
Abstract
Background Increased numbers of circulating endothelial progenitor cells (EPC) are associated with improved vascular function. Exercise is a central component of the primary prevention of vascular diseases. The effect of physical activity on circulating EPC in healthy individuals is not known.
Design A prospective crossover study.
Methods and results In order to study a potential link between the extent of physical exercise and progenitor cells in humans, EPC were quantified by flow cytometry and cell culture in 25 healthy volunteers undergoing three protocols of running exercise. Intensive running, defined as 30 min at 100% of the velocity of the individual anaerobic threshold (IAT; ∼82% maximal oxygen consumption; VO2max), as well as moderate running with 30 min at 80% of the velocity of the IAT (∼68% VO2max), increased circulating EPC numbers to 235±93% and 263±106% of control levels, respectively. However, moderate short-term running for 10 min did not upregulate EPC counts. The maximum increase in circulating EPC numbers was observed 10–30 min after intensive running. Exercise increased EPC migratory and colony-forming capacity.
Conclusions Intensive and moderate exercising for 30 min, but not for 10 min, increased circulating levels of EPC, which may represent an important beneficial outcome of physical exercise. The data support the notion that increased numbers of EPC correlate with cardiovascular health and suggest EPC quantification as a novel surrogate parameter of the vascular effects of exercising.
Introduction
Recent evidence has shown that cardiovascular function is significantly modulated by circulating premature cells derived from the bone marrow [1]. A subset of these stem cells named endothelial progenitor cells (EPC) [2] enhances angiogenesis, promotes vascular repair and improves endothelial function [3–10]. The circulating numbers of EPC are regulated. Whereas myocardial infarction and bypass surgery acutely increase the circulating numbers, vascular risk factors have been shown to reduce EPC, suggesting that vascular health and also repair processes after injury require increased numbers of this potentially beneficial population of cells [4, 5, 11, 12].
Physical activity is associated with a decrease in cardiovascular events, and therefore is recommended as a central component of the primary prevention of vascular diseases [13–15]. Physical training improves endothelial function, exercise capacity and collateralization in patients with coronary artery disease [16, 17], chronic heart failure [18] and peripheral artery disease [19]. However, the molecular effects of physical exercise are only partly understood. Individual advice regarding the duration and intensity of physical exercise remains an important clinical problem [14, 20], and there is need for novel surrogate parameters to determine the vascular effects of specific training programmes. In a recent experimental study, running exercise increased EPC numbers in mice by a mechanism related to the bioavailability of nitric oxide [21]. In patients with vascular risk factors or established coronary artery disease, the numbers and function of EPC are impaired [22]. Recent data show that EPC increase after exercise-induced ischaemia or exhaustive symptom-limited dynamic exercise in these patients [23, 24]. Furthermore, a 4-week exercise programme was associated with an upregulation of circulating CD34+/vascular endothelial growth factor receptor 2-positive (VEGFR-2+) EPC in elderly patients with documented coronary artery disease [21].
However, it is not known whether physical exercise directly affects the number of circulating EPC in the absence of vascular disease in humans. Importantly, the intensity and duration of training needed to regulate EPC are unknown, as well as the time course of EPC release after exercise. Therefore, we hypothesized that physical exercise directly upregulates the number and function of circulating EPC in healthy individuals. In addition, we compared the effects of three well-defined running protocols and studied the kinetics of EPC regulation.
Methods
Study population
We tested the effect of running exercise on EPC in 25 healthy, non-smoking, male volunteers. Participants were moderately endurance-trained students at the medical school or the school of sports medicine at our institution or participants of a local recreational running club. The typical amount of training was at least as recommended by the American Heart Association, four to five times a week for 30 min. Their maximal oxygen consumption (VO2max) and individual anaerobic threshold (IAT) corresponded to this level of physical activity. Subjects had no medical history, no vascular risk factors and had no medical treatment. Physical examination, echocardiogram (ECG) at rest as well as during exercise and venous blood sampling excluded apparent pathologies (for physical characteristics of the subjects see Table 1).
Determination of individual anaerobic threshold
A maximal incremental graded treadmill ergometry (starting with 2.5 m/s, increasing by 0.5 m/s, 3-min stages) was performed until volitional exhaustion. Capillary blood samples were taken from the hyperaemized earlobe at rest, immediately after each workload, immediately, 1, 3, 5 and 10 min after cessation of exercise for enzymatic determination of lactate. Heart rates were determined by six-lead ECG. The IAT was determined from the lactate-workload plot [25, 26]. The IAT indicates the exercise intensity corresponding to the maximal lactate steady state, and represents an established concept for the determination of intensities for endurance training [26]. VO2max was calculated using standard values.
Protocols of running exercise
The volunteers underwent three predefined controlled running protocols on a 400 m track in random crossover: intensive running defined as 30 min at 100% of the velocity of the IAT (corresponding to approximately 82% VO2max), moderate running with 30 min at 80% of the velocity of the IAT (corresponding to approximately 68% VO2max), and moderate short-term running with 10 min at 80% of the velocity of the IAT (n = 10; corresponding to approximately 68% VO2max). The time between protocols was more than one and less than 5 weeks, each running exercise was started between 0630 and 0800 h. Immediately before and 10 min after exercise venous blood samples were drawn. In a subgroup of six volunteers, blood samples were taken before, and 10 min, 30 min, 2 h, 6 h, and 24h after exercise.
