels
HG and Hyperglycemic-Propylene Glycol (HGPG) groups are known to have the highest blood glucose levels of 311.8 ± 36.0 mg/dL and 286.8 ± 41.4 mg/dL, respectively (Fig. 2), and these levels can be categorized as hyperglycemia, as noted by Gleeson et al. and Shin et al. who showed that fish with an average blood glucose level >200 mg/dL are hyperglycemia.44,63 The later study also showed that the application of alloxan and glucose in swimming water can produce HG zebrafish. 44 Alloxan and glucose are thought to enter the body of fish through aquaporins (AQP) and GLUT1, present in the gill and skin epithelium. 64 GLUT1 is one of the insulin-independent isoforms of glucose transporters 65 and is spread over various tissues, including epithelial skin, epithelial gills, brain, astrocytes, and oligodendrocytes. 64
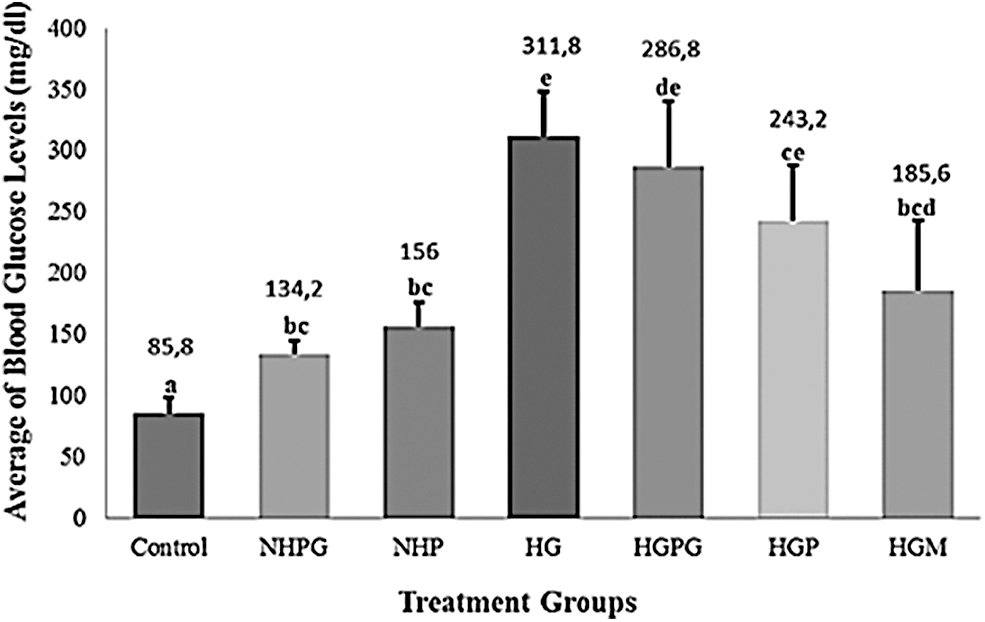
The average of fasting blood glucose levels (mg/dL) at 96 hpa (hours postamputation). Control: normal zebrafish. NHPG: normal zebrafish+propylene glycol 1%. NHP: normal zebrafish+propolis 15 ppm. HG: zebrafish hyperglycemia (alloxan 400 mg/100 mL NaCl+Glucose 2%). HGPG: zebrafish hyperglycemia+propylene glycol 1%. HGP: zebrafish hyperglycemia+propolis 15 ppm. HGM: zebrafish hyperglycemia+metformin 10 μM. At least five zebrafish were used for each group. Data are shown as mean ± standard deviation. Statistical analysis of the data used a one-way ANOVA followed by a Games-Howell post hoc test. The different lowercase letters (a, bc, e, de, ce, and bcd) show statistical significance compared to the control group (p < 0.05). ANOVA, analysis of variance; HG, hyperglycemic.
