
Abstract
Background Chronic heart failure (CHF) is accompanied by an inflammatory activation which occurs both systemically and in the skeletal muscle. Exercise training has been shown to reduce the local expression of cytokines and inducible nitric oxide synthase (iNOS) in muscle biopsies of CHF patients. INOS-derived NO can inhibit oxidative phosphorylation and contribute to skeletal muscle dysfunction in CHF.
Design To investigate the correlation between changes in local iNOS expression associated with regular exercise and changes in aerobic enzyme activities in the skeletal muscle of patients with CHF. Twenty male CHF patients [ejection fraction 25% (SE 2), age 54 (SE 2) years] were randomized to a training (n = 10) or a control group (C, n = 10).
Methods At baseline and after 6 months skeletal muscle iNOS expression was measured by real-time polymerase chain reaction. INOS protein and protein nitrosylation were assessed by immunohistochemistry. Cytochrome c oxidase (COX) activity was quantified electrochemically using the Clark oxygen electrode.
Results Exercise training led to a 27% increase in cytochrome c oxidase activity [from 21.8 (SE 3.2) to 27.7 (SE 3.5) nmol O2/mg per min, P=0.02 versus baseline]. Changes in iNOS expression and iNOS protein content were inversely correlated with changes in COX-activity (r= −0.60, P=0.01; r= −0.71, P<0.001).
Conclusions The inverse correlation between iNOS expression/iNOS protein content and COX-activity indicates that local anti-inflammatory effects may contribute to improved muscular oxidative metabolism.
Keywords
Introduction
The progression of chronic heart failure (CHF) depends not only on hemodynamic parameters but also on the process of systemic and local inflammatory activation with increased expression of tumor necrosis factor alpha (TNF-α), interleukin 1-beta (IL-1β) and interleukin 6 (IL-6) in the skeletal muscle of CHF patients [1–4]. Recently, we showed that a 6-month aerobic endurance training program attenuates skeletal muscle cytokine expression in patients with moderate CHF independent from serum cytokine levels [4]. This finding may have several important consequences for skeletal muscle mass, function and metabolism.
Cytokines act as catabolic factors involved in peripheral muscle wasting and cardiac cachexia [1, 2]. Increased levels of serum TNF-α for example were found in patients with reduced leg cross-sectional area and peripheral muscle strength [5]. Therefore, a reduction in local cytokine expression may also attenuate skeletal muscle wasting in CHF [6].
Cytokines increase local generation of reactive oxygen species (ROS) which further potentiate the cytotoxic and proapoptotic effects of the inflammatory process [7–10].
Cytokines induce the muscular expression of inducible nitric oxide synthase (iNOS) via NFκB-dependent mechanisms in CHF [11, 12]. The intracellular concentrations of iNOS-derived NO may reach cytotoxic levels and inhibit the mitochondrial aerobic energy generation and transfer [12,13]. In addition, the presence of elevated intracellular ROS concentrations accelerates the reaction of NO and O− 2 to peroxynitrite (ONOO−), which also acts as a potent inhibitor of the aerobic metabolism [14–16].
The aim of this prospective randomized clinical trial was therefore to assess the potential correlation between changes in local inflammatory activation, iNOS expression, and inhibition of the aerobic energy generation [as measured by cytochrome c oxidase (COX) activity] associated with a 6-month exercise training program in patients with stable CHF.
Methods
Patient population and baseline characteristics
The study group included 20 male patients ≤70 years with chronic heart failure (New York Heart Association [NYHA] – functional class II–III) as a result of dilated cardiomyopathy or ischemic heart disease as assessed by cardiac catheterization. All patients had clinical, radiological and echocardiographic signs of chronic heart failure and a left ventricular ejection fraction (LVEF) of < 40% as assessed by ventriculography. They received their individually tailored medication including angiotensin-converting enzyme inhibitors, digitalis, diuretics, and β-blockers when tolerated. They had to be in a clinically stable condition for a minimum of 3 months prior to enrolment. No patient had any clinical evidence of peripheral arterial occlusive disease.
The study protocol was approved by the Ethics Committee of the University of Leipzig, and written informed consent was obtained from all subjects. The investigation conforms with the principles outlined in the Declaration of Helsinki.
Twenty patients with stable CHF were randomly assigned to the exercise training group (10 patients) or to the control group (10 patients). As previously reported [4] patients in the training group and in the control group showed a significantly reduced left ventricular ejection fraction [training group: 26.1 (SE 3.1)%, control group: 24.7 (SE 2.4)%, P = NS] and exercise capacity as determined by peak oxygen uptake [training group: 20.3 (SE 1.0) ml/kg per min, control group: 17.9 (SE 1.6) ml/kg per min, P = NS] (Table 1).