Flow cytometry
Mononuclear cells were selected using Ficoll densitiy gradient centrifugation (Biocoll Separating Sol.; Biochrom, Berlin, Germany) from 20ml human blood drawn in sodium citrate and resuspended in 100 μl endothelial cell basal medium (EBM, CellSystems, St Katharinen, Germany) with supplements (1 μg/ml hydrocortisone, 3 μg/ml bovine brain extract, 30 μg/ml gentamicin, 50 μg/ml amphotericin B, 10μg/ml human epidermal growth factor, 20% fetal calve serum). Cells were labeled with 10 μl of each IgG 2a FITC and IgG 2a phycoerythrine (PE; Pharmingen, Heidelberg, Germany) for isotype controls, 10 μl of each CD34 FITC, CD117 PE (Becton Dickinson, Heidelberg, Germany), CD133 PE (Miltenyi, Bergisch-Gladbach, Germany), 5 μl goat-anti-human VEGFR-2 (R&D Systems, Wiesbaden, Germany) labeled with 50 μl anti-goat streptavidin PE in 1:10 solution (Dako, Hamburg, Germany). All antibodies except for VEGFR-2 were added directly to the cell suspension and kept for 20 min in the dark at room temperature. Cells were washed three times in cell-wash and 2% paraformaldehyde. For preparation of the VEGFR-2 antibody 6 μl of human immunoglobuline (Polyglobulin 10%; Bayer, Wuppertal, Germany) was added as a blocking reagent to the cell suspension and incubated for 30 min on ice. For primary unconjugated antibodies, cells were incubated in 6 μl 0.5% goat serum for 20 min on ice. Staining with streptavidin PE was performed for 30 min on ice in the dark. The samples were washed twice with FACS buffer containing phosphate buffer solution, 0.1% bovine albumin and aprotinin (20 μl/ml). To complete the staining procedure the other antibodies were added following the protocol. Cells were washed twice with buffer and resuspended in 200 μl of each cell wash and 2% paraformaldehyde. Cell fluorescence was measured by flow cytometry using a FACSCalibur (Becton Dickinson) and the data were analysed using the Cellquest software (Becton Dickinson). A total of 10 × 104 events were analysed.
Culture of circulating human endothelial progenitor cells
An aliquot of 1 × 107 mononuclear cells were cultured on fibronectin-coated dishes in endothelial basal medium (CellSystems) [9]. After 4 days in culture, adherent cells were incubated with 1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine-labeled acetylated low-density lipoprotein (DiLDL, 2.4 μg/ml, CellSystems) and stained using FITC labeled Ulex europaeus agglutinin I (lectin, 10 μg/ml) (Sigma, Hamburg, Germany). Double-positive cells were counted manually by two independent observers blinded to the study. For determination of colony-forming units, 1 × 106 non-adherent cells were transferred after 48 h into a new fibronectin-coated well. Growth medium was changed every 3 days and clusters of more than 15 cells were counted after 7 days.
Migration assay
The migratory capacity of EPC was determined in a modified Boyden chamber. EPC were harvested by centrifugation, resuspended in 500 μl EBM with supplements, counted and placed in the upper chamber of a modified Boyden chamber (1 × 105 cells/per chamber; BD Biosciences, Heidelberg, Germany). The chamber was placed in a 24-well culture dish containing EBM with supplements and vascular endothelial growth factor (VEGF) (50 ng/ml). After incubation at 37°C for 24h, the lower side of the filter was washed, fixed and incubated with DiLDL. Migrated cells at the lower part of the filter were counted manually in four random microscopic fields.
Serum cortisol and vascular endothelial growth factor
Serum cortisol levels were determined by luminescence-enzyme immunoassay (Access; Beckman-Coulter, Krefeld, Germany). Serum VEGF was measured by high-sensitivity enzyme-linked immunosorbent assay (Quantikine; R&D Systems, lower detection limit 9.0 pg/ml).
Statistical analysis
Results for continuous variables are presented as mean ± SD. Wilcoxon test and analysis of variance for multiple comparisons were employed when applicable. Post-hoc comparisons were performed using the Neuman–Keuls test. Probability values of P < 0.05 were considered significant.
Results
The characteristics of the young and physically active study population and selected blood values are summarized in Table 1. Serum lactate increased after intensive exercise (30 min, 100% IAT) from 1.3 ± 0.5 at baseline to 5.1 ± 1.7 mmol/l (P < 0.01) (Fig. 1). Moderate running (80% IAT) for 30 min or short-term moderate exercise for 10 min did not significantly alter lactate levels (30 min: 1.2 ± 0.5 to 1.6 ± 0.6 mmol/l; 10 min: 1.3 ± 0.67 to 1.7 ± 0.6 mmol/l).
