
Abstract
The effects of manganese (Mn) toxicity in different organs and tissues in humans and other vertebrates have been studied since the beginning of the past century, but most of its cellular effects remain largely unknown. In this study, we studied the effects of Mn in zebrafish, at the cellular level, due to the transparent nature of zebrafish larvae that enables a powerful analysis under the light microscope. The collection of our results shows that environmental concentrations of 0.5 mg/L affect swim bladder inflation; at concentration of 50 and 100 mg/L Mn (1) induces alterations in viability, swim bladder, heart, and size of zebrafish larvae, (2) induces an increase in melanocyte area and the formation of cellular aggregates in the skin, and (3) induces an accumulation of β-Catenin in mesenchymal cells in the caudal fin of zebrafish larvae. Our data suggest that increased levels of Mn induce cell aggregate formation in the skin and the presence of more melanocytes in the zebrafish caudal fin. Interestingly, the adhesion protein β-Catenin was activated in mesenchymal cells near the cell aggregates. These results open important new questions on the role of Mn toxicity on cellular organization and β-Catenin responses in fishes.
Introduction
Manganese (Mn) is a trace metal and a cofactor to enzymes with essential biological functions in nearly all human tissues, including metabolism, growth, replication, and differentiation.1,2 However, exposure to high levels of Mn can lead to severe pathologies, where neurological disorders are the most evident.3–6 Importantly, there is still a void in knowledge regarding the effects of Mn in tissues other than the nervous system, particularly using analysis at the molecular and cellular levels. One of the most promising vertebrate animal models for studies of human-related diseases is the zebrafish (Danio rerio). The many advantages of zebrafish include its well-characterized genome and its excellent optical clarity, which is well suited for studies on early stages of embryonic development.
Zebrafish has been widely used to investigate the effects of Mn, including a manganism model.3,7–13 It has been shown that Mn content levels increase in the zebrafish after 5 days postfertilization (dpf), when they start to feed from external sources. 14 Interestingly, at this developmental stage the larvae increase sensitivity to Mn exposure.3,8,10,12 Nevertheless, none of these studies analyzed the cellular effects of Mn during zebrafish early developmental stages. Although zebrafish get more sensitive after 5 dpf, we have used early stages until 96 hours postfertilization (hpf) larvae to investigate in more detail the complexity of larvae phenotypes exposed to Mn. We found that at concentrations of 0.5 mg/L, five times above the acceptable for potable water in Brazil, 15 detected in polluted environments,16–18 the zebrafish embryos presented a noninflated swim bladder.
Moreover, we describe interesting phenotypes besides the neurological effects in the literature, such as the formation of skin aggregates, cellular infiltration, and increased number of melanocytes. Therefore, we aimed to analyze the cellular effects of Mn during zebrafish development using high-quality light microscopy and confocal fluorescence microscopy. Mn induced an increased expression of the signaling protein β-Catenin on cells beneath the skin aggregates with mesenchymal morphology. Our data suggest a disruptive effect of Mn on skin cellular organization and the regulation of Wnt/β-Catenin on mesenchymal cells that were not previously described in the literature.
Materials and Methods
Zebrafish husbandry and embryo exposure to Mn
Zebrafish (Danio rerio) were maintained in aquaria with recirculating water system at 28°C ± 1°C on a 14:10 light/dark cycle in animal facility at the Institute of Biodiversity and Sustainability (NUPEM) of the Federal University of Rio de Janeiro, Macaé-RJ, Brazil. 19 Animals were handled and experimented according to Institutional Animal Care and Use Committee protocols under the number 067/17. Embryos and larvae were collected, and the treatment started at 8–1 k cell-stage with increasing concentrations of MnCl2 (0.5, 5, 50, 100, and 500 mg/L) up to 96 hpf. Twenty-four-well plates were used for the experiments. Five embryos were distributed per well containing 2 mL of solution. Each MnCl2 concentration used 4 replicates (20 embryos/concentration/experiment). In total, the experiment was repeated three times.
Sudan Black labeling
Sudan Black (SB) labeling protocol was modified from previous studies.20–22 Control and Mn-treated zebrafish larvae at 96-hpf stages were fixed in 4% paraformaldehyde overnight. Next, larvae were washed five times with phosphate buffered saline (PBS) and incubated with 0.3% SB Solution (dissolved in ethanol, phenol, and Na2HPO.12H2O) for 20 min. Then, larvae were washed five times with 70% ethanol and incubated for 15 min with 1% KOH, 1%H2O2, and EtOH 70%. Larvae were incubated for 10 min with PBS containing 0.1% Tween 20 and 50% ethanol, followed by 10 min with 30% ethanol and digital images of larvae were obtained in a light microscope.
