
Abstract
Background
In addition to the well established cardiovascular risk factors, evidence suggests a possible role of genetic and non-classical risk factors in the development and progression of atherothrombosis. We aimed to determine the relationship of classical and non-classical cardiovascular risk factors with candidate gene polymorphisms potentially involved in cardiovascular risk in the general Mediterranean population.
Design
Cross-sectional study.
Methods
We have determined the prevalence of classical (lipid profile, blood pressure, glycaemia, diabetes, smoking, body mass index, menopause and family history of coronary heart disease) and non-classical cardiovascular risk factors (infectious processes, homocysteinaemia, oxidative status, C-reactive protein, lipoprotein (a) and fibrinogen) in a population-based study. We analysed the relationship of these risk factors with the following five gene polymorphisms potentially involved in cardiovascular risk: ATP-binding cassette transporter A1-R219K, Peroxisome proliferator-activated receptor (PPAR)-α-L162V, Lipoprotein lipase (LPL)–HindIII, Paraoxonase (PON)1-Q192R, and Tumour necrosis factor (TNF)-α- G-308A.
Results
We found PPAR-α-V and LPL-H+ alleles to be associated with decreased high-density lipoprotein-cholesterol (HDL-c) concentration and with increased total cholesterol: HDL-c and triglyceride: HDL-c ratios. Regarding the non-classical risk factors, C-reactive protein concentration was higher for the PPAR-α-V allele. A higher oxidative status was shown in homozygotes for LPL-H+ and TNF-α-G alleles, although the latter also had lower homocysteinaemia.
Conclusions
Three of the genetic variants analysed, PPAR-α-L162V, LPL-HindIII, and TNF-α-G-308A, were associated with non-classical risk factors, specifically lipid profile, inflammation, and oxidative status.
Introduction
Atherosclerosis is the leading cause of death and disability in the developed world. The death rate from coronary heart disease (CHD) has declined in most industrialized countries in recent decades [1]. Our understanding of atherogenesis has evolved from a focus on lipid deposition within the arterial wall causing obstruction to a broader view of inflammatory and oxidative processes, which involves specific cellular and molecular responses to endothelial dysfunction [2]. In addition to the well-established cardiovascular risk factors of elevated total and low-density lipoprotein cholesterol (LDL-c), hypertension, diabetes and cigarette smoking, multiple additional factors are suspected culprits in both the development and progression of atherothrombosis. The recent observation that traditional risk factors for CHD fail to identify approximately 10% of the individuals who present the symptoms of acute coronary syndromes has stimulated interest in considering other determinants of the pathogenesis of atherosclerosis [3]. Heritability may account for 50% of risk factor prevalence [4–14]. Multiple evidence supports a major role for the following non-classical risk factors: infectious processes, homocysteinaemia, oxidative status, C-reactive protein, lipoprotein (a) and fibrinogen. Understanding gene-environment interactions may also facilitate preventive interventions to reduce CHD risk [15]. The non-classical risk factors may be useful for refining the risk estimations in individuals with an intermediate risk. CHD prevention based on a comprehensive risk factor evaluation should permit better decision-making [16]. On the other hand, gender differences in the prevalence and magnitude of the effects of these non-classical coronary risk factors have already been reported [2].
Risk factors are not homogeneously distributed and their effects vary among populations [17]. CHD mortality rates follow a decreasing north–south gradient in western Europe; the development of geographic epidemiological studies is considered essential to characterize the populations of European Mediterranean countries, where the high prevalence of classical risk factors does not produce expected CHD mortality rates [18]. The role of non-classical coronary risk and protective factors needs to be thoroughly analysed in the context of genetic studies to explain this paradox.
The aim of this study was to determine the relationship of classical and non-classical cardiovascular risk factors with several candidate gene polymorphisms potentially involved in cardiovascular risk in the Mediterranean population.
Methods
A cross-sectional study was designed with a two-stage population random sample, stratified into five 10-year age (25–34, 35–44, 45–54, 55–64 and 65–74 years) and sex groups. The response rate in this study was 73%. Between September 1994 and March 1996, 1748 participants were examined. Results and methods have been published in further detail elsewhere [18]. All established major cardiovascular risk factors [i.e. glycaemia, total cholesterol, high-density lipoprotein cholesterol (HDL-c), tryglycerides, lipoprotein (a), low-density lipoprotein cholesterol (LDL-c), blood pressure and smoking], along with height and weight, were measured by standard methods [18].