Patient characteristics
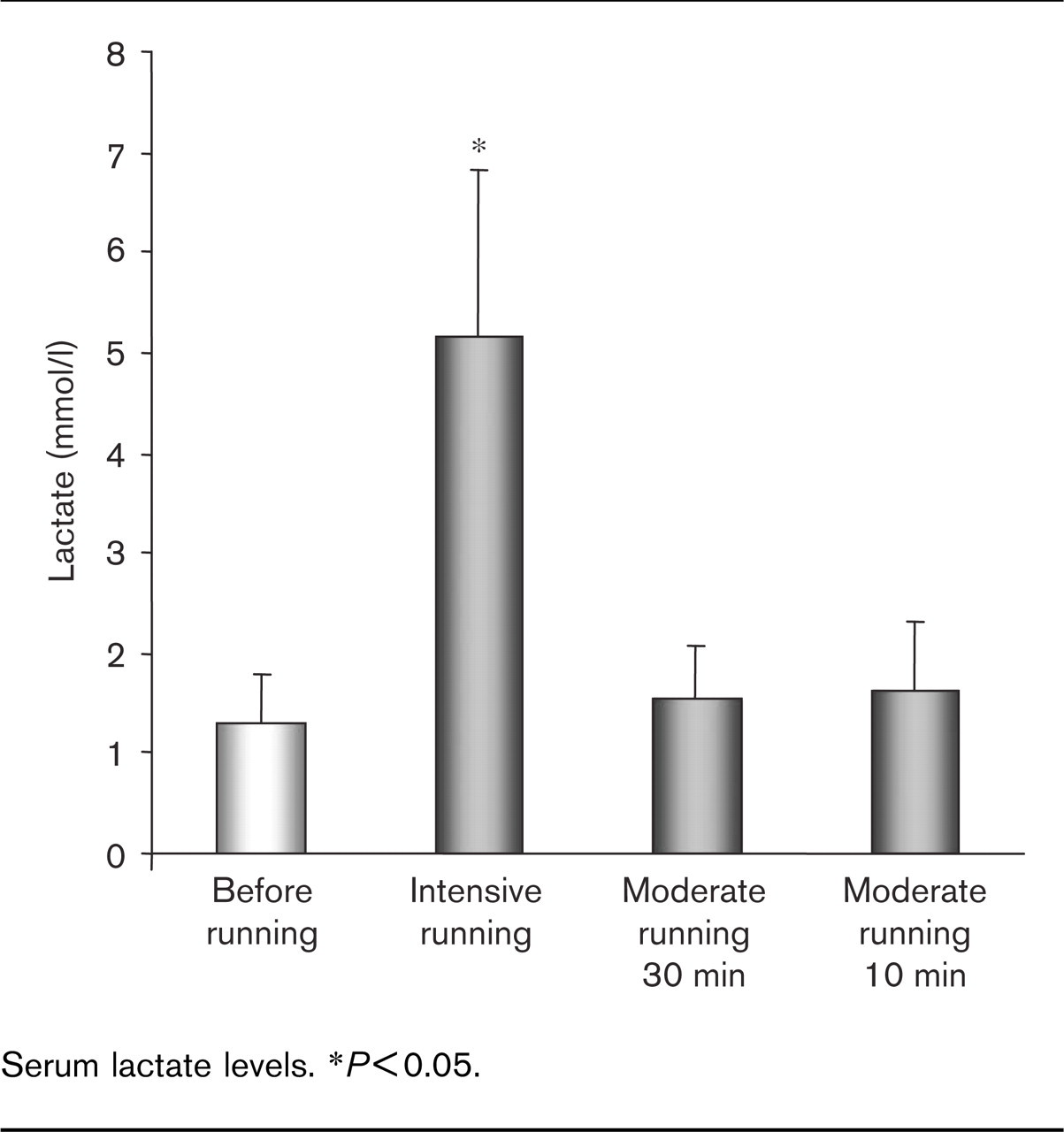
Serum lactate levels. ∗P<0.05.
Figure 2 depicts a representative FACS scan. Flow cytometry revealed significant upregulation of circulating CD34+/VEGFR2+, CD34+/CD133+ and CD34+/CD117+ populations 10 min after intensive running (Fig. 3). Similarly, 30 min of moderate running upregulated CD34+/VEGFR2+, CD34+/CD133+ and CD34+/CD117+ EPC (absolute numbers are listed in Figure 3 and Table 2). In order to assess the effects of exercise on EPC by an additional method, circulating EPC were isolated, cultured and quantified using DiLDL uptake and lectin staining [2]. Intensive as well as moderate running for 30 min upregulated circulating EPC (Fig. 3). Notably, the extent of EPC upregulation observed by flow cytometry or cell culture was not statistically different between both intensive and moderate exercise.