Antibodies and probes
Rabbit polyclonal antibody against β-Catenin (code no. C-2206) was from Sigma-Aldrich. DNA-binding probe 4,6-diamidino-2-phenylindole dihydrochloride (DAPI) and Alexa Fluor 546-goat anti-rabbit IgG antibody was from Molecular Probes.
Immunofluorescence
Dechorionated zebrafish larvae at 96 hpf were fixed in 4% paraformaldehyde in PBS for 1 h at room temperature. Larvae were then permeabilized with 0.5% Triton-X 100 in PBS (PBS/T) three times for 30 min and incubated overnight at 4°C with primary antibodies (all diluted 1:100 in PBS/T). Then, larvae were washed for 30 min with PBS/T and incubated for 1 h at 37°C with Alexa Fluor-conjugated secondary antibodies (all diluted 1:200 in PBS/T). Nuclei were labeled with 0.1 μg/mL of DAPI in 0.9% NaCl. Larvae were mounted on #1.5 24 × 60-mm glass coverslips (with spacers) using Prolong Gold (Molecular Probes). Experiments with zebrafish larvae were repeated four times.
Fluorescence image acquisition and processing
Zebrafish larvae were examined in an Axiovert 100 microscope (Carl Zeiss, Germany) coupled to an Olympus DP71 high-resolution camera, and in a DSU Spinning Disk confocal scanner mounted on an inverted fluorescent microscope (Olympus, Japan) and TCS SPE confocal (Leica). For live imaging, live embryos were mounted in 1% methyl cellulose and bright-field microscopy was performed on a Leica stereomicroscope (Leica, Germany). Image processing (brightness, contrast adjustments, and area quantification) was performed using Fiji software 23 and figure panels were mounted with Adobe Photoshop software (Adobe Systems, Inc.).
Statistical analysis
Statistical analysis was carried out using the GraphPad Prism software version 8. The results are expressed as mean ± standard error of the mean. The groups were performed with at least five independent experiments (n = 5), where ****p < 0.0001, ***p < 0.001, **p < 0.01, and *p < 0.05 versus control group. Statistical analysis was performed using one-way analysis of variance.
Results and Discussion
Analysis of mortality, hatching, and alterations in swim bladder and heart after in zebrafish larvae after manganese treatment
To study the effects of manganese (Mn) in zebrafish embryos, we first analyzed the mortality rate (Supplementary Fig. S1) of zebrafish larvae exposed to different concentrations of Mn (0.5, 5, 50, 100, and 500 mg/L). The mortality rate for all concentrations until 96 hpf were <10% (Supplementary Fig. S1). It is comparable with what has been described in the literature, where the mortality to Mn exposure increase after 96 hpf.3,8,10,12 However, the hatching rate at 500 mg/L was significantly impaired with strong morphological retardation of the embryos at 96 hpf (Fig. 1A, I). It is possible that nonhatching embryos exposed to 500 mg/L (9 mM) would not survive after 5 dpf. If we consider the LC50 of embryo-larvae exposed to MnCl2 for 48 h after 3.5 dpf the LC50 was of 2.0 mM. 8 In another study, the embryos exposed from 48 to 120 hpf had an LC50 range of 6.0–10 mM. 10