Matsushige et al. suggested that propolis can protect pancreatic beta cells in mice against the toxic effect of the diabetogenic agent streptozotocin, because propolis-treated mice exhibited lower blood glucose levels compared to mice not administered with it. 66 Furthermore, alloxan-treated mice and obese rats administered propolis for a long period (8 and 12 weeks) showed significantly lower blood glucose levels.67,68 Several studies showed supporting data on the antioxidant activity of EEP in decreasing blood glucose levels after a long exposure time in zebrafish.40,41
Effect of EEP on caudal fin regeneration and gene expression (shha, igf2a, bmp2b, and col1a2)
The growth of blastema as one of the initial stages of regeneration is a characteristic of zebrafish caudal fin regeneration.22,69 Our results showed improved caudal fin regeneration in both the propolis-treated and positive control groups compared to HG fish (Figs. 3 and 4). Hyperglycemia conditions in zebrafish have been shown to prolong the inflammatory phase.70,71 and thought thereby to increase oxidative stress and secretion of interleukin-8 on keratinocyte. 72 However, the expression of immune-related cytokines could also be associated with ischemia of amputated caudal fin and caused by an excessive hypoxic condition. 17
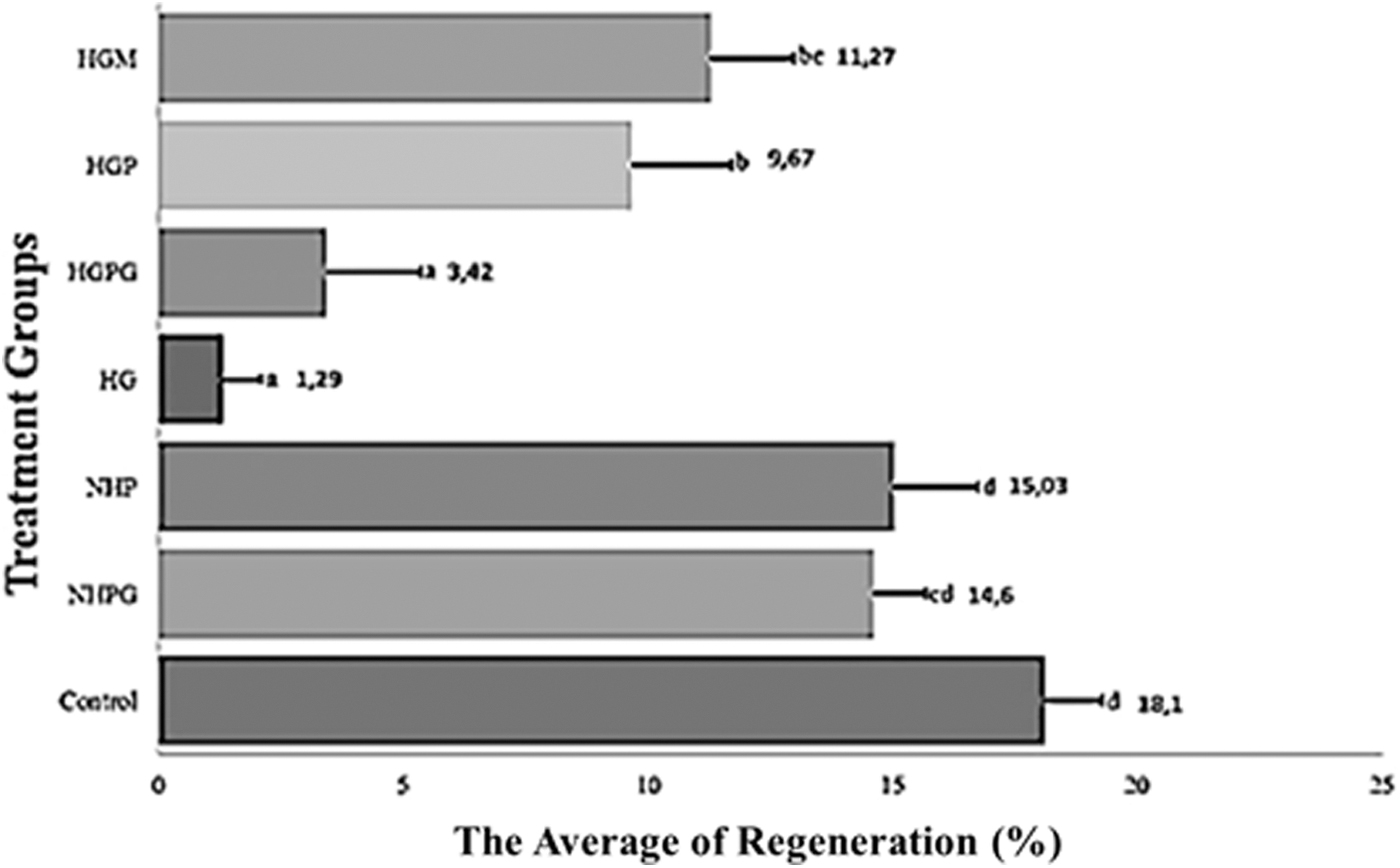
The average percentage of zebrafish caudal fin regeneration at 96 h postamputation. At least five zebrafish were used for each group. Data are presented as mean ± standard deviation. The different lowercase letters (d, cd, a, b, and bc) show the statistical significance compared to the control group (p < 0.05).
