
Abstract
Background
Chronic otitis media with effusion (COME) develops after sustained inflammation and is characterized by secretory middle ear epithelial metaplasia and effusion, most frequently mucoid. Staphylococcus epidermidis, typically considered a commensal organism, is very frequently recovered in chronic middle ear fluid and in middle ear biofilms. Although it has been shown to drive inflammation in sinonasal epithelium, the impact of S. epidermidis on COME is markedly understudied. The goal of this study was to examine the in vitro effects of S. epidermidis lysates on murine and human middle ear epithelial cells.
Methods
Staphylococcus epidermidis lysates were generated and used to stimulate submerged and differentiated human and murine epithelial cells (MEECs) for 24 to 48 hours. Quantitative real time-polymerase chain reaction, Western blot, enzyme-linked immunosorbent assay, and immunocytochemistry techniques were performed to interrogate the mucin gene MUC5AC and MUC5B expression and protein production, chemokine response, as well as NF-κB activation. Luciferase reporter assays were performed to further evaluate nuclear factor κB (NF-κB) activation and query specific promoter responses after S. epidermidis exposure.
Results
Staphylococcus epidermidis induced a time- and dose-dependent MUC5AC and MUC5B overexpression along with a parallel overexpression of Cxcl2 in mouse MEEC and IL-8 in human MEEC. Further investigations in mMEEC showed a 1.3 to 1.5 induction of the MUC5AC and MUC5B promoters. As potential mechanisms for these responses, induction of an oxidative stress marker, along with early nuclear translocation and activation of NF-κB, was found. Finally, chronic exposure induced marked epithelial thickening of cells differentiated at the air liquid interface.
Conclusions
Staphylococcus epidermidis lysates activate a proinflammatory response in MEEC, including mucin gene expression and protein production. Although typically considered a nonpathogenic commensal organism in the ear, these results suggest that they may play a role in the perpetuation of an inflammatory and mucogenic response in COME.
Staphylococcus epidermidis, typically considered a commensal nonpathogenic organism, has, for a long time, been known to be present in “sterile” chronic middle ear fluid9,10 and, more recently, been shown to be a very common, if not the most common, pathogen recovered from middle ear biofilms associated with COME.11,12 Along the same lines, S. epidermidis biofilms are frequently encountered in tympanostomy tubes 13 and middle ear ossicle explants. 14 Chronic otitis media with effusion fluid is characterized by the presence of mucin glycoproteins, which are the macromolecules primarily responsible for the viscoelastic properties of middle ear fluid. The 2 major airway mucin glycoproteins are MUC5AC and MUC5B. Although MUC5B seems to be predominant in human middle ear effusion,15,16 both of these proteins have been identified in the middle ear epithelium. 17 Indeed, MUC5AC is the most extensively studied mucin glycoprotein in the middle ear epithelium.
Despite the fact that S. epidermidis is one of the most common organisms cultured from middle ear mucinous fluids, how this bacterial species may actually affect middle ear inflammatory responses or the cellular production of mucin glycoproteins has not been studied to date. Interestingly, in the nasal epithelium, S. epidermidis has been shown to induce a potent proinflammatory response characterized by activation of the transcription factor nuclear factor κB (NF-κB) and subsequent overexpression of IL-8. 18 Both NF-κB and the functional homologue of IL-8 in mice, Cxcl2, have been shown to be important mediators of COME mucin responses.19,20
For these studies, we hypothesized that S. epidermidis lysates would induce a proinflammatory response in the middle ear epithelium similar to the one shown in the nasal epithelium, with an activation of NF-κB, an oxidative stress response, and induction of IL-8/Cxcl2. Further, we aimed to determine whether this response correlated to an overexpression and overproduction of MUC5B and MUC5AC.
