
Abstract
Background
Myocardial oxidative stress is believed to play an important role in the pathogenesis of alcoholic cardiomyopathy. Strenuous physical exercise has been shown to increase or decrease myocardial oxidative stress depending on the mode and duration of the exercise intervention. Given the possibility of individuals to engage in both alcohol consumption and weight-training exercise, we have examined the effect of resistance exercise training and chronic alcohol consumption on myocardial oxidative stress in rats.
Methods
Forty Sprague-Dawley rats were randomly assigned to one of four experimental groups: sedentary, sedentary plus alcohol treatment, resistance training, or resistance training plus alcohol treatment. Rats in the alcohol groups received a liquid diet containing alcohol (35% of kilocalorie intake) for 6 weeks. Non-alcohol groups were pair-fed the same liquid diet supplemented with a maltose dextrin caloric substitute. Rats in the resistance training groups were trained to rise onto their hind limbs while wearing lead-weighted vests 30 times per training session, 3 days per week during the 6 week experimental period.
Results
Alcohol treatment in the sedentary animals resulted in greater levels of cardiac malondialdehyde, a marker of lipid peroxidation, and a depressed index of myocardial antioxidant potential compared with all other groups (P≤0.05). Hearts from the resistance training plus alcohol animals exhibited malondialdehyde and antioxidant levels similar to sedentary controls, suggesting that resistance training protected against the alcohol-induced myocardial stress.
Conclusion
These results suggest that resistance training may attenuate the damaging effects of alcohol on the heart and preserve myocardial antioxídant capacity.
Introduction
Cardiac muscle relies heavily on oxidative metabolism at rest and during exercise. Most of the oxygen consumed by the myocardium serves as the final electron acceptor in the mitochondrial respiratory chain where it is reduced to water. Some of the oxygen, however, will undergo a series of single electron reductions that result in the formation of reactive oxygen species (ROS) [1]. Excess production of ROS can lead to cellular injury such as membrane lipid peroxidation and protein oxidation if they are not scavenged by the myocardial antioxidant system [2]. ROS-induced myocardial injury is thought to play a fundamental role in the pathophysiology of several heart diseases [3].
Chronic ethanol consumption is known to impair cardiac function [4,5] and significantly increase risk of cardiovascular disease [6]. Substantial evidence suggests that myocardial oxidative stress plays a primary role in the pathogenesis of alcoholic cardiomyopathy [7–9]. Ethanol treatment has been reported to increase lipid peroxidation [7,8], oxidatively damage mitochondrial DNA [10], and impair antioxidant defenses [11–13] in the heart. Furthermore, supplemental treatment with ROS-scavenging antioxidants has been shown to attenuate these adverse effects and preserve myocardial integrity [8,10,11].
Strenuous physical exercise has been reported to increase [14,15] or decrease [16] oxidative stress in the myocardium. Several studies indicate that chronic exercise results in an adaptive upregulation of the myocardial antioxidant system that serves to protect the heart against the excess ROS generated during subsequent bouts of exercise [16–18]. Others have reported, however, no change or even decreases in the activity of myocardial antioxidant enzymes following exercise training [15,19–21]. These equivocal findings are most likely due to variations in the mode and duration of the exercise stimulus, and inconsistent analytical procedures used to quantify myocardial antioxidant status.
Ethanol consumption is common among college athletes and recreational fitness enthusiasts who regularly engage in physical exercise [22]. While endurance exercise training has been shown to attenuate ethanol-induced myocardial lipid peroxidation and increase antioxidant enzyme activity in the heart [12], there have been no studies examining the effects of resistance exercise training on ethanol-induced oxidative stress in the heart. Resistance training is known to result in significant myocardial stress and elicits adaptations that differ from those observed following endurance training [23]. In light of the potential for strenuous exercise to increase or protect against myocardial oxidative stress, and the putative role of oxidative stress in ethanol-induced myocardial injury, it is important to investigate the combined effects of resistance exercise training and ethanol consumption on myocardial oxidative stress. Therefore, the purpose of this investigation was to examine the effects of resistance training and chronic ethanol consumption on lipid peroxidation and antioxidant status in the myocardium. We hypothesized that resistance exercise training would attenuate ethanol-induced increases in myocardial MDA and prevent an ethanol-induced depression of antioxidant defenses in the heart.