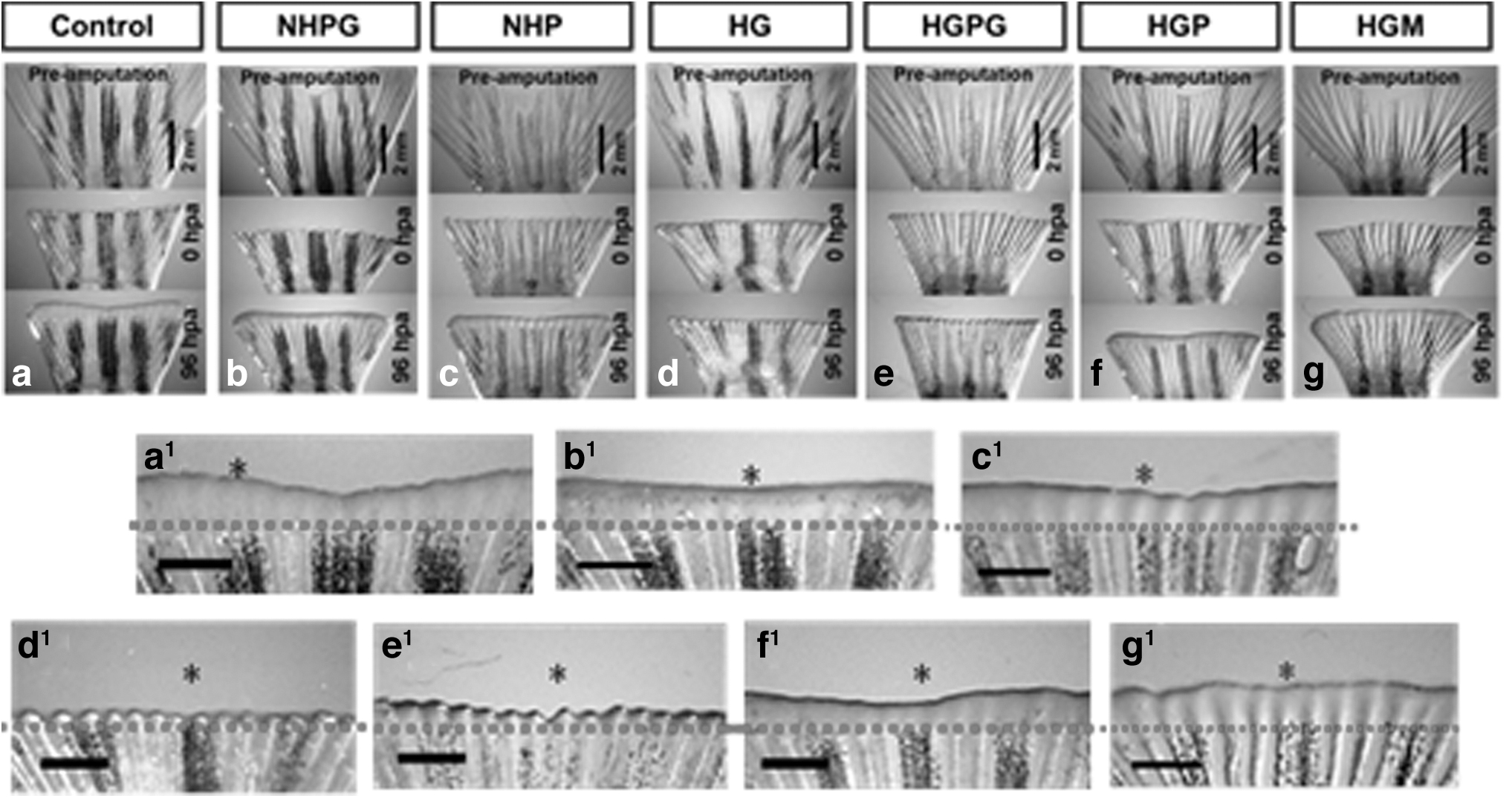
Caudal fin regeneration. The picture was taken using a stereomicroscope at preamputation, 0 hpa, and 96 hpa. The magnification of each group at 96 hpa are (a, b, c, d, e, f, and g). The asterisks (*) show the comparison of fin regeneration from all treatment groups to control. The scale bar shows 2 mm on (a–g) and 1 mm for (a1, b1, c1, d1, e1, f1, and g1). The red dashed line shows the amputation plane.