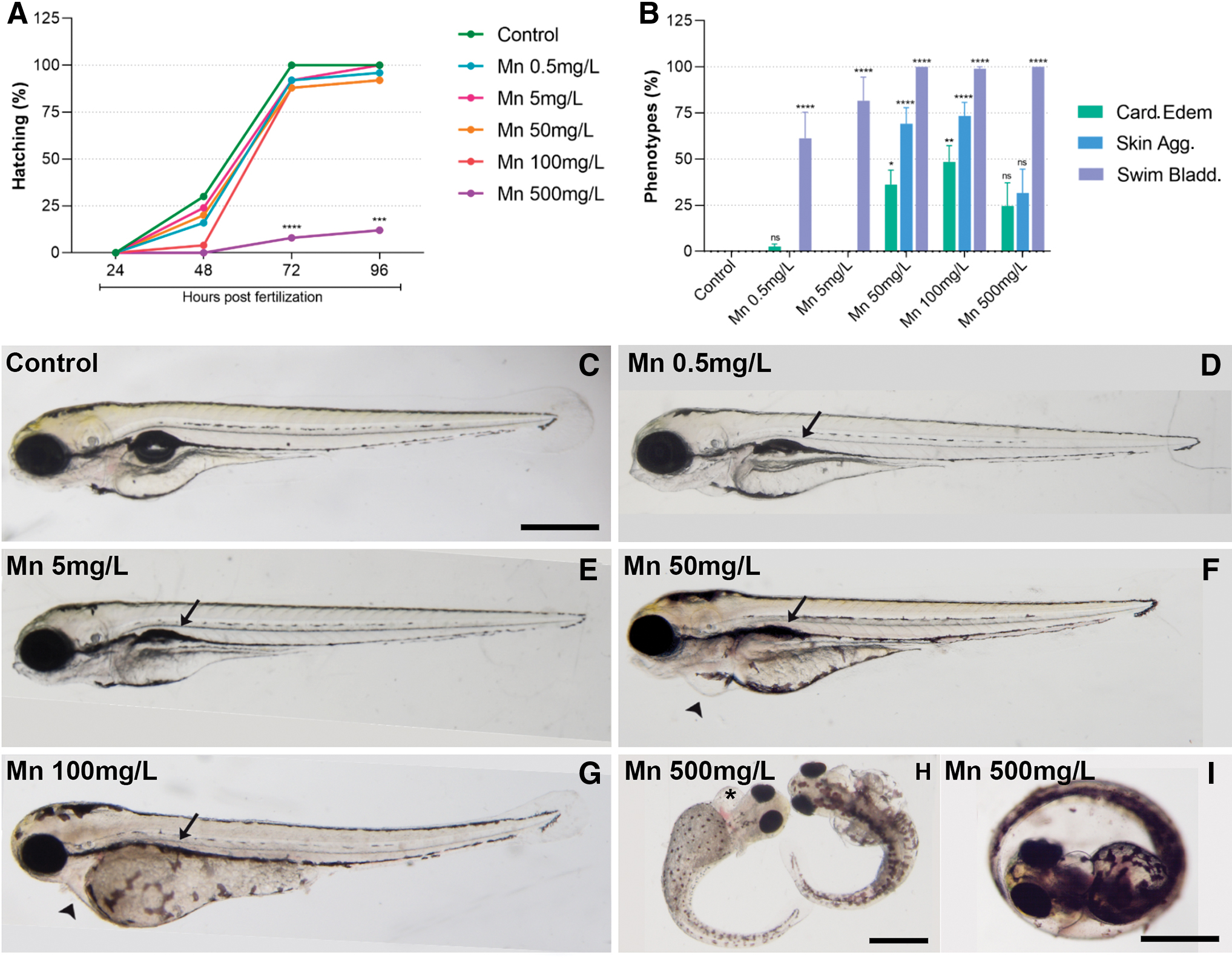
Mn treatment affects hatching and induces heart, swim bladder, and skin alterations. Zebrafish eggs were treated with Mn at 0.5, 5, 50, 100, and 500 mg/L and analyzed under a light microscope. Only 500 mg/L of Mn showed a significant inhibition of hatching
Regarding the morphological phenotypes, apart from neurological and craniofacial morphological malformations observed in the literature,3,8,12 we found alterations in swim bladder, heart, and cell aggregates on the skin of zebrafish larvae. Swim bladder fails to inflate in 60% of the larvae in 0.5 mg/L and 77% in 5 mg/L (Fig. 1). Whereas all embryos exposed to 50, 100, and 500 mg/L of Mn had an effect on swim bladder inflation (Fig. 1C–I). The Mn concentration exposure started with 0.5 mg/L that is similar to what has been found in polluted environmental samples16–18 and is above the 0.1 mg/L permitted in potable water in Brazil. 15 The swim bladder inflates at 4.5 dpf when larvae are of 4.4 mm length.24,25 It is a gas-filled organ that is important for fish buoyancy and has similarities to tetrapod lungs.24,25 These data reinforce the importance of keeping the Mn levels in the water <0.5 mg/L.
Another phenotype noticed in zebrafish embryos was cardiac edema, which was detected in 30% of the larvae in 50 mg/L, 50% in 100 mg/L, and 25% in 500 mg/L (Fig. 1F–I). Indeed, Mn has been shown to cause heart disturbance in many vertebrates, including humans.26,27 Workers exposed to Mn present higher alterations in electrocardiogram than control group, caused by different types of cardiac alterations. 27 In zebrafish embryos, Mn induces calcium decrease 10 and pericardial edema. 12 Besides calcium dysregulation leads to failure of heart development in zebrafish, 26 calcium itself has important roles along zebrafish development. 28
In case calcium signaling disruption in the embryo would be proportional to Mn concentration, it could mean that as higher the concentration of Mn, more calcium imbalance would be generated. This could be one of the explanations why the embryos exposed to 500 mg/L had a lower percentage of heart edema than the ones exposed to 100 mg/L, including its strong impairment in development (Fig. 1I). However, this should be further investigated in future study.