Materials and methods
Animal subjects
Forty male Sprague–Dawley rats (Harlan, Indianapolis, Indiana, USA) (mean body weight 464 ± 3 g) were stratified by body weight and randomized into one of four experimental groups: sedentary (SED; n = 8), sedentary + ethanol consumption (EtOH; n = 8), resistance training (RT; n = 12), or resistance training + ethanol consumption (RT–EtOH; n = 12). RT and RT–EtOH groups were larger to accommodate potential non-compliance with the exercise training protocols. Animals were housed individually in a temperature and humidity-controlled animal facility on a 12-h light/dark cycle. During the first week, all animals were acclimated to the Bio-Serve Liquid Rat Diet LD'82 (Bio-Serve, French-town, New Jersey, USA).
Alcohol treatment
Alcohol (95% ethyl alcohol; AAPER Alcohol and Chemical Co, Shelbyville, Kentucky, USA) concentration in the diet of EtOH rats was increased from an initial concentration of approximately 12–35% of total kilocalorie intake over a 1-week period. Alcohol diets were administered ad libitum throughout the 6-week study to the EtOH and RT–EtOH groups. Alcohol consumption was recorded daily and non-ethanol-receiving rats were pair-fed with their respective counterparts using the Bio-Serve Liquid Rat Diet LD'82 with a maltose dextrin caloric substitute in place of ethanol, according to the manufacturer's instructions. Blood alcohol content was determined spectrophotometrically in blood samples obtained at the time of sacrifice to confirm the presence of alcohol in the circulation using a commercially available assay kit (333-UV; Sigma Chemical, St Louis, Missouri, USA) using a Shimadzu (Kyoto, Japan) US-VIS Recording Spectrophotometer (UV2100U). All samples were assayed in duplicate and any samples that varied by more than 10% were reassayed.
Resistance training protocol
Animals from the RT and RT–EtOH groups were fitted with vests constructed of denim fabric and filled with varying amounts of lead pellets. Resistance exercise was conducted during the animals’ dark cycle under red light, in a plexi-glass cage with an electrical grid floor. All rats were acclimated to handling, wearing of weight vests and training in the ‘squatting cage’ for 2 weeks prior to the start of the study. Animals were trained to rise onto their hind limbs and stretch upwards to turn off a light. Cage tapping and mild electric shock were used sparingly to motivate animals to perform the exercises. After returning to a four-legged resting position, animals were motivated to rise onto their hind limbs again, bearing the full load of the weighted vest. Animals repeated this exercise 30 times per exercise session, 3 days per week. The load lifted was initiated at 20% body weight and increased by 20% per week over the 6-week training period to a final load of 120% body weight. Animals were sacrificed 24 h after the final exercise session by carbon dioxide inhalation. The rectus muscle was removed and weighed to establish an adaptive response to the resistance training protocol. The heart was rapidly removed, weighed, frozen in liquid nitrogen, and stored at – 80°C for future biochemical analyses.
Myocardial lipid peroxidation and antioxidant potential
Frozen cardiac tissue was pulverized in liquid nitrogen, homogenized in ice-cold 20 mmol/l tris-HCl, pH 7.4 (1 : 5 w/v) and centrifuged at 10 000 g for 10 min. Malon-dialdehyde (MDA), the most abundant end product of lipid peroxidation, was estimated in the supernatant spectrophotometrically at 586 nm on a Genesys 20 spectrophotometer (ThermoSpectronic, Rochester, New York, USA) using the Bioxytech MDA-586 assay kit (OxisResearch, Portland, Oregon, USA). Total antioxidant potential (AOP) of the supernatant was determined spectrophotometrically on a Spectramax 250 microplate reader (Molecular Devices, Sunnyvale, California, USA) using the Bioxtech AOP-490 assay kit (OxisResearch). The AOP assay is based on the reduction of 2189 mol/l Cu2+ to Cu+ by the combined effects of all antioxidants in the homogenized sample supernatant. Absorbance at 490 nm is recorded before and after addition of Cu2+ to the sample in the presence of bathouproine, which selectively combines with Cu+ to form a 2 : 1 chromogenic complex with a maximum absorbance at 490 nm. Uric acid was used as the standard, and thus sample AOP is expressed as uric acid equivalents (UAEs). All samples were assayed in duplicate and any samples that varied by more than 10% were reassayed.