Hypoxia triggers transcription factor HIF-1α expression, as a response to oxygen deprivation, 73 leading to the activation of shha gene.74–76
The glucose excess in the caudal fin of blood vessels epithelial cells increases glucose metabolism and induces superoxide or ROS generation. Superoxide can inhibit GAPDH and increase methylglyoxal (MGO) accumulation, known as a glucose metabolite radical. 11 The higher levels of MGO will disrupt HIF-1α transcription factor stabilization, which leads to inhibition transcription of target genes such as shha. 76 This outcome is indicated in the decrease of shha expression in the zebrafish hyperglycemia treatment group (Fig. 5).
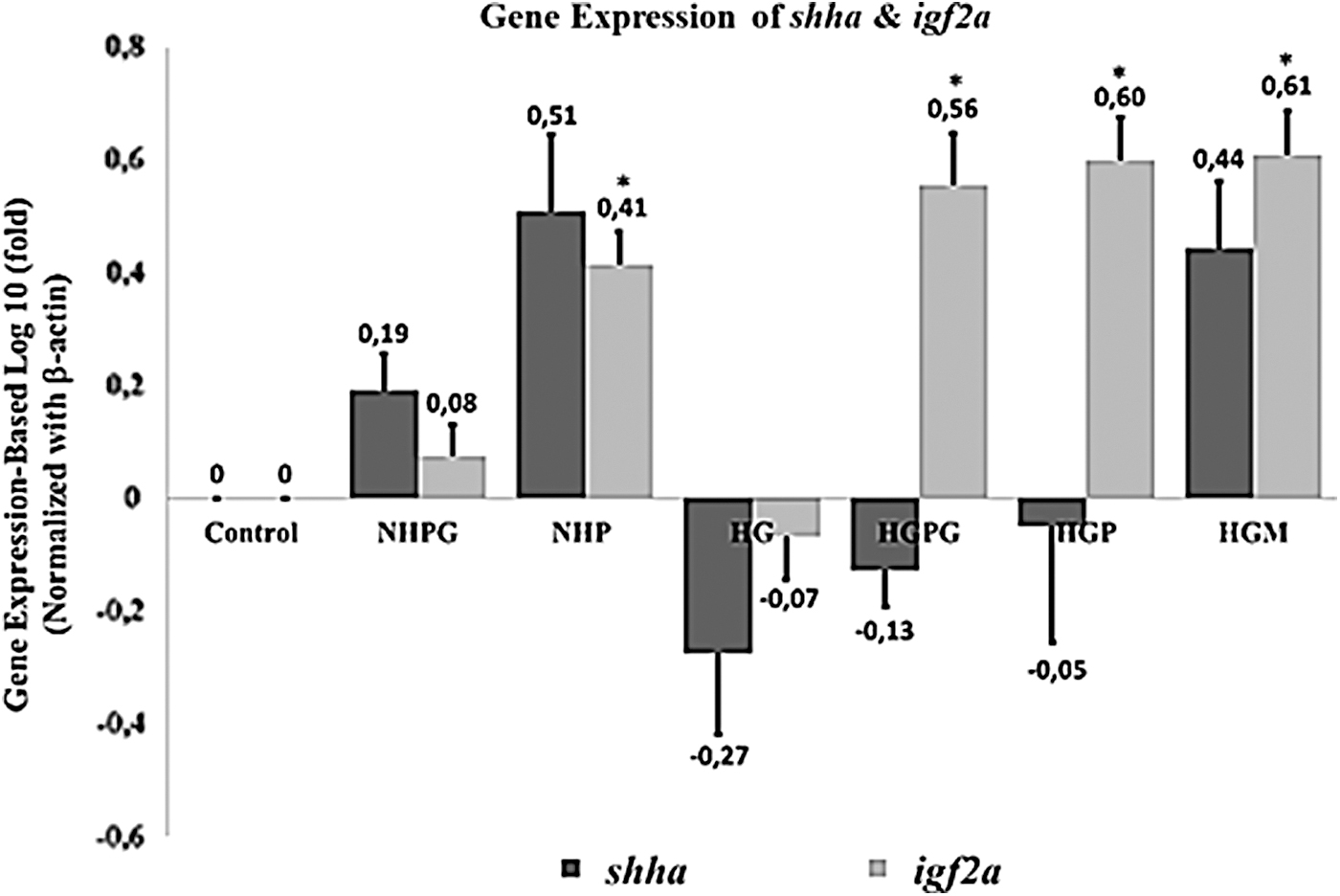
Expression profile of shha (sonic hedgehog-a) and igf2a (insulin-like growth factor 2a) genes on caudal fin blastema at 96 hpa. Three replicates were performed for each treatment,. Data are shown as mean ± standard deviation. Statistical analysis of the data used a one-way ANOVA followed by a post hoc Tukey HSD test. The asterisks (*) represent significance compared to the control group (p < 0.05). HSD, Honest Significant Difference.
The downregulation of the relative expression of the shha gene was accompanied by a downregulation of igf2a mRNA, known as one of the downstream target genes of HH signaling, 21 causing a decrease in the activity of cell proliferation, angiogenesis, and