Mn treatment induces an increase in melanocyte area and the formation of skin aggregates in zebrafish larvae
Larvae treated with 500 mg/L Mn had severe development impairment; therefore, we carried out the detailed phenotype investigation of 50 and 100 mg/L Mn-treated embryos. At 96 hpf, the larvae had a significant decrease in the total length after 100 mg/L of Mn (6% decrease compared with untreated larvae) (Supplementary Fig. S2).
Further analysis of zebrafish larvae caudal fin size under the light microscope, exhibited an extended area of melanocyte occupation after Mn exposure. Quantification of the melanocyte area showed an 50% increase after 50 mg/L Mn, whereas no change was observed after 100 mg/L Mn (Fig. 2). Interestingly, Mn has a high affinity for melanin tissues in vitro and in vivo 29 and accumulate in specific regions of the brain, especially in pigmented neurons of substantia nigra. 30 The toxicity of Mn in the nervous system has been widely described, but little data are related to skin melanocytes. There is an indication in the literature that human melanocytes viability decreases in a dose-dependent manner upon Mn exposure. 31 However, growth and lethality of melanocytes in response to Mn need further investigation. Mn in the brain induces mitochondrial impairment that leads to a series of events ending in apoptosis induction (reviewed in Pajarillo et al.). 32 Would it be the same for other melanin-containing cells?
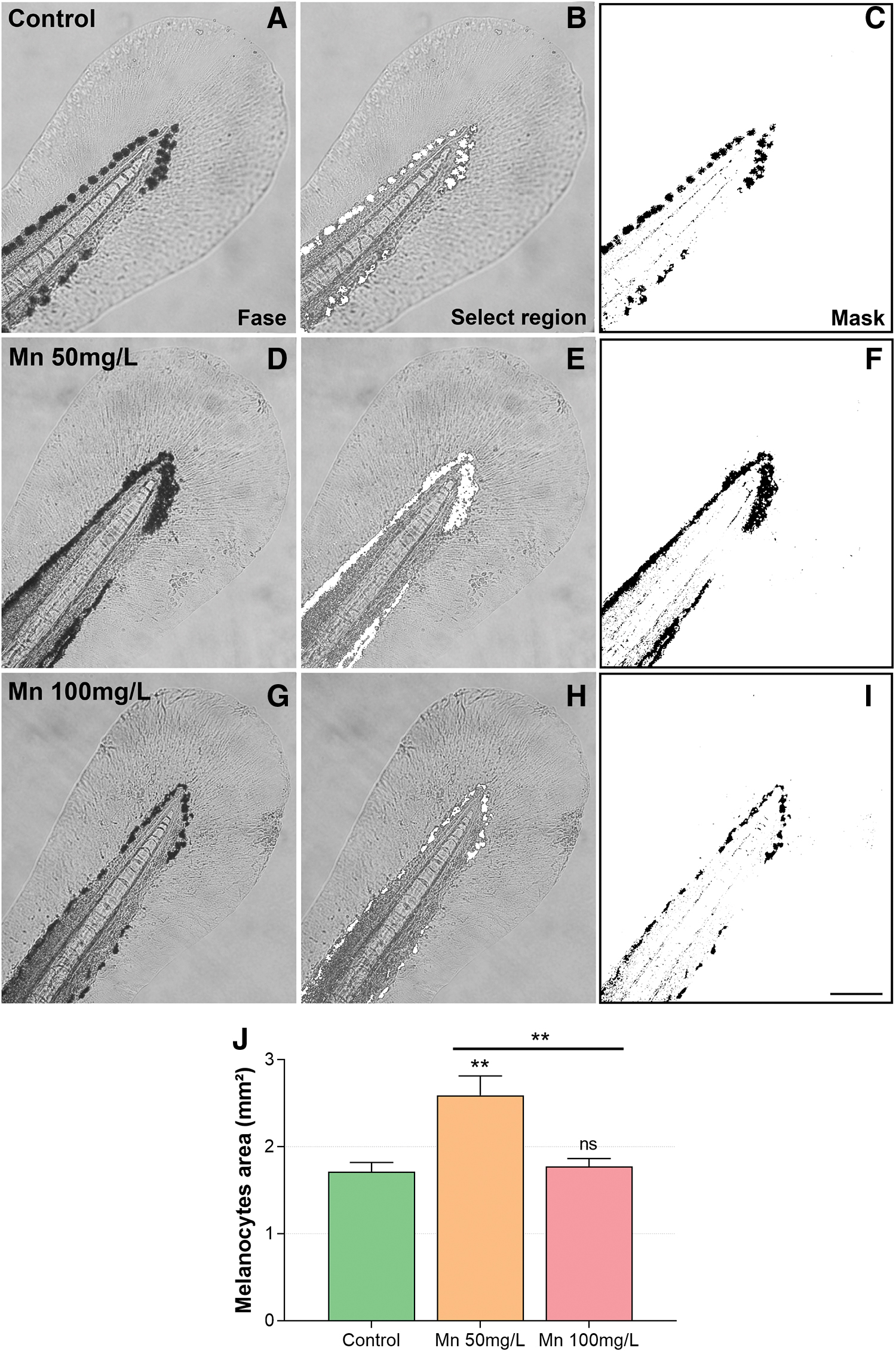
Mn treatment increases the area of melanocytes at the zebrafish fin. Zebrafish larvae were treated with Mn at 50 or 100 mg/L
We analyzed another group of cells that presented abnormal behavior that were not previously described in the literature. One of the unexpected phenotypes found was the presence of skin aggregates on the larvae. We found a 1.5-fold increase in skin aggregates in the larvae treated with 50 mg/L Mn (Fig. 3). The cellular aggregates were clearly visible under the light microscope without any kind of labeling or staining. Quantification of the size of the cellular aggregates showed that they have an average size of 1.53 mm2 after 50 mg/L of Mn and 0.43 mm2 in 100 mg/L Mn (Fig. 3).