Laboratory analyses
Blood samples were collected after overnight fasting. Serum cholesterol (Roche Diagnostica, Basel, Switzerland) and triglyceride levels (Roche Diagnostica) were determined enzymatically. LDL-c was calculated by the Friedewald formula [19]. HDL-c was measured as cholesterol after precipitation of apolipoprotein B-containing lipoproteins with phosphotungstic-Mg2+ (Boehringer Mannheim, Mannheim, Germany).
Inter-assay coefficients of variation were 2.5, 4.5 and 3.2% for total cholesterol, HDL-c and triglycerides, respectively. Serum lipoprotein (a) [Lp(a)] concentration was determined by immunonephelometric testing (Immunotrub Lp(a), Immuno Diagnostica, Vienna, Austria).
Antibodies against Chlamydia pneumoniae (CPNEU-Ab) and cytomegalovirus (CMV-Ab) were determined in solid phase by enzyme-linked immunosorbent assay (ELISA) qualitative tests performed on microtitre plates (Chlamydia pneumoniae ELISA IgG, Vircell S.L., Granada, Spain; cytomegalovirus IgG enzyme immunoassay well, Radim, Rome, Italy).
Lipid peroxide assay was assessed by the generation of malonylaldehyde equivalents, and measured by the thiobarbituric acid reactive substances method (TBARS) [20].
Antibodies against oxidized LDL (OLAB) were measured in serum by ELISA, using copper-oxidized LDL coated on to microtitre strips as antigen and specific peroxidase-conjugated anti-human IgG antibodies to detect the presence of bound antibodies (OLAB; Biomedica, Vienna, Austria).
Homocysteine was detected by fluorescence polarity immunoassay (FPIA; Homocystein Imx System, Abbot, Oslo, Norway). C-reactive protein (CRP) concentration was determined by immunonephelometric testing (Tina-quant CRP; Roche Diagnostics, Mannheim, Germany). Fibrinogen concentration was determined by a coagulometric method in an optical autoanalyser (Toa Medical Electronics Co., Ltd, Kobe, Japan).
Genetic analyses
Genomic DNA was isolated from white cells. Several gene polymorphisms were selected for population geno-typing, based on a previously described relationship with CHD and on minimum population prevalence close to 10%. Allele frequencies for these polymorphisms were estimated by the gene counting method.
PON1-Q192R and Lipoprotein lipase (LPL)-HindIII genotypes were determined by standard methods described previously [21, 22]. ATP-Binding Cassette Transporter A1 (ABCA1)-R219K, Peroxisome Proliferator-Activated Receptor (PPAR)-α-L162V and Tumour Necrosis Factor (TNF)-α-G-308A were determined by Taqman assay (ABI Prism 7900HT; Applied Biosystems, Foster City, California, USA), using the set of probes and primers shown in Table 1.
Probes and primers for gene polymorphisms

Statistical analysis
All continuous variables are presented as mean (SD). When their distribution departed from normal, the median value and the first and third quartiles are presented. Categorical variables are presented as percentages. Differences between normally distributed group data were analysed by the unpaired Student's t-test as well as one-factor analysis of variance (ANOVA) and non-parametric tests when appropriate. The chi-squared test was used to compare categorical variables among groups. A chi-squared goodness of fit test was performed to investigate the hypothesis that the genotype frequencies follow the Hardy–Weinberg distribution. Statistical analysis was performed using SPSS 12 (SPSS Inc., Chicago, Illinois, USA).
Results
The mean age of participants was 51 years (SD 13.7). Among the 1748 participants aged 25–74 years, 838 (47.9%) were men.
Classical risk factors
The classical risk factors by sex are shown in Table 2. Risk factor differences between men and women followed expected patterns. Male subjects had higher values of blood pressure, glycaemia, triglycerides, LDL-c, log triglyceride: HDL-c and total cholesterol: HDL-c ratios, as well as a higher proportion of smokers and diabetics. Women had higher HDL-c levels than men. The remaining characteristics were similar in both sexes.