Protein assay and statistical analysis
Myocardial MDA and AOP are expressed relative to total protein in the sample homogenates determined using the method of Bradford [23] with bovine serum albumin as the standard. All data are reported as mean ± standard error (SE). Two-way analyses of variance (ANOVA) were used to determine significant differences due to the main effects (exercise and ethanol consumption) and the interaction of these two factors. Independent sample t-tests were used to determine differences between individual groups. A significance level of P < 0.05 was established for all statistical analyses.
Results
Animal characteristics
Animal characteristics at the time of sacrifice are presented in Table 1. Nineteen of the 24 exercised animals were compliant with the resistance training protocol (nine from the RT group, and 10 from the RT–EtOH group) and data from non-compliant animals were excluded from analyses. There was no significant difference in the level of ethanol consumption between the EtOH (84.8 ± 0.2 ml/day) and RT–EtOH groups (83.8 ± 0.2ml/day). As expected, ethanol groups had significantly greater blood alcohol content than the non-ethanol groups at the time of sacrifice (P < 0.05). Body weight at the time of sacrifice was significantly lower in exercise-trained groups (P < 0.05), resulting from a mean body weight gain of only 54.9 ± 7.4 and 54.6 ± 2.9 g in the RT and RT–EtOH groups compared with a 70.5 ± 3.9 and 63.4 ± 5.3g gain in the SED and EtOH groups, respectively. This is consistent with data from Reed et al. [24], who reported that male rats do not compensate for increased physical activity by increasing their caloric intake. There was a general trend towards rectus hypertrophy in the exercise-trained groups, suggesting an adaptive skeletal muscle response to the resistance training protocol. Both resistance exercise training and ethanol treatment induced a trend towards cardiac hypertrophy. Heart weights relative to body weight were significantly greater in RT–EtOH than SED (P < 0.05). Heart and rectus weights were expressed per 100 g of body weight to correct for differences in body weight gain over the course of the study. Therefore, it is important to note that the observed increases in relative tissue weights may be due in part to training-induced reduction in overall body weight gain during the course of the study.
Myocardial lipid peroxidation
Figure 1 illustrates the effects of various treatments on cardiac MDA. Two-way ANOVA analyses indicated a significant effect of exercise (P < 0.01) and ethanol (P < 0.01) treatments on cardiac MDA, as well as an interaction between these main effects (P < 0.01). Trained groups exhibited significantly lower MDA levels than non-trained groups, while ethanol groups had greater cardiac MDA levels than the non-ethanol groups. Hearts from EtOH rats were found to have significantly greater levels of MDA (275 ± 61 pmol/mg of protein) than SED (97 ± 36 pmol/mg of protein) and RT (119 ± 32 pmol/mg of protein) animals, indicating significant ethanol-induced oxidative stress in the heart. Myocardial MDA levels in RT–EtOH were not significantly different from non-ethanol group levels (110 ± 64 pmol/mg of protein), suggesting a protective effect of resistance exercise training on ethanol-induced oxidative injury.