Baseline characteristics
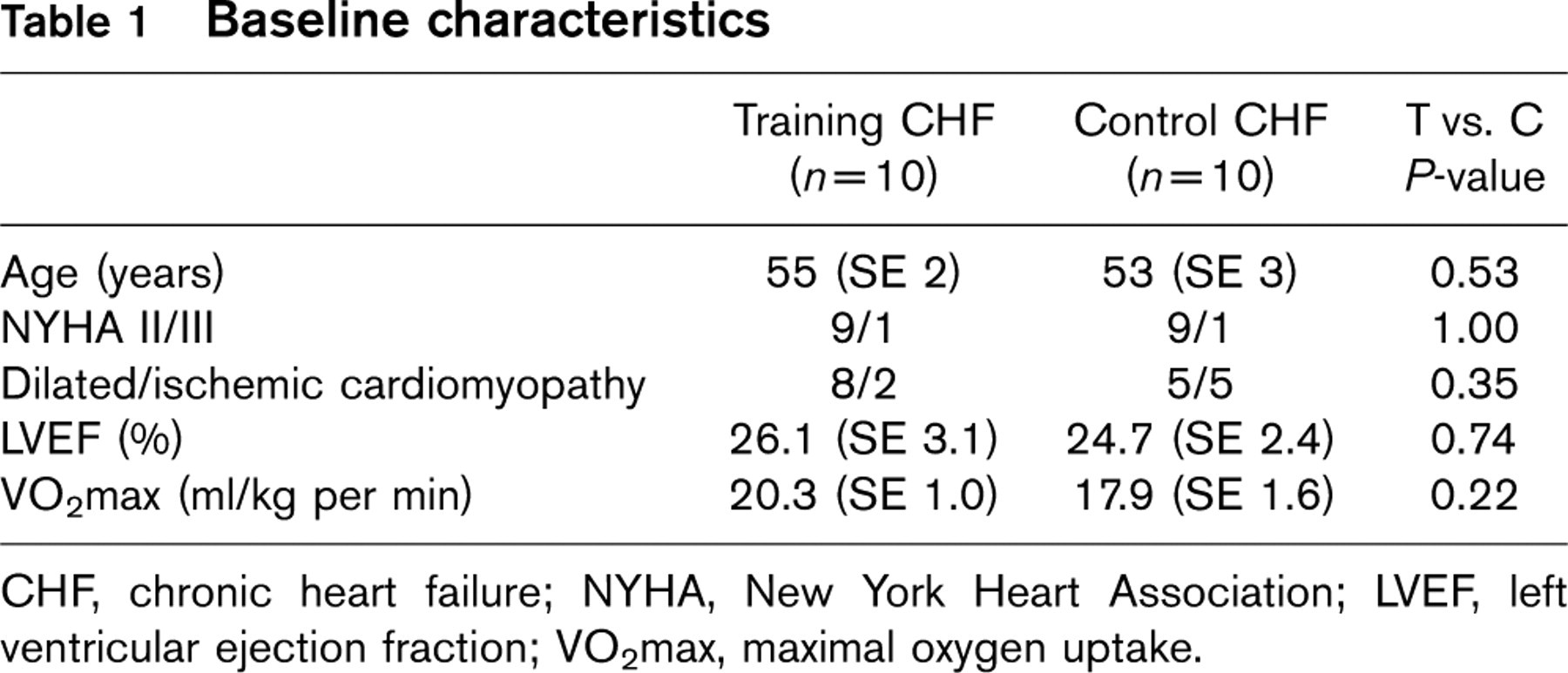
CHF, chronic heart failure; NYHA, New York Heart Association; LVEF, left ventricular ejection fraction; VO2max, maximal oxygen uptake.
Correlation of changes in local inflammatory parameters with changes in cytochrome c oxidase activity
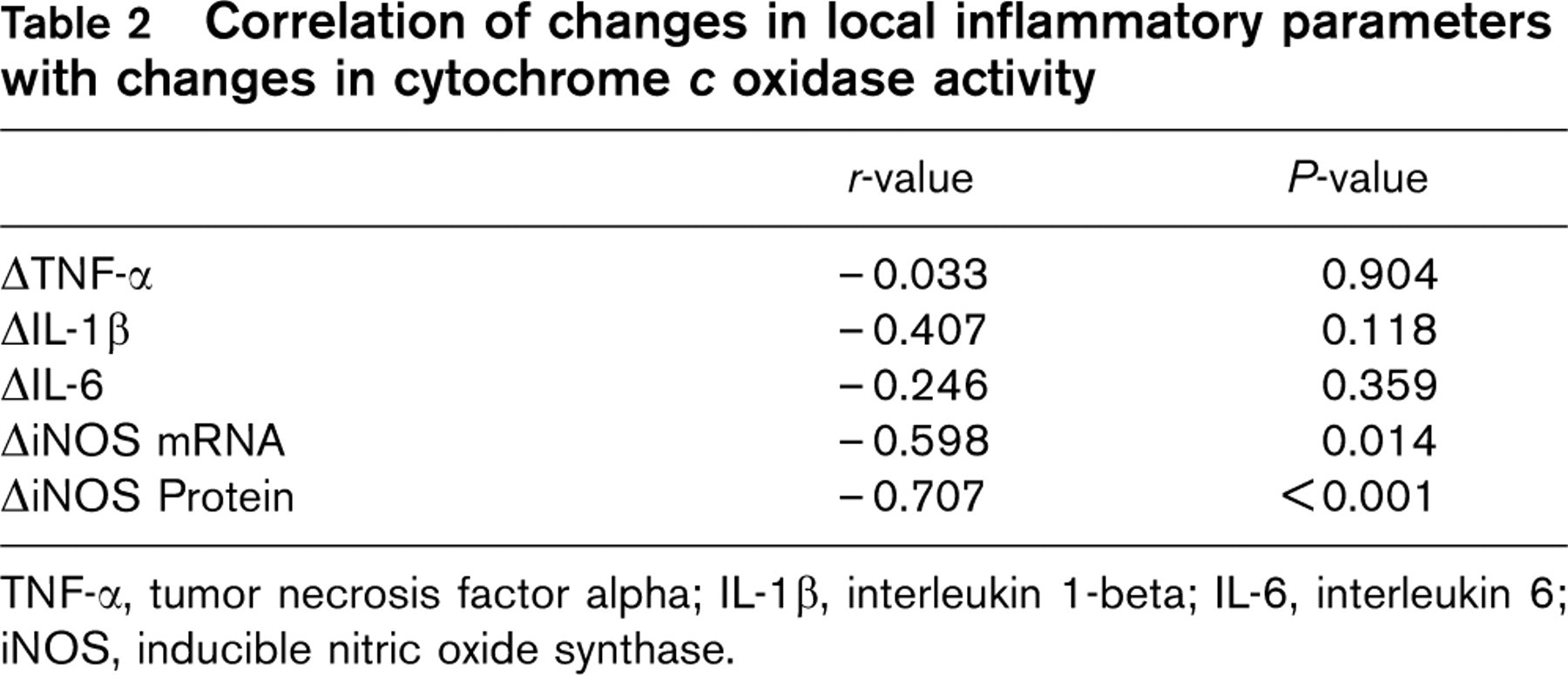
TNF-α, tumor necrosis factor alpha; IL-1β, interleukin 1-beta; IL-6, interleukin 6; iNOS, inducible nitric oxide synthase.
Medical therapy was similar in the training and the control group. Patients were on angiotensin-converting-enzyme inhibitors (10/10 in the training group and 9/10 in the control group), digitalis (5/10 and 5/10, respectively), diuretics (4/10 and 9/10, respectively), and β-blockers (4/10 and 4/10, respectively). The medication did not change between 4 weeks prior to enrolment and the end of the study period after 6 months (Table 2).
Training intervention
In order to ensure close supervision, the initial phase of the exercise program was performed on an in-patient basis. During the first 2 weeks patients exercised four to six times daily for 10 min on a bicycle ergometer. Workloads were adjusted so that 70% of the symptom-limited maximal oxygen uptake was reached. Before discharge from the hospital, maximal symptom-limited ergospirometry was performed to calculate training target heart rate for home exercise training, which was defined as the heart rate reached at 70% of the maximal oxygen uptake during symptom-limited exercise. Upon discharge patients were provided with bicycle ergometers for daily home exercise training. They were asked to exercise close to their target heart rate daily for 20 min per day. In addition, they were expected to participate in at least one group training session of 60 min each week. Exercise sessions consisted of walking, callisthenics, and non-competitive ball games.