Non-classical risk factors
Non-classical risk factor distribution by sex is also shown in Table 2. Men had higher mean homocysteine levels and TBARS, and higher prevalence of CPNEU-Ab, than women. Women had more elevated levels of fibrinogen and OLAB, and a higher proportion of CMV-Ab. Median values of CRP and Lp(a) were similar in both sexes.
Risk factor characteristics of the representative sample of the Girona population aged 25–74 years by sex
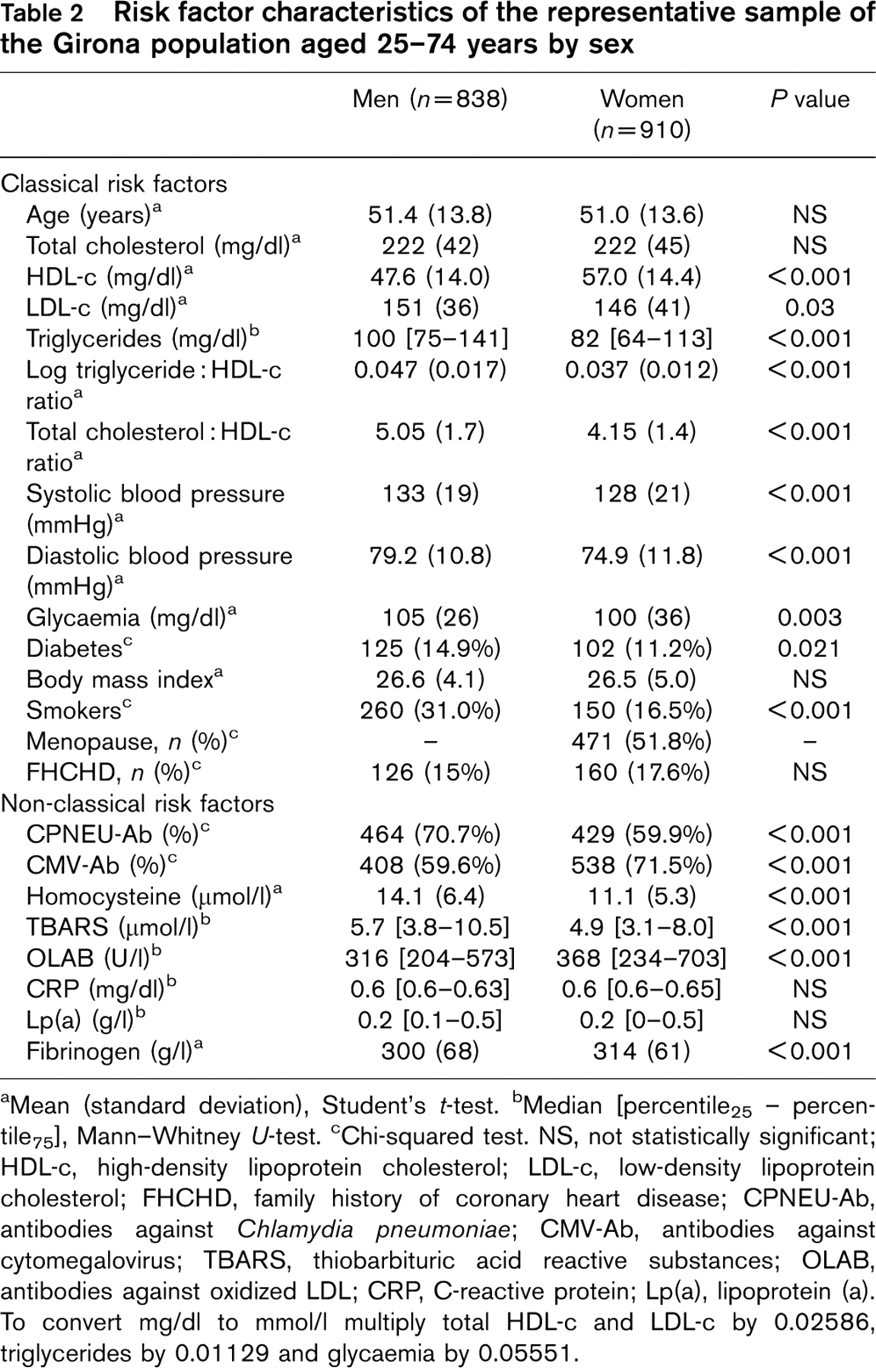
aMean (standard deviation), Student's t-test.