Animal characteristics at time of sacrifice
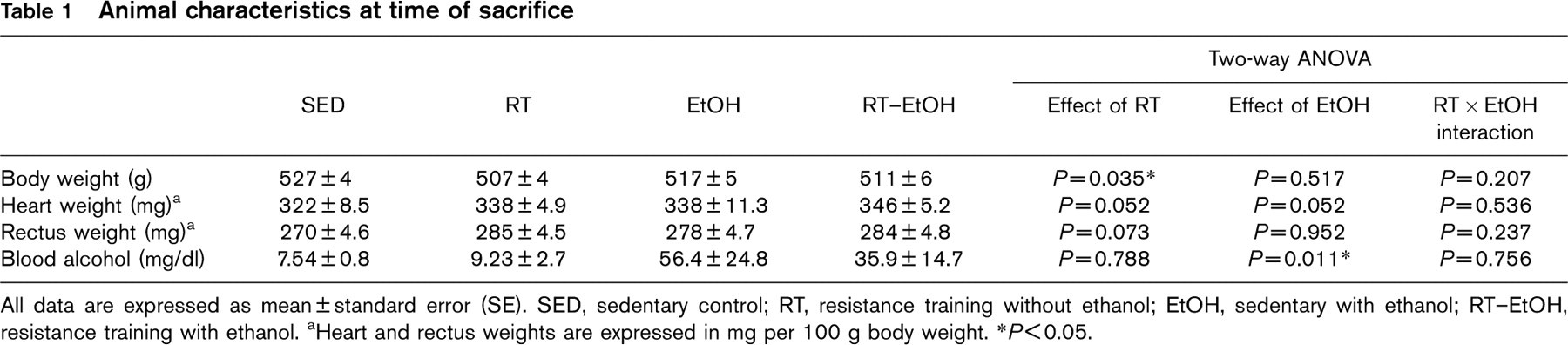
All data are expressed as mean ± standard error (SE). SED, sedentary control; RT, resistance training without ethanol; EtOH, sedentary with ethanol; RT–EtOH, resistance training with ethanol. aHeart and rectus weights are expressed in mg per 100 g body weight. ∗P≤0.05.
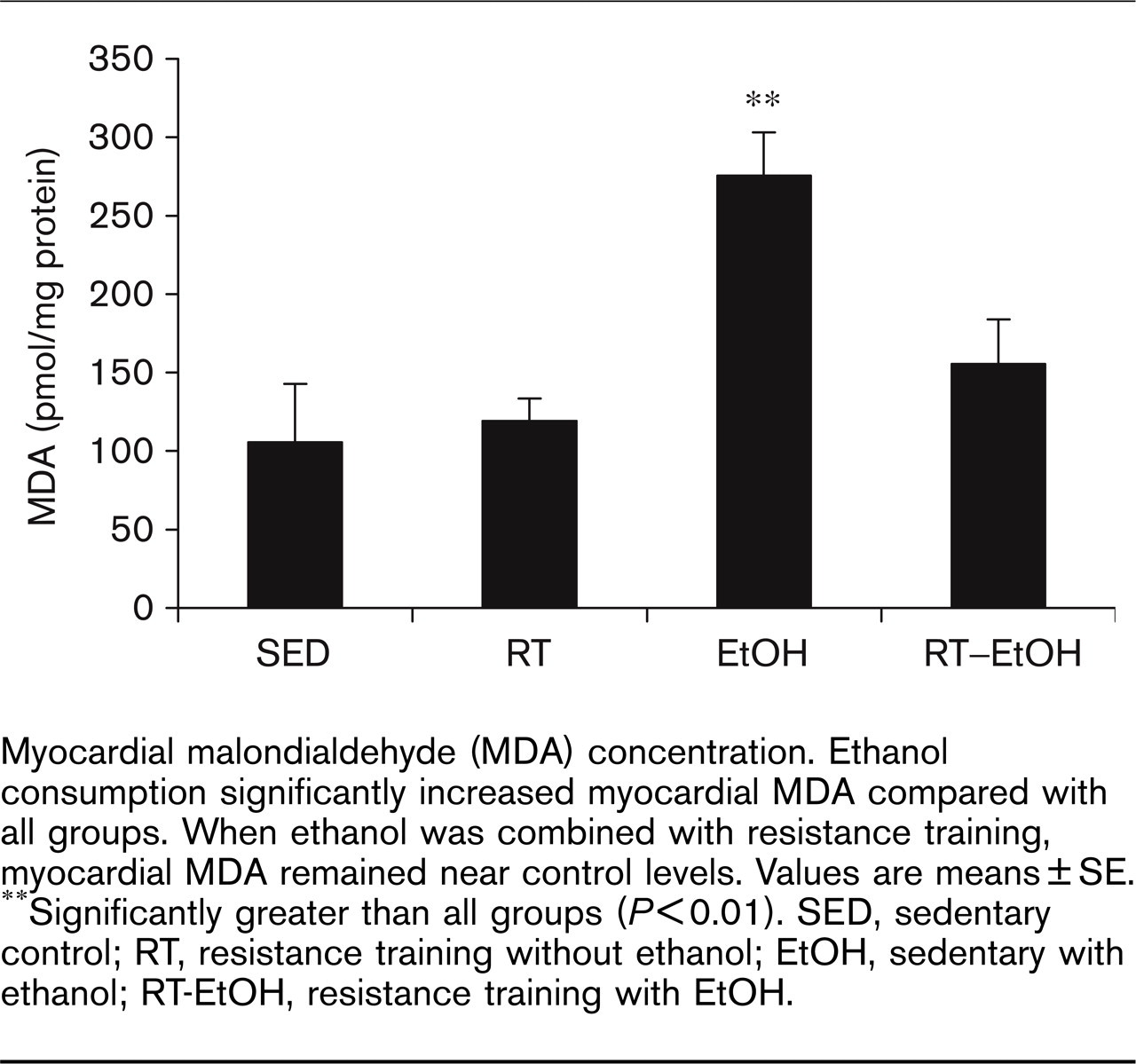
Myocardial malondialdehyde (MDA) concentration. Ethanol consumption significantly increased myocardial MDA compared with all groups. When ethanol was combined with resistance training, myocardial MDA remained near control levels. Values are means ± SE. ∗∗Significantly greater than all groups (P≤0.01). SED, sedentary control; RT, resistance training without ethanol; EtOH, sedentary with ethanol; RT-EtOH, resistance training with EtOH.
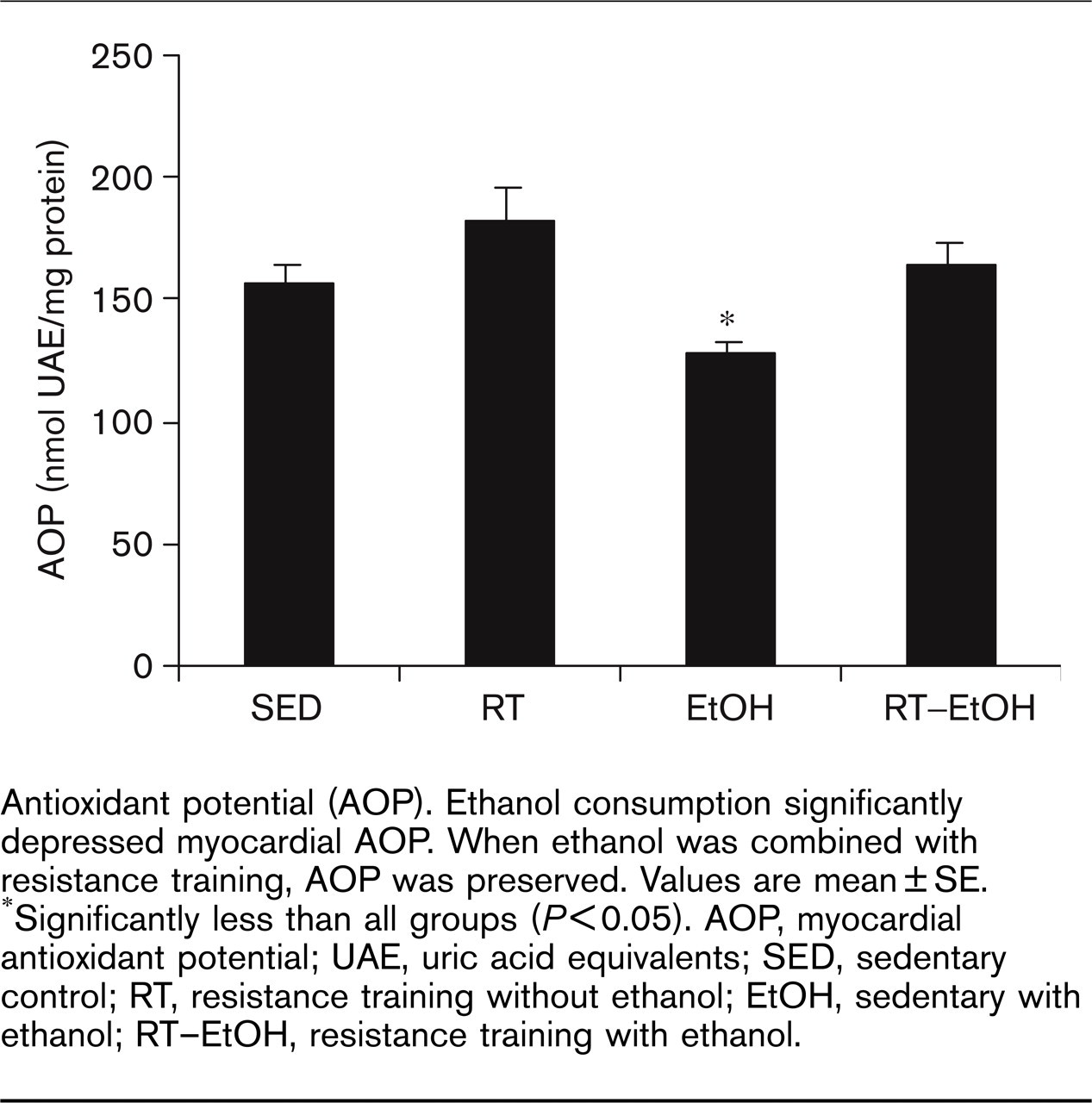