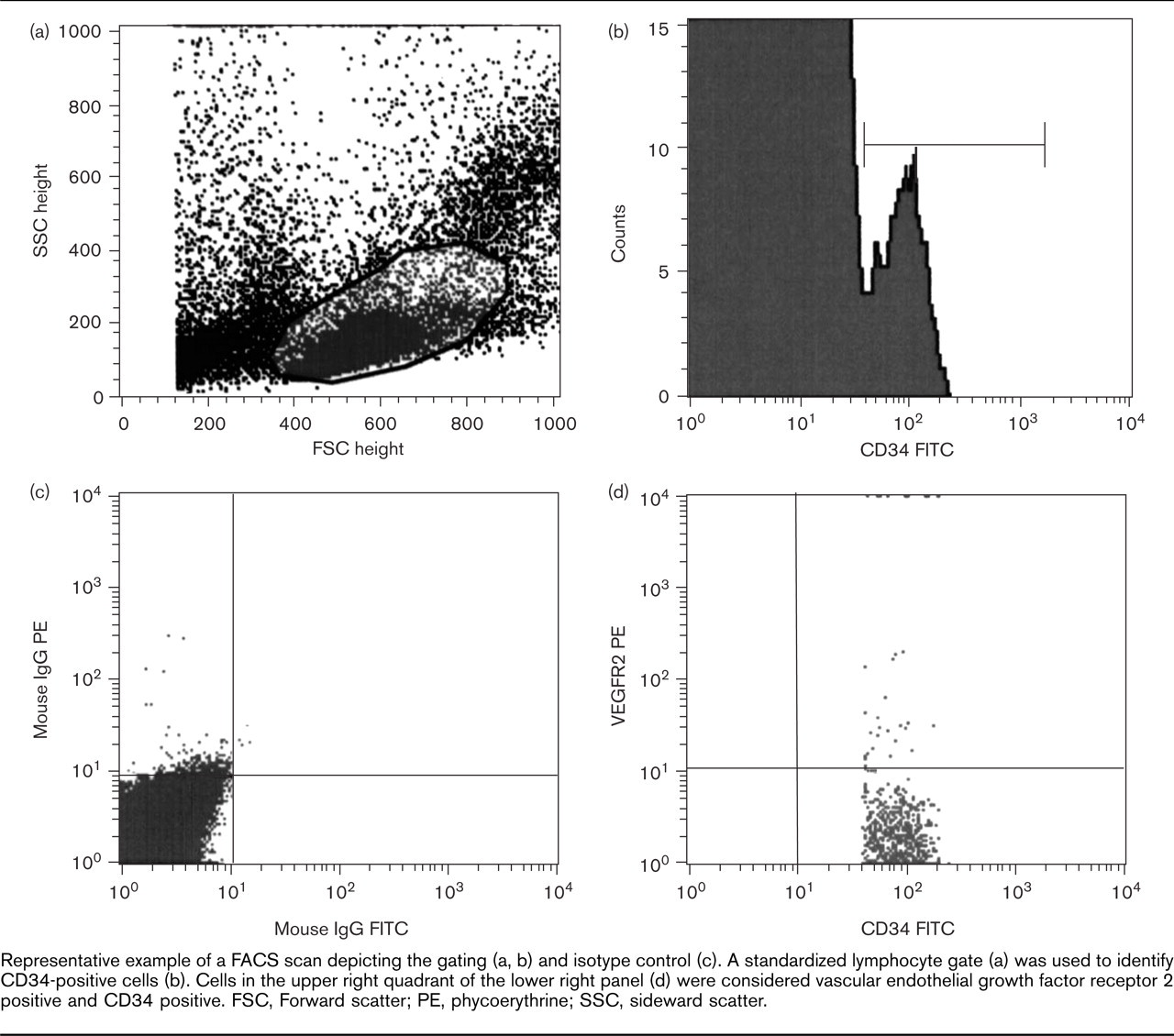
Representative example of a FACS scan depicting the gating (a, b) and isotype control (c). A standardized lymphocyte gate (a) was used to identify CD34-positive cells (b). Cells in the upper right quadrant of the lower right panel (d) were considered vascular endothelial growth factor receptor 2 positive and CD34 positive. FSC, Forward scatter; PE, phycoerythrine; SSC, sideward scatter.
Importantly, and in contrast to these findings, short-term moderate exercise for 10 min did not significantly upregulate any of the EPC parameters (Table 2).
Serum VEGF and cortisol have been associated with the upregulation of EPC and can be increased by physical exercise [27, 28]. In this healthy population, neither serum cortisol nor VEGF levels were significantly altered 10 min after intensive running (Fig. 4).
To characterize the time-dependent effects of running on EPC numbers and function, additional analyses were performed in a subgroup of six volunteers before, and 10 min, 30 min, 2 h, 6 h, and 24 h after intensive running. EPC numbers as determined by EPC culture (Fig. 5a) as well as flow cytometry (Fig. 5b, c) transiently increased with a maximum of 10–30 min after running.