bMedian [percentile25 – percentile75], Mann-Whitney U-test.
cChi-squared test. NS, not statistically significant; HDL-c, high-density lipoprotein cholesterol; LDL-c, low-density lipoprotein cholesterol; FHCHD, family history of coronary heart disease; CPNEU-Ab, antibodies against Chlamydia pneumoniae; CMV-Ab, antibodies against cytomegalovirus; TBARS, thiobarbituric acid reactive substances; OLAB, antibodies against oxidized LDL; CRP, C-reactive protein; Lp(a), lipoprotein (a). To convert mg/dl to mmol/l multiply total HDL-c and LDL-c by 0.02586, triglycerides by 0.01129 and glycaemia by 0.05551.
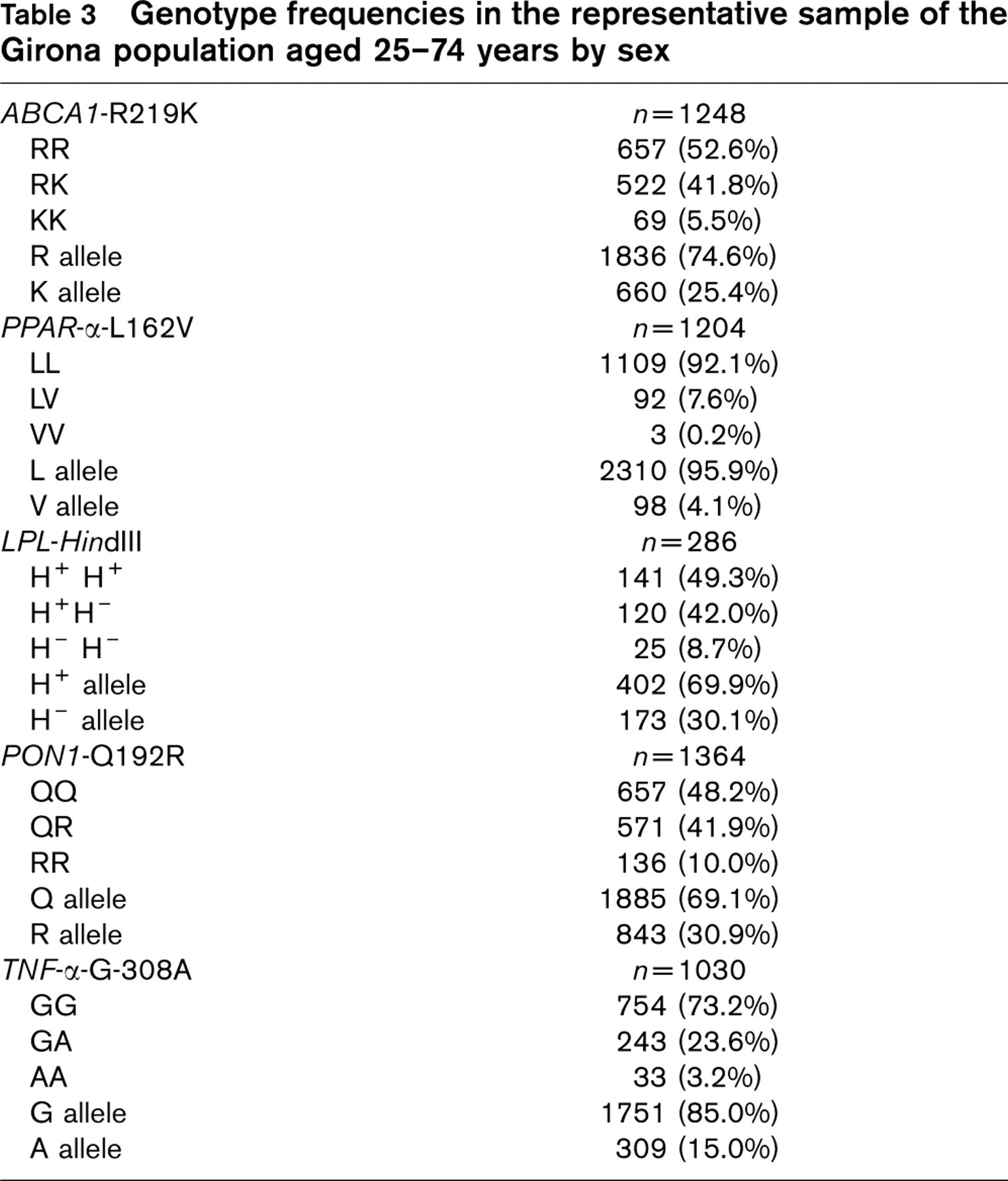
Genetic polymorphism frequencies
The observed genotype frequencies fitted those predicted by the Hardy-Weinberg distribution. The allelic frequencies of gene polymorphisms are shown in Table 3.