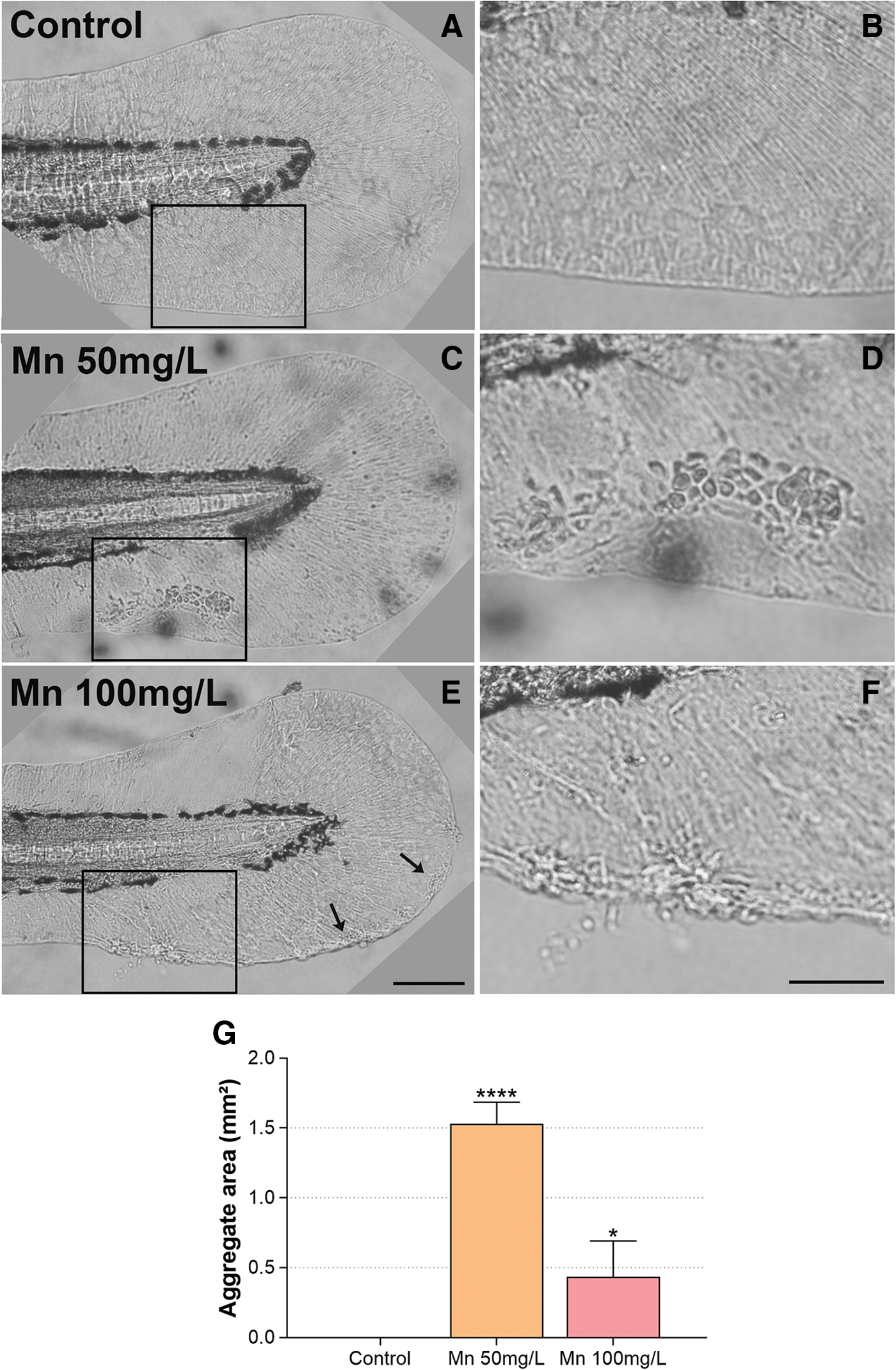
Mn treatment induces the formation of cellular aggregates at the zebrafish fin. Zebrafish larvae were treated with Mn at 50 or 100 mg/L
Previous reports of Mn toxicity in zebrafish larvae did not cite those skin aggregates. The tail fin is a very fragile structure. Possibly the larvae preparation used for analyses smashed the tail fins and this phenotype was overlooked.8,10,12 It is known that Mn-deficient diets in young men lead to skin rashes. 33 In contrast, patients exposed to MnCl2 presented irritant reactions and it was cytotoxic to keratinocytes cell culture, 34 suggesting an important role of Mn in skin homeostasis. Ionic imbalance in zebrafish atpase1b1a mutant, a mutation in Na,K-ATPase β1-subunit, demonstrated the formation of skin aggregates derived from the proliferation of basal keratinocytes with metastasis capacity. 35
The skin aggregates after Mn exposure were morphologically very similar to atpase1b1a mutant. Mn affect the expression and/or function of other atpases as ATP13A2 and ATP2C1 and calcium balance.36–38 The aggregates from atpase1b1a mutants produced metalloproteinase-9 (MMP) capable of degrade basement membrane and is a malignancy marker. 35 Mn induces MMP9 expression in human keratinocytes 39 and rat astrocytes. 40 Future experiments should characterize if the skin aggregates have similar malignant characteristics as the ones formed by ionic disturbances.