Patients assigned to the control group continued their individually tailored cardiac medication and were supervised by their private physicians. All examinations including exercise testing were repeated after 6 months.
Exercise testing and respiratory variables
Exercise testing was performed on a calibrated, electronically braked bicycle in an upright position with workload increasing progressively every 3 min in steps of 25 W beginning at 25 W. Respiratory gas exchange data were determined continuously throughout the exercise test as previously described [17].
Skeletal muscle biopsy
Percutaneous needle biopsies were obtained at least 2 days before exercise testing from the middle part of the vastus lateralis muscle under local anesthesia as described previously [18]. The biopsies were either fixed with 4% buffered formaldehyde or snap frozen in liquid nitrogen and stored at −80°C.
Quantification of local iNOS expression by quantitative reverse transcriptase-polymerase chain reaction
To quantify local iNOS mRNA expression real time polymerase chain reaction (PCR) (Light Cycler System, Roche Diagnostics Inc., Mannheim, Germany) was used. Briefly, aliquots (1 μg) of isolated total RNA were reverse transcribed in a volume of 25 μl into cDNA using random hexamers. cDNA (1 μl) was added to the PCR reaction containing the following specific primers: iNOS-U: 5′-CAA GAA CGT GTT CGC CAT GAG GC-3′, iNOS-L: 5′-GGT GAG GGC TTG CCT GAG TGA-3′.
The results for mRNA concentrations are expressed as a ratio over 18SrRNA, which was amplified as house keeping gene using the following primers 18S rRNA-U: 5′-TAG AGG GAC AAG TGG CGTTC-3′ and 18S rRNA-L: 5′-TGT ACA AAG GGC AGG GAC TT-3′. The variability for triplicate measurements was less than 5%.