Relationship of genetic polymorphisms with classical risk factors
The relationship between the phenotype for several classical risk factors and the genotype of gene polymorphisms is presented in Table 4. The H−-carrier genotype of the LPL-HindIII polymorphism was associated with a lower log triglyceride: HDL-c and total cholesterol: HDL-c ratios, but higher mean of HDL-c. The Vallele of the PPAR-α-L162V polymorphism showed higher log triglyceride: HDL-c and total cholesterol: HDL-c ratios, but lower mean of HDL-c.
Genotype frequencies in the representative sample of the Girona population aged 25–74 years by sex
Relationship of genetic polymorphisms with non-classical risk factors
The relationship between genetic polymorphisms and non-classical risk factors is shown in Table 4. The CRP median values were lower in participants with the LL genotype of the PPAR-α-L162V polymorphism; TBARS median values were lower in participants with the H−-carrier genotype of the LPL-HindIII polymorphism. Finally, participants with the A-carrier genotype of the TNF-α-G-308A polymorphism had lower TBARS and higher homocysteine levels than the other genotypes.
Discussion
In the present study, we aimed to explore the prevalence of non-classical cardiovascular risk factors, and whether they were associated with genotypic variation in a number of candidate genes for cardiovascular risk in a large population-based sample. Non-classical risk factors were selected from those discussed in the past decade with varying results in their relationship with CHD; gene polymorphisms were selected for their suggested relationship with CHD (i.e. lipid metabolism, inflammation, and oxidation control) with a previously described allele frequency > 5%.
The prevalence of classical cardiovascular risk factors in the present study was similar to that found in other studies in a Spanish population [18, 23]. The REGICOR Study has shown a paradox consisting of a high prevalence of cardiovascular risk factors and low incidence of myocardial infarction in the population of Girona, Spain [18]. In fact, the classical cardiovascular risk profile of this population is worse than that of northern European populations. This paradox in the Spanish population could be related to lower prevalence of non-classical cardiovascular risk factors, as compared with populations of countries with a high incidence of CHD.
A necropsy study [24] in young Spanish men reported a high incidence of atheroma plaques in the anterior descendent coronary artery, similar to that found in populations with much higher rates of CHD. The key points were that most lesions were stable fibrous plaques and that no vulnerable plaques were found in this study. This is consistent with our hypothesis that unknown protective factors may prevent coronary plaques from becoming unstable in subjects in our geographic area. Therefore, interesting perspectives arise from the study of the prevalence of non-classical coronary risk factors to better understand this Spanish or southern European paradox.
The prevalence of CPNEU-Ab in our population (see Table 2) is high compared with that reported in the Atherosclerosis Risk in Communities (ARIC) study [25], which was approximately 55%, but lower than that observed in northern European populations [26]. The CRP concentrations are either slightly higher than those reported in some northern European populations [27] or lower than those observed in other populations from similar areas [28], leading to inconsistent findings. Although comparison of homocysteine data is difficult unless the interlaboratory differences are known, the present study found lower levels of homocysteine than those described in an English population, and similar to those of a Dutch population study [29, 30]. Fibrinogen concentrations were similar to those described in a white English population [31]. These observations are consistent with our hypothesis of a southern Europe paradox with higher risk factor prevalence (regardless of being classical or not) and low incidence of acute myocardial infarction.