Antioxidant potential (AOP). Ethanol consumption significantly depressed myocardial AOP. When ethanol was combined with resistance training, AOP was preserved. Values are mean ± SE. ∗Significantly less than all groups (P≤0.05). AOP, myocardial antioxidant potential; UAE, uric acid equivalents; SED, sedentary control; RT, resistance training without ethanol; EtOH, sedentary with ethanol; RT–EtOH, resistance training with ethanol.
Myocardial antioxidant potential
Figure 2 illustrates the effects of resistance training and ethanol on myocardial AOP. Two-way ANOVA analyses indicated a significant effect of exercise (P < 0.05) and ethanol (P < 0.05) treatments on cardiac AOP, with no interaction between these main effects. Trained groups exhibited significantly greater AOP than non-trained groups. Ethanol groups were found to have significantly lower cardiac AOP levels than the non-ethanol groups, suggesting a suppressive effect of ethanol on cardiac AOP. When combined with resistance exercise training, this ethanol-induced reduction in AOP was prevented, suggesting that resistance training can preserve ethanol-induced suppression of myocardial AOP.
Discussion
The primary finding of this study is that resistance exercise training attenuates the increase in myocardial MDA and the decrease in myocardial AOP induced by chronic ethanol consumption. We have confirmed past findings that chronic ethanol consumption induces lipid peroxidation in the myocardium [7–9,25], and have added support to prior evidence that exercise training can protect against ethanol-induced oxidative injury in the heart [12]. To our knowledge, this is the first evidence for a cardioprotective effect of resistance training on ethanol-induced myocardial oxidative stress.