Another important characteristic of Mn toxicity is the induction of an inflammatory environment, inducing interleukin-6 and tumor necrosis factor α.34,41,42 To further analyze for cell infiltrates possibly induced by inflammation, we have stained 96 hpf larvae with SB.20,21 SB stain rich lipid organelles of granulocytes and has been characterized in zebrafish to stain myeloperoxidase-positive granules of neutrophils. 20 SB-positive cells were increased in the caudal fin of Mn-treated embryos (Fig. 4A). SB stained the cellular infiltrates in the caudal fin of zebrafish larvae, and the area of aggregates had SB-positive cells after Mn treatment, as compared with untreated larvae (Fig. 4), indicating an inflammatory response.
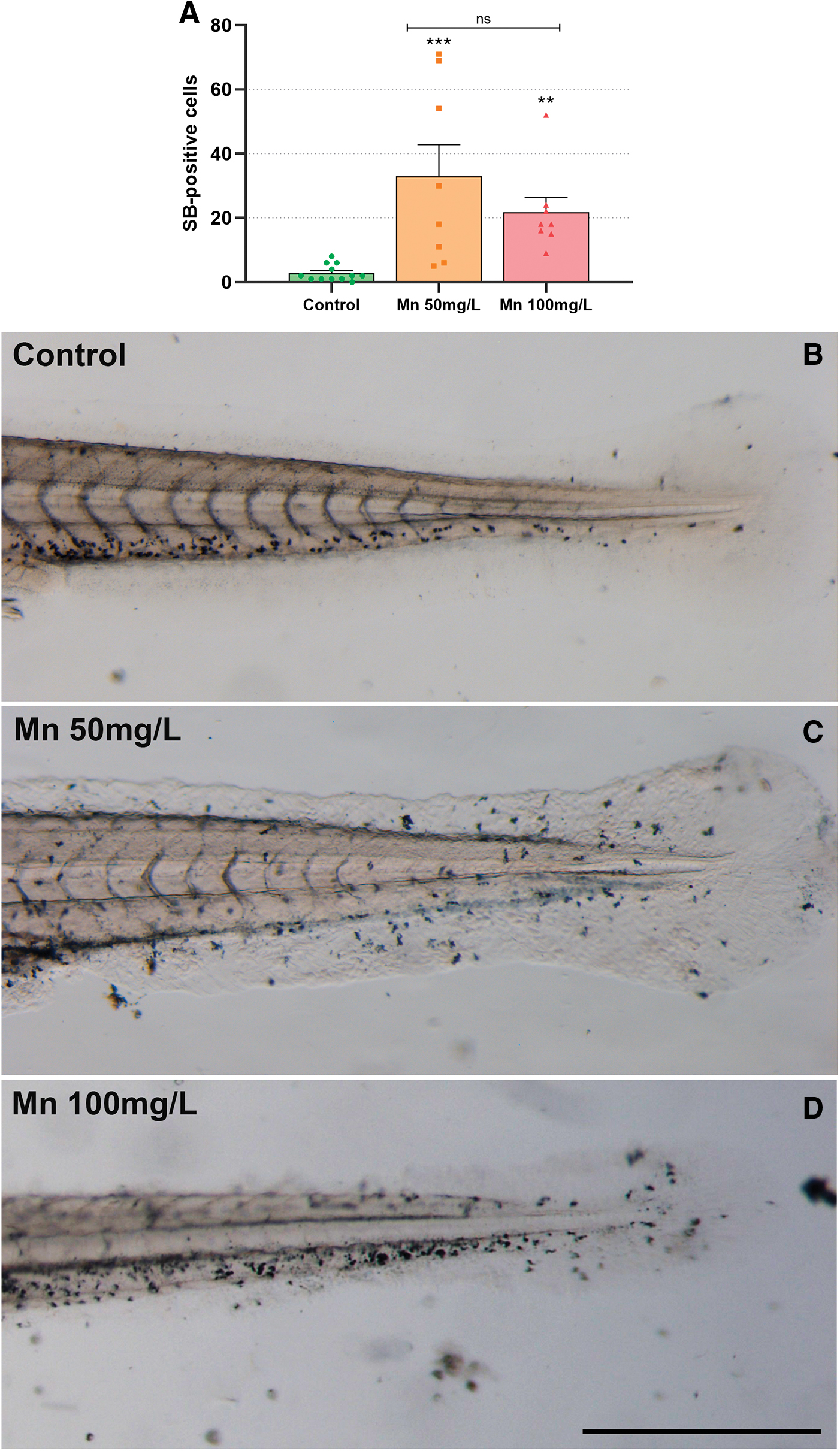
Mn treatment increases the number of SB-positive cells at the zebrafish fin. Zebrafish larvae were treated with Mn at 50 or 100 mg/L, stained with SB and analyzed under a light microscope
Mn treatment induces an accumulation of β-Catenin in aggregates in the caudal fin of zebrafish larvae
The disturbances in cell organization in the caudal fin of zebrafish larvae after Mn treatment raise the idea to analyze the distribution of proteins related to tissue structure. β-Catenin is a structural and signaling protein that is essential for the generation and maintenance of epithelial tissues organization. 43 β-Catenin localization in the nucleus indicates activation of the canonical Wnt/β-Catenin pathway. 43 Immunofluorescence labeling of β-Catenin and the nuclear dye DAPI showed β-Catenin in 96 hpf larvae control were more diffuse along the skin cells (Fig. 5A, B).
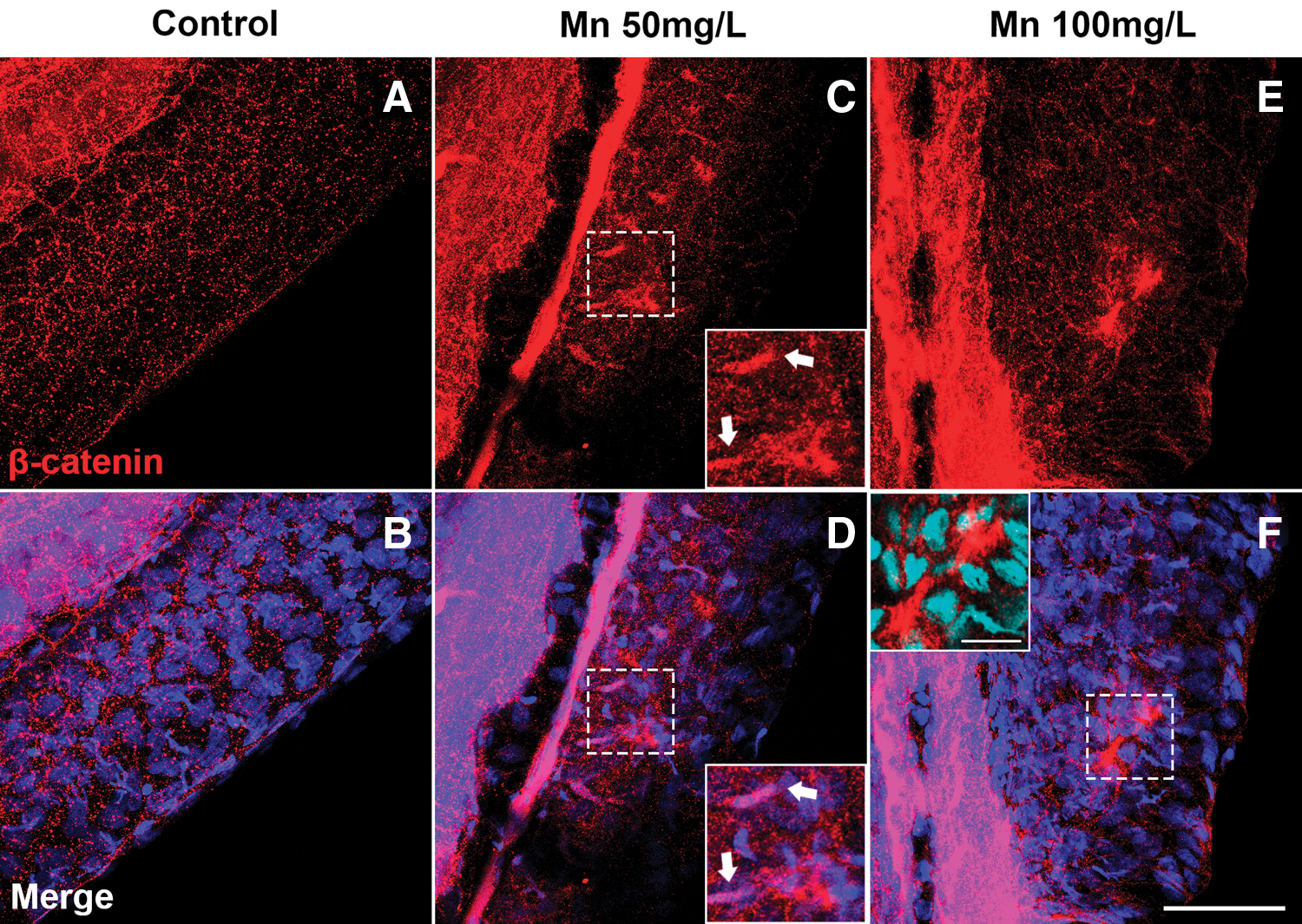
Mn treatment disorganizes the epithelium and induces the expression of β-Catenin in the caudal fin. Confocal analysis of zebrafish larvae at 96 h postfertilization stained with β-Catenin (red) and DAPI (blue). In the control embryo, β-Catenin is diffused at typical intercellular adherens junctions of epithelial cells