reepithelialization of hyperglycemia caudal fin regeneration compared to the control group. However, bmp2b may occur as a downstream target gene indirectly from Shh 51 (Fig. 6). Polyphenols can inhibit the activity of the NF-κβ transcription factor. 77 This transcription factor has been involved in the activation of IL-1β and other proinflammatory cytokines. 78 Therefore, propolis compounds are thought to be capable of being anti-inflammatory agents in zebrafish that are hyperinflammatory due to hyperglycemia. A study conducted by McLennan et al. 79 also showed that the administration of propolis to STZ-induced rats improved the wound healing process. This outcome was characterized by the normalization of neutrophil infiltration into the wound site. Ho-Jun et al. 80 also stated that the administration of phenolic compounds in the spleen cells of mice can reduce inflammatory cytokines IL-1, IL-6, and TNF-α mRNA expression levels.
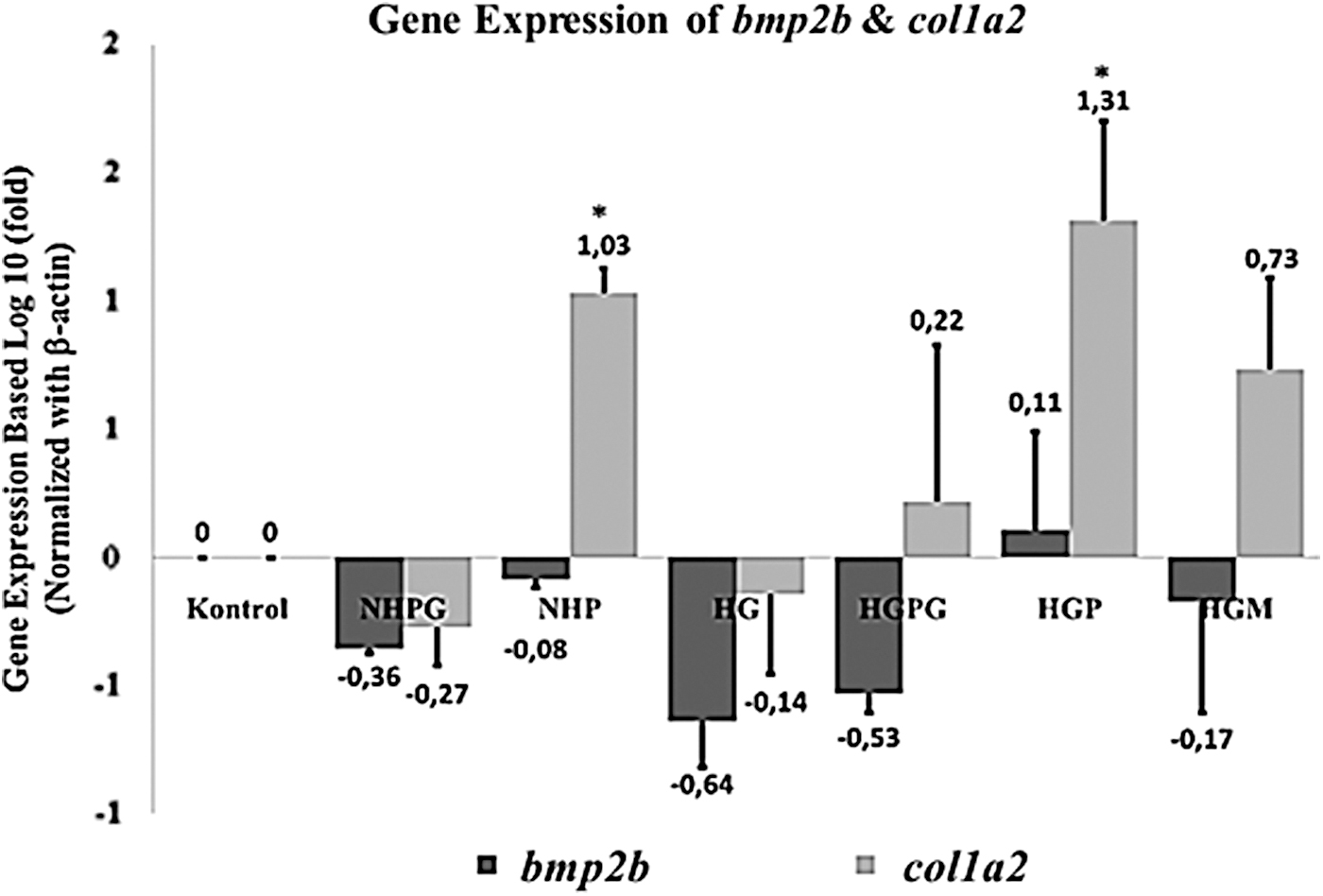
Expression profiles of bmp2b (bone morphogenetic protein 2b) and col1a2 (collagen type 1 alpha 2) genes on caudal fin blastema at 96 hpa. Three replicates were performed for each treatment. Data are shown as mean ± standard deviation. Statistical analysis of the data used a one-way ANOVA followed by a post hoc Tukey HSD test. The asterisks (*) represent significance compared to the control group (p < 0.05).
Polyphenols and flavonoid compounds of EEP are expected to work through the Aryl hydrocarbon receptor (AhR) and Nuclear factor-erythroid-2-related factor-2 (Nrf2) on cells involved in zebrafish caudal fin wound healing and regeneration. 81 AhR and Nrf2 are ligand-activated transcription factors that bind directly to the l exogenous ligand such as aromatic compounds (polyphenols and flavonoids), and serve as an essential transcription factor in various genes and detoxification cells against oxidative stress.81–83 With the support of these findings, we hypothesize that the transcriptional activation of genes involved in wound healing and regeneration of HIF-1α transcription factor may result in wound healing and enable the regeneration of zebrafish caudal fin.
Conclusion
Based on the results, phenols and flavonoids from EEP as an antioxidant are alleged to improve the caudal fin regeneration of HG zebrafish, which is characterized by an increase in fin regeneration and the relative gene expression of shha, igf2a, bmp2b, and col1a2.
Ethical Standards
All animal procedures research related to this study comply with Indonesian laws governing animal experiment.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was partly supported by grants (to IW), IPUPT Desentralisasi DIKTI Grant, Riset Kolaborasi, BPPDN DIKTI Scholarship and grants (to SE) PPKI 2019, and Insinas 2020.