Gender differences in classical and non-classical cardiovascular risk factors were also found in the present study. Men had significantly higher levels of classical cardiovascular risk factors (i.e. LDL-c, triglycerides, log triglyceride: HDL-c and cholesterol: HDL-c ratios, systolic blood pressure, diastolic blood pressure, glycaemia, diabetes prevalence and smoking) while women showed higher levels of some non-classical cardiovascular risk factors (i.e. CMV-Ab, OLAB and fibrinogen) and decreased levels of others (i.e. CPNEU-Ab, homocysteine and TBARS). It has been reported previously that oestrogens appear to increase CRP and fibrinogen levels and decrease homocysteine level and lipid oxidation status [2]. The concentration of OLAB, which is inversely correlated with lipid oxidation [32], was higher in women than in men in our study, suggesting that oxidized LDL may be different between sexes. In fact, this is supported by our finding that TBARS, a non-specific oxidative stress measurement, is significantly lower in women. Although the data published so far are judged inconclusive concerning the correlation between clinical evidence of arteriosclerosis and the levels of antibodies to oxidized LDL, some observations support the hypothesis that the immune response to oxidized LDL may have a protective role at an early stage of human atherosclerosis [33]. The CPNEU-Ab prevalence is consistently higher worldwide in adult men than in women [25].
Risk factor characteristics by ABCA1, PPAR-α, LPL, PON1 and TNF-α genotypes in the representative sample of the Girona population aged 25–74 years
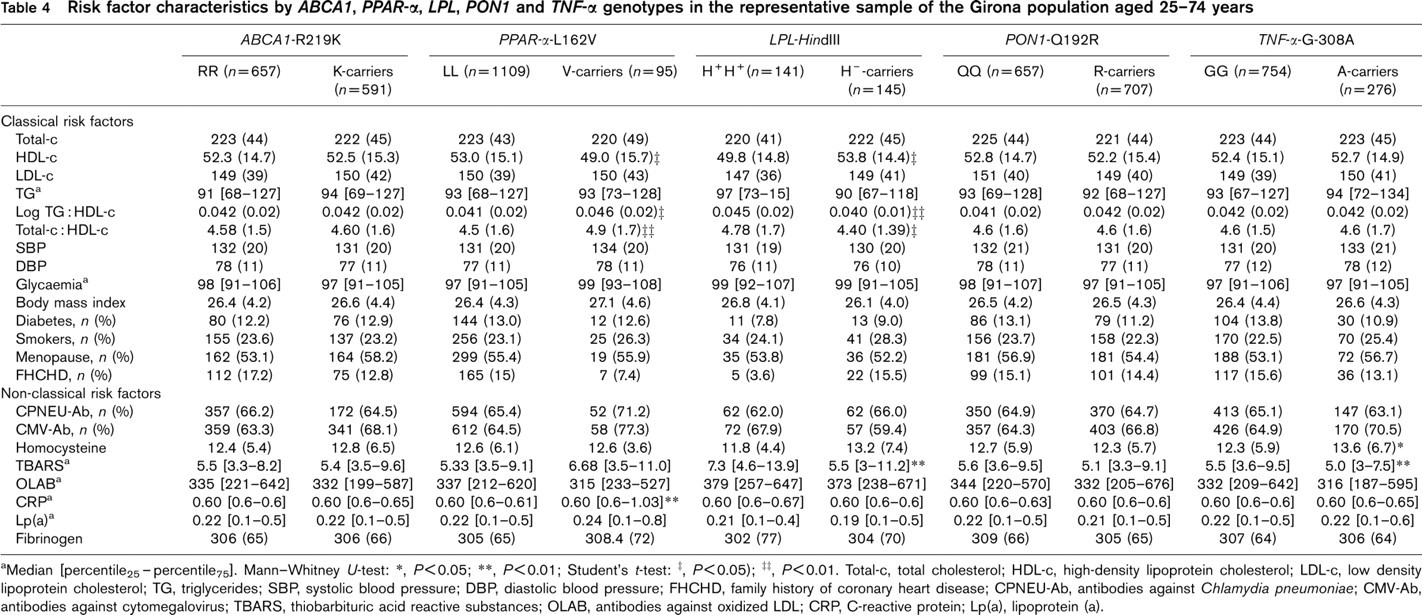
aMedian [percentile25-percentile25]. Mann–Whitney U-test:
∗, P<0.05
∗∗, P<0.01
Student's t-test:
‡, P<0.05
‡‡, P<0.01.