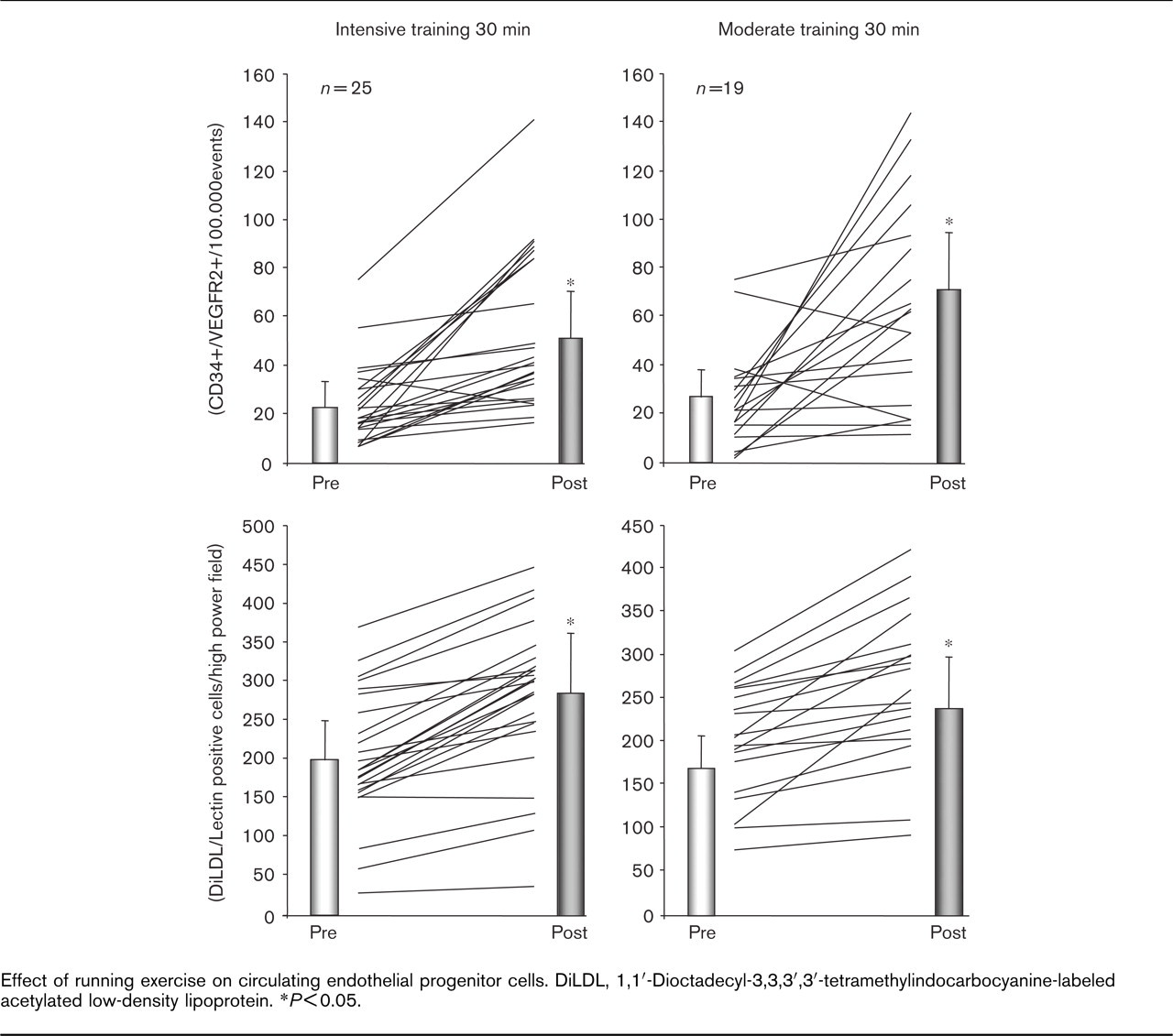
Effect of running exercise on circulating endothelial progenitor cells. DiLDL, 1,1′-Dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine-labeled acetylated low-density lipoprotein. ∗P<0.05.
Flow cytometry of CD34+/VEGFR2+, CD34+/CD133+ and CD34+/CD117+ endothelial progenitor cells
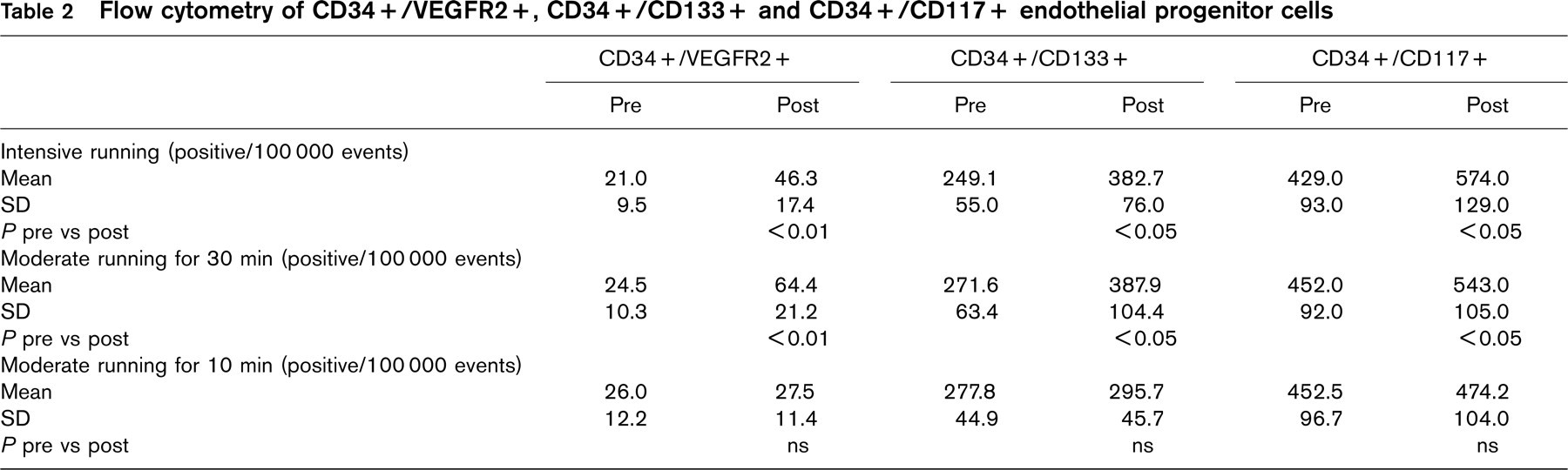
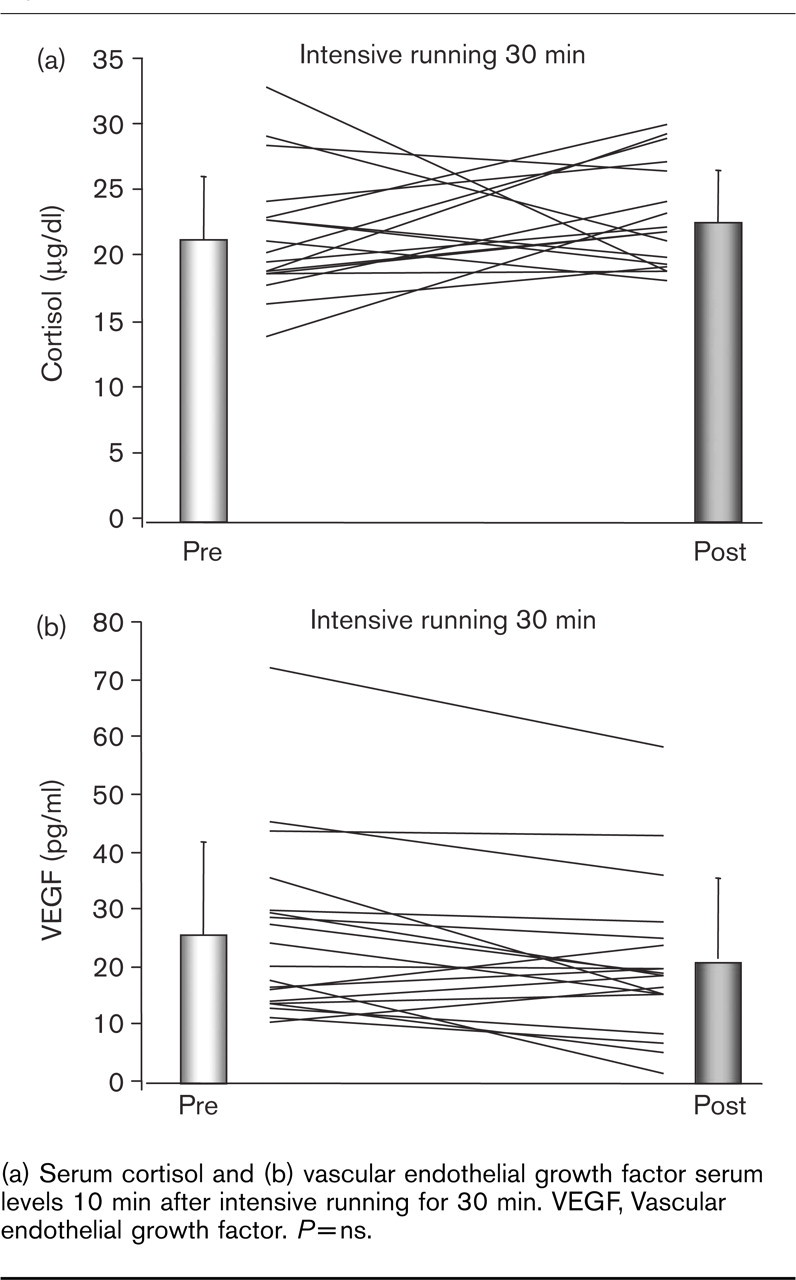
(a) Serum cortisol and (b) vascular endothelial growth factor serum levels 10 min after intensive running for 30 min. VEGF, Vascular endothelial growth factor. P=ns.
As EPC function may differ from EPC numbers, migratory capacity and colony-forming units were assessed in these volunteers. Modified Boyden chamber assays showed an increase in EPC migration in parallel to the time course of EPC numbers (Fig. 6a, b). Similarly, running upregulated the potency of EPC to generate colonies (Fig. 6c). Interestingly, this effect appeared to be more prolonged and remained significantly increased until 6 h after exercise.
Discussion
Recent evidence demonstrated the importance of EPC for vascular regeneration processes and their potential as a therapeutic as well as a preventative intervention [1, 3, 4, 6, 9, 10, 12, 29]. Here we identify running exercise as an intervention to increase EPC numbers in healthy men. The upregulation of EPC was observed using established FACS parameters as well as the quantification of cultured EPC.