Immunohistochemistry of iNOS and nitrotyrosine
To specifically detect the inducible isoform of nitric oxide synthase in tissue sections a polyclonal anti-iNOS specific antibody (Transduction Laboratories, Lexington, USA) was used and the sensitivity was enhanced by an immunohistochemical protocol using biotinylated tyramide (NEN-DuPont, Germany) as described previously [18]. For consecutive duplicate biopsy samples, the variation regarding the percentage of iNOS-positive tissue area has been shown to be in the order of less than 10%.
Nitrotyrosine production in the skeletal muscle biopsies was assessed in order to confirm that nitric oxide and peroxynitrite, respectively, were present before fixation. To specifically detect the nitration of the ortho-position of tyrosine residues on paraffin sections the method described by Bachmaier et al. [19] was applied.
Measurement of cytochrome c oxidase activity
In muscle homogenates the activity of cytochrome c oxidase (EC 1.9.3.1) was measured polarographically at room temperature using a Clark oxygen electrode as described by Sottocasa et al. [20]. In brief, an aliquot of the homogenate was added to 1.6 ml of Hepes–Sucrose buffer (250 mmol/l sucrose, 10 mmol/l Hepes, 20 mmol/l taurine) containing 2 mmol/l ascorbate, 25 μg/ml antimycin A and 0.3 mmol/l N,N,N′,N′;-tetramethyl-p-phenyle-nediamine (TMPD). The oxygen consumption was measured continuously and the reaction was initiated by the addition of 10 μmol/l cytochrome c. The activity of cytochrome c oxidase was calculated assuming an oxygen concentration of 240 nmol O2/ml and expressed as nmol O2 consumption/min per mg total protein.
Measurement of cytochrome c oxidase protein content
Cytochrome c oxidase protein expression was measured by Western blot using a polyclonal anti-COX antibody (MobiTec, Göttingen, Germany). Briefly, 20 μg skeletal muscle proteins were separated on a 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), followed by electrotransfer to a polyvinylidene fluoride membrane (PVDF; Serva Biochemica, Heidelberg, Germany) according to the method described by Towbin et al. [21]. The transferred proteins were incubated with the specific polyclonal anti-COX antibody. The bound antibody was detected by a peroxidase-coupled anti-rabbit antibody followed by a color reaction using diaminobenzidine (DAB) as substrate for peroxidase. To quantify COX protein the stained blot was analyzed by densitometry with a one-dimensional-analysis software package (One-Dscan; Scanalytics, Billerica, Massachusetts, USA). To compensate for blot-to-blot variations in staining intensity an internal standard was loaded on each blot, and the densitometry results were expressed as relation between sample intensity/internal standard intensity. The variability for duplicate measurements was less than 10%.
Statistical analysis
All data are expressed as mean value ± standard error of mean (SE). The data were tested for normal distribution using the Kolmogorov–Smirnov test, and for homogeneity of variances using Levene's test. Baseline data in healthy subjects, control patients, and training patients were compared using the one-way analysis of variance (ANOVA) procedure. The prevalence of dilated and ischemic cardiomyopathy in each group was compared using Fisher's exact test. Among the training and control group intragroup comparisons were made using the Wilcoxon test and intergroup comparisons were assessed using the Mann–Whitney-U test. Possible correlations between variables were analyzed using linear regression analysis. A P-value of less than 0.05 was considered statistically significant.
Results
Clinical follow-up
Exercise training improved peak oxygen uptake by 29% in the training group {from 20.3 [95% confidence interval (CI) 18.1–22.4] to 26.1 (95% CI 22.7–29.5) ml/kg per min, P < 0.001 versus control group} whereas it remained virtually unchanged in the control group [17.9 (95% CI 14.4–21.5) versus 18.1 (95% CI 15.7–20.5) ml/kg per min, P = NS]. No deaths occurred during the study period. One patient in the training group was admitted to hospital due to symptomatic bradyarrhythmia requiring pacemaker implantation. He continued the study after discharge from hospital.
Skeletal muscle iNOS expression and nitrotyrosine concentration
Six months of aerobic exercise training reduced local iNOS expression by 52% [from 6.3 (SE 1.2) to 3.0 (SE 1.0) relative units, P = 0.017 versus baseline, P = 0.007 versus control group] and iNOS protein content by 35% [from 2.3 (SE 0.5) to 1.5 (SE 0.4)% positive tissue area, P = 0.017 versus baseline, P = 0.007 versus control group for change]. The reduction in local iNOS expression after the training intervention was associated with a 38% decrease in skeletal muscle nitrotyrosine content [from 4.7 (SE 0.8) to 3.1 (SE 0.5)% positive tissue area, P = 0.028 versus baseline, P = 0.016 versus control group for change, compare [4]].