However, in Mn-exposed embryos, β-Catenin was concentrated in cells beneath the caudal fin aggregates; and not in the cell aggregates (Fig. 5C–F). In higher magnification, it is possible to see that β-Catenin-positive cells have an elongated morphology, resembling mesenchymal migrating cells below the skin in the caudal fin (Fig. 5C–F) as seen in migrating mesenchymal cells in the fin during development. 44 However, the staining is not all over the dermal-mesenchymal cells that migrate into the caudal fins, 44 they appear to be a specific type of migrating cells. It is possible that they could be myeloid migrating cells.
Further investigation on the nature of these positive-β-catenin cells should be analyzed. However, it is exciting to think that there is a cross talk between nuclear factor kappa B (NFκB) and Wnt/β-Catenin in inflammatory events, acting positively or negatively. 45 Recent literature demonstrates that ficetin, a flavonoid, activated β-Catenin in zebrafish larvae and induced melanogenesis 46 and in macrophages the β-Catenin activation can inhibit the NFκB signaling, acting as an anti-inflammatory. 47 This leads to future questions to be answered on the role of Mn toxicity in β-Catenin activation in mesenchymal cells with the potential to act as an inflammatory mediator in skin damage.
Conclusions
In this study, we studied the effects of Mn during zebrafish development and found that concentration of 50 and 100 mg/L of Mn (1) induces alterations in swim bladder inflation, heart, and size of zebrafish larvae, (2) induces an increase in melanocyte area and the formation of cellular aggregates in the caudal fin, and (3) induces an accumulation of β-Catenin in mesenchymal cells in the caudal fin of zebrafish larvae. These results open important new questions on the Mn role in skin cell organization, melanocyte behavior, and the multifunctional protein β-Catenin in the cellular responses in zebrafish.
Data Availability Statement
The data sets generated for this study can be found within the article figures.
Ethics Statement
The use of zebrafish embryos was approved by the Ethics Committee for Animal Care and Use in Scientific Research from the Federal University of Rio de Janeiro and received the approval number 067/17.
Consent to Publish
All listed authors consent and agree with the publication of the data present within this article. The authors declare that they have no conflict of interest.
Footnotes
Authors' Contributions
N.M.F. conceived the study. G.G. and J.L.O. contributed to the acquisition of data. C.M., G.G., J.L.O., M.C., and N.M.F. contributed to the analysis and interpretation of data. C.M. and N.M.F. wrote the first draft of the article. All authors consent to participate, contributed to article revision, read, and approved the submitted version.
Disclosure Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
This study was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, funding number 302115/2017-0 for CM, 301443/2018-1 for MLC, 431352/2018-6 for NMF) and Fundação de Apoio à Pesquisa do Estado do Rio de Janeiro (FAPERJ, funding number E-26/202.920/2019 for CM and E26/210.220/2018 for MLC).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.