Several reports indicate that ethanol administration induces significant cardiac oxidative stress indicated by increased levels of myocardial lipid peroxidation [7,8,11,25], oxidative degradation of cardiac mitochondrial DNA [10] and depletion of myocardial glutathione [12,13]. In this study, we observed 1.6-fold higher levels of myocardial MDA in ethanol-treated rats than in non-ethanol controls, which is consistent with results obtained by others [25]. MDA is the stable end product of lipid peroxidation, a common form of cellular injury mediated by ROS and other free radicals. Ethanol-induced generation of ROS may be mediated by increased myocardial levels of xanthine oxidase [26] and increases in the activity of acyl CoA-oxidase [25]. Oxidation of short-chain alcohols by the cytochrome P450 mixed function oxygenases, particularly P4502E1 (CYP2E1), may also contribute to free radical production and injury in the myocardium [10,27].
Further evidence for a free radical mechanism of ethanol cardiotoxicity comes from studies that report cardioprotective effects of antioxidant treatment during or prior to ethanol administration. El-Sokkary et al. [8] found that subcutaneous injections of melatonin (10 mg/kg), a free radical scavenging antioxidant, 30 min before ethanol injections (3 g/kg subcutaneously daily for 30 days) completely prevented ethanol-induced increases in myocardial lipid peroxidation. Similarly, Mansouri et al. [10] reported that melatonin injections (10 mg/kg intraperitoneally) 30 min before intragastric administration of ethanol (5 g/kg) attenuated ethanol-induced degradation of cardiac mitochondrial DNA in mice. These damaging effects were also attenuated by daily intragastric administration of vitamin E (45 mg/kg) for 5 days before ethanol treatment. Interestingly, administration of 4-methylpyrazole (1 mmol/kg intraperitoneally), an inhibitor of both alcohol dehydrogenase and CYP2E1, 30 min before ethanol ingestion was more effective at preventing oxidative DNA damage than antioxidant administration, suggesting that ethanol metabolism may play a crucial role in ethanol-induced oxidative damage in the myocardium.
The myocardium normally protects itself against oxidative injury by the complex interaction of several endogenous enzymatic and non-enzymatic antioxidants capable of preventing cellular injury by interfering with damaging ROS reactions. Investigators frequently measure levels of free radical-scavenging molecules and the activities of one or more of the key myocardial antioxidant enzymes including superoxide dismutase (SOD), glutathione peroxidase (GPx) and catalase (CAT) in an effort to describe the antioxidant status of the heart. The effect of chronic ethanol consumption on myocardial antioxidants has been somewhat inconsistent. While ethanol treatment has been found to increase the activity of CuZnSOD [28] and CAT [25], others have reported decreases in myocardial glutathione [12,13] and total SOD activity [12]. The results reported herein indicate that ethanol consumption decreased the overall antioxidant potential of the heart, which was associated with higher levels of myocardial MDA in the ethanol group. This finding together with the aforementioned evidence for a cardioprotective effect of antioxidant supplementation [8,10] indicates that ethanol treatment may overload the ROS-neutralizing capacity of the endogenous myocardial antioxidant system, leaving the heart vulnerable to free radical injury.