Cytochrome c oxidase activity
Training led to a 27% increase in cytochrome c oxidase activity [from 21.8 (SE 3.2) to 27.7 (SE 3.5) nmol O2/mg per min, P = 0.02 versus baseline, P < 0.01 versus control group]. In the control group a trend towards further decline in cytochrome c oxidase activity was observed [from 19.6 (SE 2.8) to 16.3 (SE 2.0) nmol O2/mg per min, P = 0.15 versus baseline] (Fig. 1).
Of note, COX protein did not change significantly in the training or the control group [training group: 0.081 (SE 0.013) versus 0.089 (SE 0.012) arbitrary units; control group: 0.078 (SE 0.013) versus 0.069 (SE 0.019) arbitrary units, P = NS].
Correlation between ΔiNOS expression and ΔCOX activity
The change in cytochrome c oxidase activity correlated with the change in peak oxygen uptake (r = 0.76, P < 0.001). The change in local iNOS expression, on the other hand, was significantly correlated with the change in COX activity (r = −0.60, P = 0.01) (Fig. 2). The linear correlation between iNOS and COX activity was even closer for iNOS protein as quantified by immunohistochemistry (r = −0.71, P < 0.001).
Discussion
It has been consistently documented that carefully designed exercise training programs in CHF have the potential to improve exercise tolerance, increase peak oxygen uptake, and augment skeletal muscle cytochrome c activity and mitochondrial volume density [22]. Until now it was unclear, however, if exercise training also affects the inflammatory process and prevents iNOS expression in CHF. We have previously shown that exercise training has potent anti-inflammatory effects in the skeletal muscle of patients with stable CHF [4]. In the present consecutive study we extend the implications of local cytokine and iNOS reduction to the metabolic and functional consequences. The decrease in local iNOS expression and peroxynitrite generation associated with training is paralleled by a significant increase in cytochrome c oxidase activity.
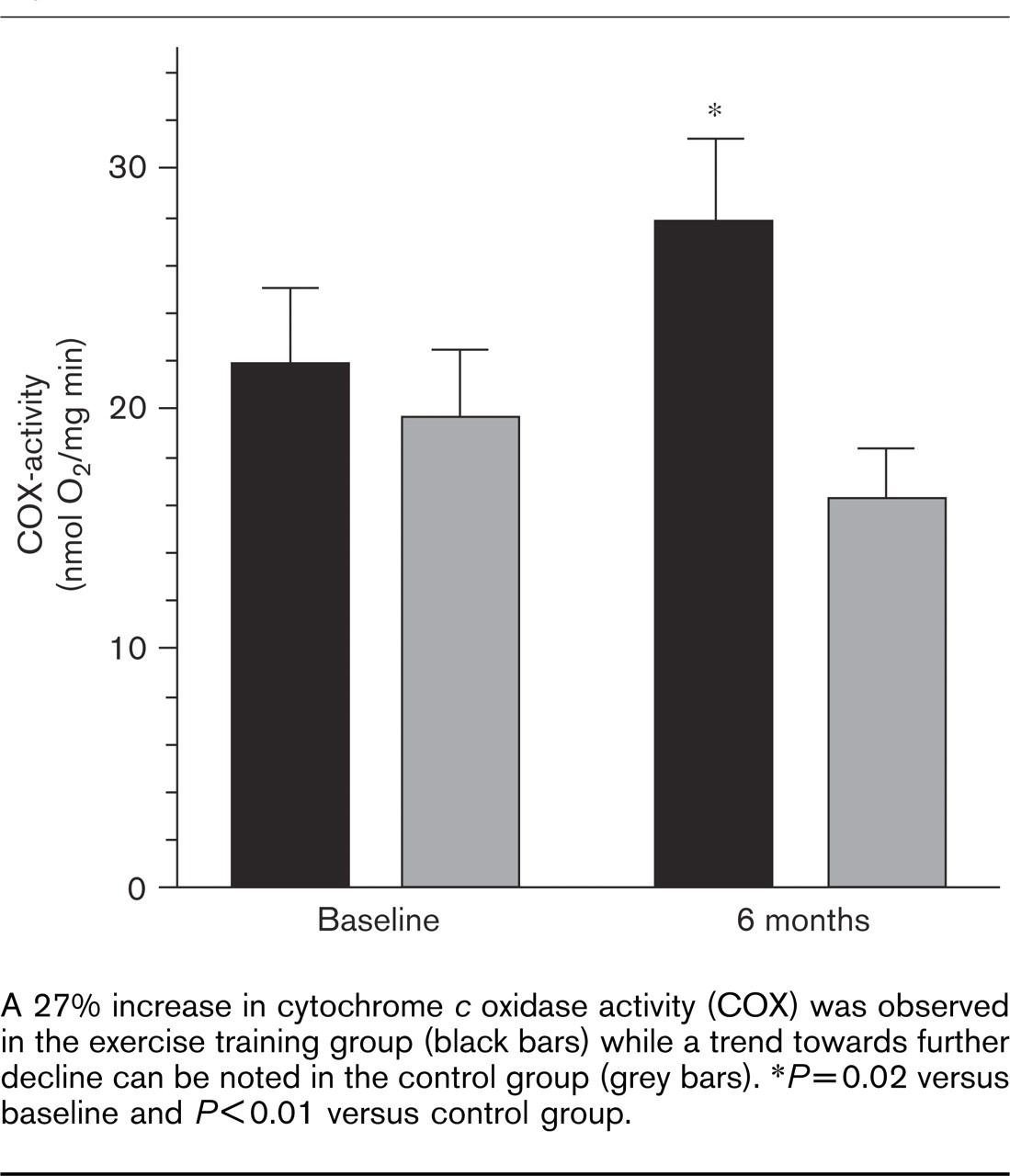
A 27% increase in cytochrome c oxidase activity (COX) was observed in the exercise training group (black bars) while a trend towards further decline can be noted in the control group (grey bars). ∗P=0.02 versus baseline and P<0.01 versus control group.