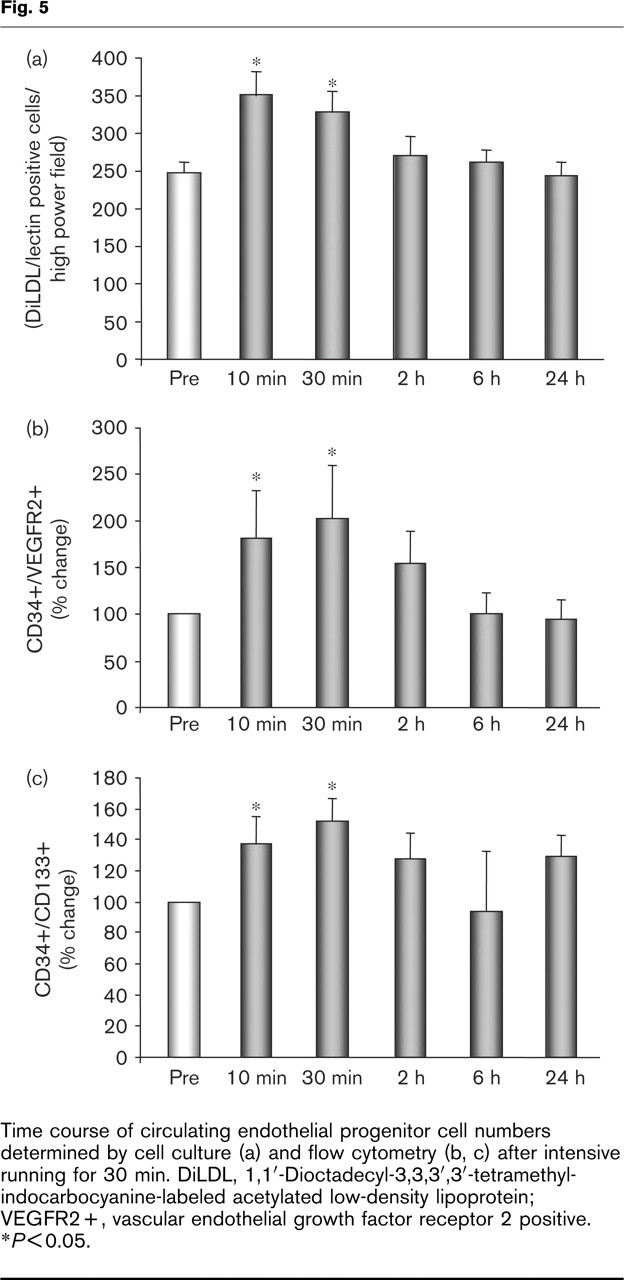
Time course of circulating endothelial progenitor cell numbers determined by cell culture (a) and flow cytometry (b, c) after intensive running for 30 min. DiLDL, 1,1′decyl-3,3,3′,3′-tetramethyl-indocarbocyanine-labeled acetylated low-density lipoprotein; VEGFR2+, vascular endothelial growth factor receptor 2 positive. ∗P<0.05.
To characterize this observation, the study compared three well-defined exercise intensities that were selected because of their relevance for primary and secondary prevention [20]. Running at 80% of the IAT (corresponding to 68% VO2max) for 30 min significantly increases circulating EPC numbers. Increasing the speed to 100% of IAT does not result in an additional increase in EPC. Moderate short-term running at 80% of IAT for 10 min is not sufficient to enhance EPC numbers. However, these data do not exclude the possibility of a functional improvement in EPC after short-term running. The findings correlate with the results from clinical recommendations, which indicated that at least a 20–30 min exercising period is needed to exert beneficial effects on surrogates such as body weight as well as on cardiovascular health [20]. Also, moderate rather than intensive sport programmes are recommended for the prevention of cardiovascular events [20].
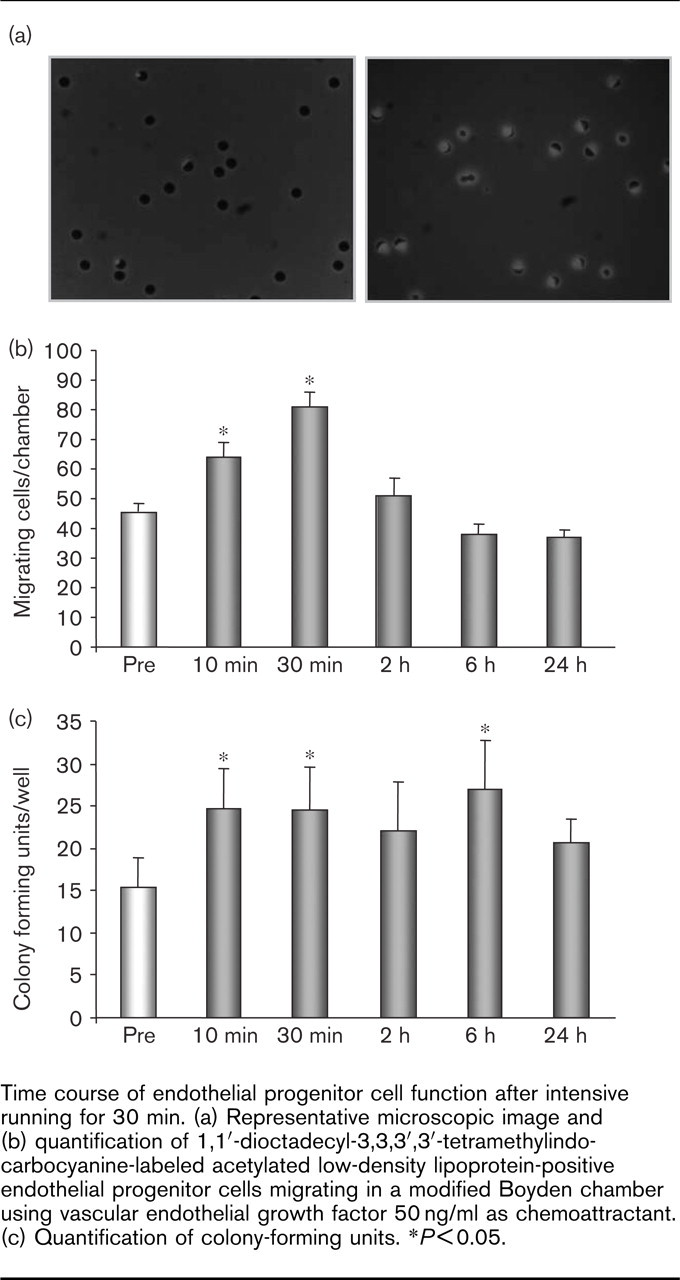
Time course of endothelial progenitor cell function after intensive running for 30 min. (a) Representative microscopic image and (b) quantification of 1,1′-dioctadecyl-3,3,3′,3′-tetramethylindo-carbocyanine-labeled acetylated low-density lipoprotein-positive endothelial progenitor cells migrating in a modified Boyden chamber using vascular endothelial growth factor 50 ng/ml as chemoattractant. (c) Quantification of colony-forming units. ∗P<0.05.