Total-c, total cholesterol; HDL-c, high-density lipoprotein cholesterol; LDL-c, low density lipoprotein cholesterol; TG, triglycerides; SBP, systolic blood pressure; DBP, diastolic blood pressure; FHCHD, family history of coronary heart disease; CPNEU-Ab, antibodies against Chlamydia pneumoniae; CMV-Ab, antibodies against cytomegalovirus; TBARS, thiobarbituric acid reactive substances; OLAB, antibodies against oxidized LDL; CRP, C-reactive protein; Lp(a), lipoprotein (a).
There is general agreement that the influence of single genetic polymorphisms on CHD development is slight at the population scale, because they can contribute little to the overall burden of atherosclerosis. However, single nucleotide polymorphism analysis may be useful for calculating individual CHD risk [34]. Some recent observations suggest that a number of relevant genetic variants in candidate genes, coupled with environmental and phenotypic risk factors, produce models that may be important for future risk assessment [35].
Identification of a genetic north-south gradient among European populations may be important to a better understanding of the CHD mortality rate differences in Europe. In general, our analysis of genotype or allele frequencies drew similar results compared with those reported in other genetic epidemiology studies in European populations [23, 36]. Nevertheless, some aspects merit consideration. In the present study, the prevalence of the 162 V allele of the PPAR-α-L162V polymorphism, which appears to be associated with a more harmful lipid profile and higher BMI but not with atherosclerosis progression [37], was lower than that observed in a Finnish population [38]. A decreasing frequency in the H+ allele of the LPL-HindIII polymorphism has been described from northern to southern Europe [36]. In our study, the H + allele frequency was higher (roughly 70%) than that described in other southern European countries: approximately 62% [36]. The TNF-α G allele and the PON1 R allele were more often observed in the Belfast population than in our population, which has one-quarter the CHD morbidity and mortality rates of Belfast [39]. These findings, together with differences in lifestyle, might contribute to explaining part of the geographical variability in CHD incidence and mortality rates observed in Europe.
In our study, the association of selected gene polymorphisms and risk factors is limited to lipidic and some inflammation and oxidation characteristics.
The association of the H+H+ genotype of the LPL-HindIII polymorphism with an observed adverse lipid profile concurs with other analyses of Spanish population studies [40]. The H+H+ genotype of the LPL-HindIII polymorphism and the A allele of the TNF-α-G-308A polymorphism were associated with higher TBARS concentration, consistent with a poor antioxidant/oxidant balance. The Vallele of the PPAR-α-L162V polymorphism was found to be associated with higher CRP levels. This fact may be related to a high level of circulating interleukins and hence a pro-inflammatory state. Finally, the relationship between high homocysteine levels and the A allele of the TNF-α-G-308A is unclear.
Study characteristics
For at least some of the non-classical risk factors there is some degree of discussion as to whether they are indeed risk factors or not. In this respect, the recently reported Vitamin Intervention for Stroke Prevention (VISP) Study [41] cast doubt on homocysteine as a risk factor. On the other hand, the recently presented Azithromicin and Coronary Events Study (ACES) [42] found no association of azithromycin treatment with reduced risk of CHD, raising questions about the role of infection in atherosclerosis. Furthermore, it is not clear whether CRP is a marker or a causative factor of atherosclerosis. These observations may constitute a possible limitation of the study; however, this limitation is difficult to address until definite conclusions are reached.
The present work relies on cross-sectional descriptive data and is essentially descriptive in nature. However, genetic variants, which are unchangeable characteristics, would have the value of a retrospective cohort study in phenotype analyses. An added value includes the description by gender of classical and non-classical cardiovascular risk factors in a Mediterranean population. Given the polygenic and multifactorial nature of cardiovascular diseases, future studies should comparatively analyse the effect of genotypic associations with environmental factors on particular phenotypes (i.e. classical and non-classical risk factors) in different populations.
In conclusion, three of the genetic variants analysed, PPAR-α-L162V, LPL-HindIII, and TNF-α-G-308A, were associated with non-classical risk factors, specifically lipid profile, inflammation and oxidative status.