Close linear relations were confirmed between changes in local iNOS expression/protein content and changes in COX activity. Changes in COX activity, on the other hand, were significantly correlated to changes in peak oxygen uptake. These findings imply that the reduced NO/ONOO− production may contribute to the improved aerobic metabolism by disinhibition of key enzymes of the oxidative phosphorylation. However, VO2max as a global physiologic index of exercise tolerance integrates organ functions beyond the skeletal muscle metabolism (i.e. cardiac output, vascular function, and pulmonary function). The observed correlation between changes in COX activity and peak oxygen uptake does therefore not exclude that changes in these physiologic determinants may also have contributed to the overall benefit of the training intervention.
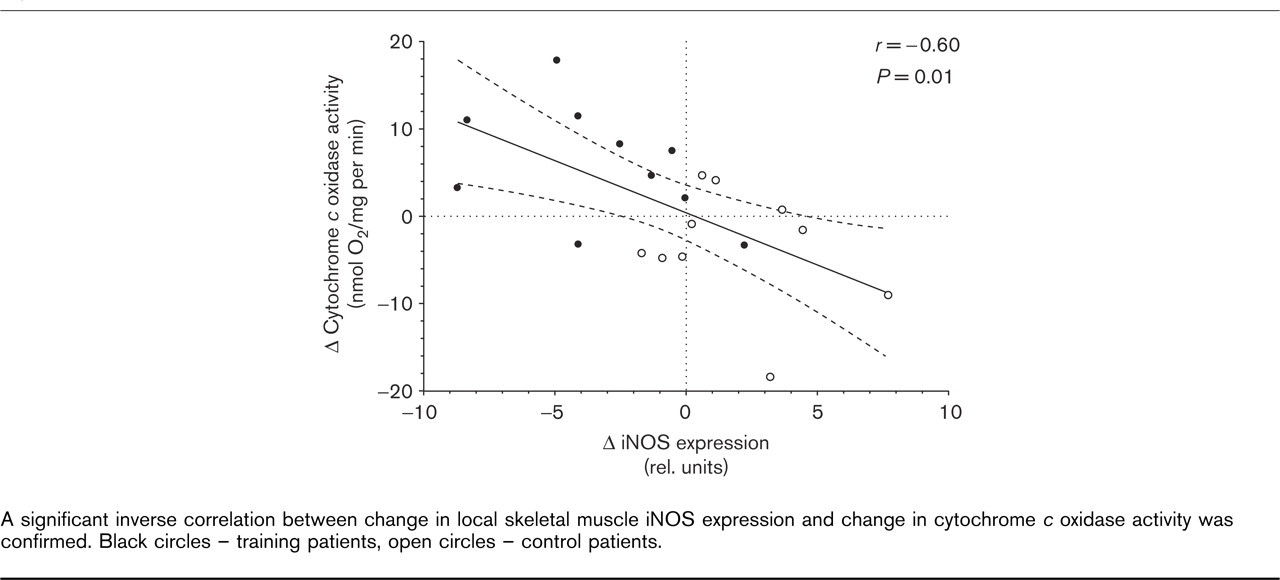
A significant inverse correlation between change in local skeletal muscle iNOS expression and change in cytochrome c oxidase activity was confirmed. Black circles – training patients, open circles – control patients.
Effects of exercise on iNOS-induction in the skeletal muscle of patients with CHF
Since the first description of iNOS expression in skeletal muscle biopsies from patients with CHF [12, 18, 23] it has been a matter of discussion whether to view it as a bystander of the inflammatory activation or as an important part of it. Recently, we were able to document that iNOS can produce intracellular NO concentrations high enough to interfere with key aerobic enzymes (i.e. cytochrome c oxidase) and with the transfer of high-energy phosphates from the mitochondrium into the cytosol (i.e. mitochondrial creatine kinase) [18].