The selection of a very homogeneous study population of young and apparently healthy men mainly excludes the conceivable causes of EPC regulation other than exercise, such as ischaemia, inflammation or medication. This is supported by the lack of upregulation of serum cortisol or VEGF levels 10 min after intensive running. Cortisol represents an established marker of exercise stress and has been suggested potentially to mediate the mobilization of neutrophils from the bone marrow [27]. The volunteers were likely not to be totally relaxed because of a pre-starting condition. In addition, the duration of 30 min running was too short for the significant upregulation of cortisol levels. VEGF is well established as a mediator of ischaemia-induced EPC regulation [1, 11, 23, 28]. As intensive exercise did not alter VEGF serum concentrations, ischaemia appears unlikely to be the mechanism of the observed running-induced EPC upregulation in these healthy individuals. Therefore, the mechanism by which exercising increases the number of circulating EPC remains to be determined. It is reasonable to speculate that the well-established increase in nitric oxide bioavailability during exercising may be instrumental in an elevated release of EPC from the bone marrow [7, 16, 21]. In mice, the exercise-mediated upregulation of EPC was absent in endothelial nitric oxide synthase –/– animals or in the presence of a nitric oxide synthase inhibitor [21].
Little is known on the time course of EPC release after interventions that increase EPC numbers in humans. The kinetic study shows that exercise leads to a maximum increase between 30 min and 2 h. After 24 h a trend towards increased EPC remains that is no longer statistically significant. In contrast to patients with vascular risk, baseline EPC numbers are not impaired in healthy and physically active men [22, 23], therefore a more permanent increase in EPC after one exercise period is not likely. We speculate that repetitive exercise may result in a permanent increase in circulating EPC, especially in individuals with reduced baseline levels. Interestingly, the colony-forming units that reflect the potency of EPC to proliferate remain upregulated longer than circulating EPC numbers. This finding may suggest a period of improved endothelial regeneration and maintenance induced by exercise. Obviously, future studies need to determine whether regular exercising chronically increases EPC in the bloodstream and whether different modalities of activity exert differential effects on premature cells.
Prospective clinical trials quantitating the vasculoprotective effects of exercise in primary prevention are rare. Therefore, individual recommendations regarding the intensity and duration are difficult [14, 20]. For most vascular risk factors validated surrogate parameters are available, such as blood pressure for hypertension, or serum lipid and glucose levels for hypercholesterolemia and diabetes. However, few parameters are available to assess the efficacy of the vascular protection of an individual exercise training programme. It would be helpful to use a blood test to compare different training modalities and to advise patients regarding treatment goals. The study shows that the exercise-induced upregulation of EPC is observed after a 30-min period of running, whereas a moderate short-term 10-min exercise programme shows no impact on EPC numbers. In addition, not only intensive but also moderate running is sufficient to increase EPC in the peripheral blood. Therefore, the quantification of EPC may provide a novel surrogate parameter and tool to foster the study and the clinical use of exercise for the prevention of cardiovascular disease.
Despite extensive epidemiological data demonstrating a variety of beneficial effects of exercise, little is known about the underlying molecular mechanism [16–19, 30]. On the basis of our data we propose that the upregulation of EPC represents an important beneficial mechanism of physical training. In addition to the protective effects of EPC on the vasculature, recent data suggest that EPC may have the potential to transdifferentiate, for example into functionally active cardiomyocytes [10, 29]. Finally, it is interesting to speculate that the exercise-mediated regulation of adult stem cells may not be limited to endothelial progenitor cells, and may be important beyond cardiovascular medicine.
Footnotes
Acknowledgements
The authors would like to thank Simone Jäer and Isabel Páez-Maletz for excellent technical assistance.