We hypothesized that resistance exercise training may attenuate ethanol-induced increases in myocardial MDA by enhancing the myocardial antioxidant defense system. Several studies indicate that chronic endurance exercise protects the heart against oxidative stress [29] and reduces basal levels of myocardial lipid peroxidation [15–17]. While exercise-induced cardioprotection has often been associated with increases in myocardial antioxidant enzyme activity, specific changes in individual antioxidant molecules have been inconsistent [19,30]. Lui et al. [14] found that 8 weeks of treadmill running resulted in higher myocardial levels of α-tocopherol and ubiquinone in rats. Recently, Husain and Hazelrigg [17] reported increases in CAT, SOD, and GPx activity in the rat myocardium following 8 weeks of endurance training. Powers et al. [20] examined the effect of low, moderate and high intensity treadmill training in rats over 10 weeks and found that while myocardial SOD activity increased after all training protocols, no changes were observed in CAT or GPx activity after any of the training protocols. Ji [30] reported no changes in SOD, CAT, or GPx in the rat myocardium following 12 weeks of treadmill running. A study by Atalay et al. [31] provided one of the few examinations of chronic anaerobic exercise on the cardiac antioxidant system. These investigators reported increases in cardiac GPx activity following 6 weeks of high intensity sprint training, but found no changes in SOD activity. Venditti and DiMeo [15] utilized a method of assessing overall antioxidant capacity of the rat heart following 10 weeks of swim training. While they found that exercise training failed to increase GPx activity, it increased overall myocardial antioxidant capacity and reduced basal myocardial MDA. Collectively, these results suggest that the response of a single antioxidant enzyme is not indicative of the total antioxidant status of the myocardium, and that the effect of exercise training on myocardial antioxidants is highly variable. In light of the equivocal findings from studies examining the effects of ethanol and exercise on individual antioxidant enzyme molecules, and the fact that, to our knowledge, no studies have examined the effect of resistance training on myocardial antioxidants, we have employed a method of quantifying the total antioxidant capacity of myocardial tissue homogenates. The results of this study suggest that resistance training may prevent ethanol-induced decreases in myocardial antioxidant potential, which may serve to protect the heart against lipid peroxidation induced by ethanol consumption.
Few studies have examined the effects of exercise training on ethanol-induced myocardial oxidative stress. Husain and Somani [12] found that rats given 20% ethanol (2 g/kg) daily for 6.5 weeks had cardiac MDA levels 130% of sedentary controls, an effect that was abolished when ethanol was combined with treadmill exercise training. Ethanol also decreased the activities of SOD and GPx, but when combined with treadmill exercise training, GPx activity increased to 143% of sedentary controls. Exercise training alone did not significantly increase myocardial MDA or SOD protein levels. We have found in the current study that while resistance training alone did not significantly alter myocardial AOP or basal MDA concentration from SED levels, there was a significant effect of the resistance training treatment on myocardial AOP and MDA. In addition, when combined with chronic ethanol consumption, resistance training prevented an ethanol-induced reduction in myocardial AOP. We believe that this preservation of myocardial AOP is responsible, at least in part, for the prevention of ethanol-induced increases in myocardial MDA.
While an enhanced myocardial antioxidant potential could contribute to the protective effects of exercise against ethanol-induced oxidative injury, other mechanisms may be involved. Ardies et al. [32] reported that both acute and chronic treadmill running exercise increased rates of in-vivo ethanol clearance in rats. They suggested that chronic exercise enhanced ethanol metabolism by increasing the activity of the hepatic cytochrome P450 mixed function oxygenases. As mentioned above, evidence that CYP2E1 mediates the formation of ROS during ethanol metabolism [9], and that CYP2E1 inhibition protects the heart against ethanol-induced oxidative damage [10] support the notion that ethanol metabolism is fundamental in ethanol-induced oxidative injury. Evidence that exercise training can both enhance CYP2E1 activity and reduce myocardial oxidative injury seems somewhat paradoxical. Therefore, it is plausible that enhanced ethanol metabolism by the liver, an increased ethanol clearance rate, may reduce the level or duration of ethanol exposure in the heart. In this study, blood alcohol content was measured only at the end of the treatment protocol, so it is impossible to know the effects of resistance training on ethanol clearance rate throughout the experimental period. Interestingly, a trend towards lower blood alcohol content in RT–EtOH versus EtOH rats was observed at the time of sacrifice. Further investigation is needed to confirm an effect of resistance exercise training on ethanol metabolism and any role such an interaction may play in attenuating ethanol-induced myocardial oxidative stress.
In summary, our results suggest that resistance training protects against ethanol-induced oxidative stress in the myocardium. Ethanol consumption increased myocardial MDA and decreased myocardial AOP. When resistance training was combined with ethanol consumption, MDA and AOP levels were similar to those found in non-ethanol control hearts, suggesting that resistance training prevented ethanol-induced lipid peroxidation and preserved myocardial antioxidant defense capacity.