The hypothesis that NO acts as a physiologic inhibitor of oxidative phosphorylation, for example by reversibly competing with oxygen for the reduced oxygen binding site of cytochrome oxidase, has been confirmed both in vitro and in vivo [24]. The reduced iNOS expression in the skeletal muscle after 6 months of exercise training may therefore lead to a disinhibition of the aerobic metabolism and may contribute to the improved peak oxygen uptake after the training intervention.
Basically, iNOS may be induced by three known pathogenetic factors: Cytokines, reactive oxygen species (ROS), and mechanical factors like increased wall-stress in the myocardium [25]. These factors converge on the activation of NF-κB which is necessary to initiate transcription of iNOS mRNA. Recently, we were able to confirm a correlation between elevated NF-κB levels in skeletal muscle biopsies from patients with severe CHF and increased iNOS expression [26].
In addition to cytokines, oxidative stress may play an important role in amplifying iNOS expression: In hepatocytes it was observed that the iNOS promotor region contains an antioxidant responsive element (ARE) by which ROS enhance iNOS transcription [27]. The antioxidative effects of training in the skeletal muscle described by Ennezat et al. [28] may therefore contribute to the reduction of iNOS expression.
Role of NO for the regulation of the mitochondrial aerobic energy metabolism
NO exerts a biphasic effect on mitochondrial respiration. At low concentrations for short periods NO specifically and reversibly inhibits cytochrome c oxidase. At higher levels, and more prolonged exposure, NO, or its more reactive oxide ONOO−, may cause irreversible inhibition of respiration and structural damage to the mitochondria [29].
While the physiological sources of NO, the endothelial NOS (ecNOS), and the neuronal NOS (nNOS), are acutely and transiently activated by calcium/calmodulin or by phosphorylation, iNOS remains constitutively active once it is induced by cytokines (like IL-1β, TNF-α, lipopolysaccharides, or oxidative stress. As a consequence intracellular NO levels may reach toxic levels [18].
The reversible inhibition of cytochrome c oxidase (i.e. complex IV) is important in both physiological and pathological contexts: COX is responsible for about 90% of the oxygen consumption in mammals. NO competes with molecular oxygen for the active center of the enzyme starting at concentrations 1–200 nmol/l [30]. It can therefore be presumed that the elevated concentrations of NO and peroxynitrite present in the skeletal muscle of CHF patients may indeed interfere with normal mitochondrial respiration (Fig. 3).
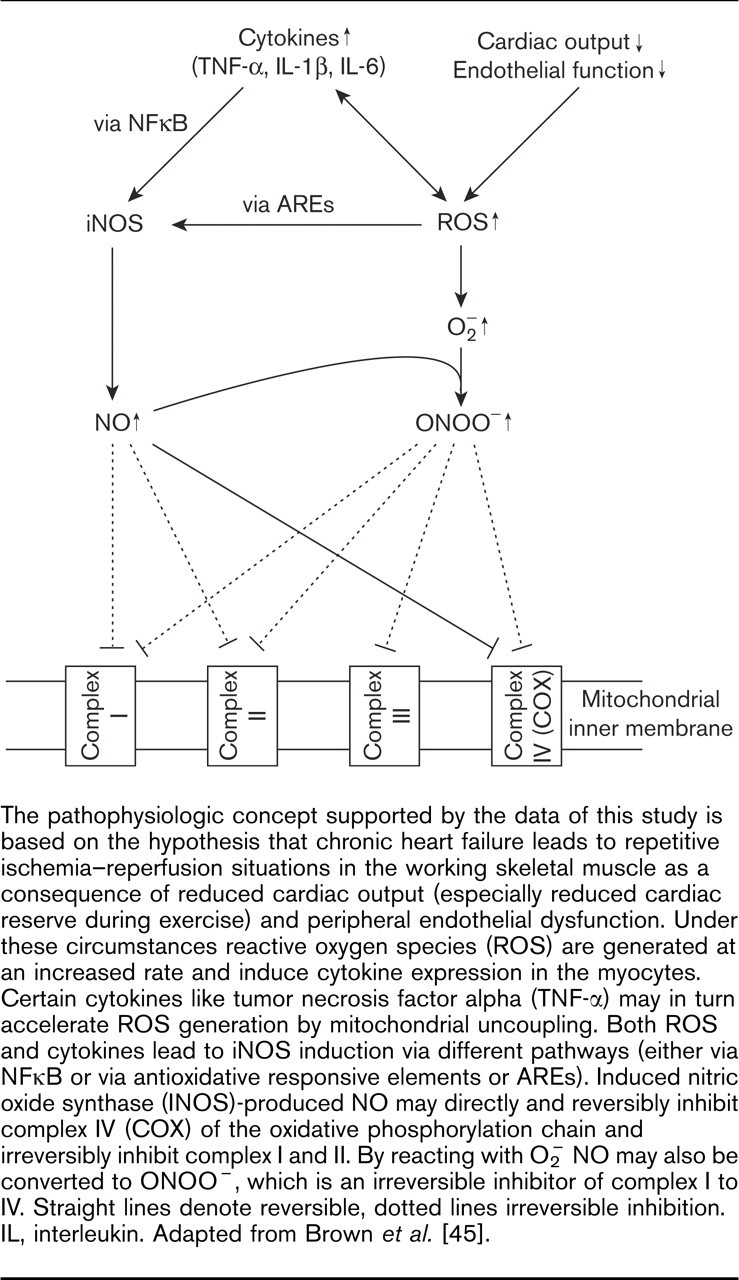
The pathophysiologic concept supported by the data of this study is based on the hypothesis that chronic heart failure leads to repetitive ischemia–reperfusion situations in the working skeletal muscle as a consequence of reduced cardiac output (especially reduced cardiac reserve during exercise) and peripheral endothelial dysfunction. Under these circumstances reactive oxygen species (ROS) are generated at an increased rate and induce cytokine expression in the myocytes. Certain cytokines like tumor necrosis factor alpha (TNF-α) may in turn accelerate ROS generation by mitochondrial uncoupling. Both ROS and cytokines lead to iNOS induction via different pathways (either via NFκB or via antioxidative responsive elements or AREs). Induced nitric oxide synthase (INOS)-produced NO may directly and reversibly inhibit complex IV (COX) of the oxidative phosphorylation chain and irreversibly inhibit complex I and II. By reacting with O− 2 NO may also be converted to ONOO−, which is an irreversible inhibitor of complex I to IV. Straight lines denote reversible, dotted lines irreversible inhibition. IL, interleukin. Adapted from Brown et al. [45].
NO may, however, irreversibly inhibit the mitochondrial aconitase, complex I and complex II by displacing iron from the sulphur-iron centers of these enzymes [31–33]. The range of mitochondrial components affected by ONOO− is even wider. The complexes I–IV, the ATP synthase, aconitase, and creatine kinase are inhibited, the mitochondrial genome and membrane are damaged leading to mitochondrial swelling, uncoupling, depolarization, and permeability transition [34]. The release of COX from the mitochondria into the cytosol is a potent pro-apoptotic stimulus [35].
In the present study we present conclusive evidence that a long-term training intervention is associated with reduced intracellular NO generation as indicated by lower iNOS mRNA expression, iNOS protein content, and attenuated protein nitrosylation. The lower NO production was correlated with enhanced skeletal muscle COX activity. Although it cannot be ruled out that COX activity was also increased secondary to the increased energy demand with training, it is consistent with our previous findings of a close correlation between iNOS expression and reduction of peak oxygen uptake [18] that lower local NO levels may significantly contribute to the higher oxidative capacity.
Two major factors are involved in the disinhibition of COX activity after training: (1) a reduction of local ROS generation [36]; and (2) a significant decrease in local cytokine expression and inflammatory activation with NFκB-mediated iNOS expression [4].
Both training-associated changes are closely interwoven. On the one hand, proinflammatory cytokines like TNF-α can increase iNOS expression [37]. On the other hand, increases in local ROS-generation can induce iNOS by interacting with an antioxidant responsive element in the iNOS gene [27].
While there is consensus about the inhibitory effects of NO and its oxides on mitochondrial respiration [24, 38, 39] two aspects of NO and oxidative phosphorylation remain controversial: (1) the role of endothelium-derived NO for the regulation of cellular respiration; and (2) the pathophysiological impact of the inhibition of cellular respiration for disease progression. However, both issues have been discussed primarily for the myocardium.
Several authors described an increase in oxygen consumption after inhibition of endothelial NO production both in the heart [40] and the peripheral skeletal muscle [41]. However, the inhibitors used for inhibition of NO generation in these experiments (i.e. L-NMMA and derivatives) are not specific for the endothelial isoform of nitric oxide synthase (ecNOS) and can also inhibit intracellular isoforms like nNOS and iNOS. Additionally, CHF is characterized by a reduced basal endothelial NO production [42], which would reduce NO-associated inhibition of mitochondrial respiration in the skeletal muscle of CHF patients.
Based on in vitro experiments it has been proposed that the inhibitory effects of NO on mitochondrial respiration may also have beneficial effects on myocardial function in heart failure [43]. However, no such studies have been produced for the peripheral skeletal muscle. On the contrary, peak oxygen consumption, which provides an estimate of total peripheral skeletal muscle oxidative capacity, has been viewed as an important predictor associated with improved survival in CHF. In the context of other studies indicating improved survival by exercise training in CHF [44] we believe that the reduction of iNOS expression and improved COX activity indicate a favorable effect of training on inflammatory activation and a partial reversal of disease-associated skeletal muscle metabolic impairment.
Conclusion
In conclusion, a 6-month exercise training program in patients with stable CHF is effective in reducing iNOS expression and intracellular accumulation of NO. The reduction in local iNOS expression/protein content is closely related to improved COX activity and enhanced peak oxygen uptake. In this sense the present study extends the beneficial anti-inflammatory effects of training into the metabolic functioning of the skeletal muscle and adds to the pathophysiological framework to explain the improved aerobic capacity after training interventions in CHF.