Methods
Preparation of S. Epidermidis Lysates
Lysates from S. epidermidis, American Type Culture Collection strain 35984, were prepared from cultures under conditions for growth on chocolate agar at 37°C in 5% CO2 overnight and inoculated in brain-heart infusion broth supplemented with 10 mg of nicotinamide adenine dinucleotide per milliliter as described previously for nontypeable Haemophilus influenzae (NTHi). 19 After sonication for lysis, stock solutions of S. epidermidis were in the 4-mg/mL range. The use of lysates was chosen because this is what is released in children after antibiotic treatment for acute OM and is what the vast majority of groups have used to study middle ear and respiratory inflammation effects of other bacterial agents such as NTHi.21,22
Cell Lines
Immortalized mouse middle epithelial cell line (mMEEC) and the immortalized human middle ear epithelial cell line (HMEEC-1) were both used. The mMEEC cell line was immortalized with a temperature-sensitive SV40 virus allowing for cell differentiation at 37°C and was provided to us by Dr. Jizhen Lin (University of Minnesota, Minneapolis, MN). 23 The mMEECs were maintained and passaged in full growth media (FGM) as previously described. 24 The HMEEC-1 is a human immortalized middle ear epithelial cell line described by Chun et al., 25 which was generously been provided to us by Dr. David Lim's laboratory at the House Ear Clinic in Los Angeles, CA. These cells were cultured in bronchial/tracheal epithelial cell growth medium (BEGM; Clonetics, Walkersville, MD), supplemented with 0.1-μg/mL human recombinant epidermal growth factor, 5.0-mg/mL insulin, 0.5-mg/mL hydrocortisone, 50-mg/mL gentamycin, 50-μg/mL amphotericin B, and 7.5-mg/mL bovine pituitary extract at 37°C in 5% CO2. For experimentation, both cell lines were cultured on plastic submerged in media and also under differentiating conditions after 3 weeks of growth on collagen-coated transwells at air-liquid interface (ALI) as previously described. 24
Real-Time Reverse Transcriptase-Polymerase Chain Reaction
Total RNA was extracted from cells exposed to S. epidermidis lysates (0–200 μg/mL) or vehicle (cell culture media) for specific time points using TRIzol reagent (Life Technologies Corp, Grand Island, NY). Reverse transcription reaction was performed using 2 μg of total RNA from each sample and the SuperScript III reverse transcriptase enzyme (Invitrogen, Carlsbad, CA), as previously described. 24 Generated complementary DNA was used for quantitative PCR (qPCR), specific pairs of primers that were used are listed in Table 1. β-actin primers obtained from GeneLink (Hawthorne, NY) were used as an internal control for mouse genes. Real time reverse transcriptase-PCR (RT-PCR) was performed on the generated complementary DNA products in the ABI Prism 7900 Fast RT sequence detection system (Applied Biosystems, Foster City, CA) as described previously. 24 Relative quantification of the expression of the genes of interest was obtained using the ΔΔCt method developed by Pfaffl. 26
Primers Used for PCR

Luciferase Assays
The pIgκB Luc reporter construct containing 3 concensus NF-κB binding sites upstream of the luciferase gene has been previously described24,27 and was generously provided by Frank Ondrey, MD, PhD, of the University of Minnesota, Minneapolis, MN. The MUC5B promoter plasmid was cloned in our laboratory as previously described. 20 The MUC5AC promoter plasmid was obtained from Dr. Mary Rose at Children's National Health System. 28 The NF-κB reporter plasmid and the MUC5B promoter plasmid were transiently transfected into mMEEC cells for luciferase assays as follows. Cell line cultures at 50% to 60% confluence were cotransfected with the luciferase plasmids (2 μg/mL) and a pCMV-βGal reporter construct (0.05 μg/mL) (Clontech, Mountainview, CA) in Opti-MEM medium containing 3 μg/mL of Lipofectamine 2000 (Invitrogen, Carlsbad, CA). After 16 hours, the medium was removed and the cells were placed in FGM and allowed to seed for 8 hours before treatment with S. epidermidis in serum-free media or medium alone for 16 hours. The relative luciferase activity was then determined with the dual-light reporter gene assay (Tropix, Medford, MA) and a Mythras plate luminometer (Berthold, Oak Ridge, TN) according to the manufacturers’ instructions. Results for relative luciferase units were determined as a ratio of the luciferase constructs over the pCMV-βGal reporter to normalize for harvesting and transfection efficiency.
Western Blotting for Human MUC5B Protein
Gel electrophoresis from human middle ear cell lysates (40 μg per well) was carried out as previously described in 1% agarose gels. 19 After electrophoresis and gel transfer, Western blotting was carried out as previously described. 19 The primary polyclonal antibody for MUC5B (Santa Cruz, CA), H-300, was used at a 1:300 dilution. The horseradish fluoroilluminescence detection protocol using SuperSignal West Dura Extended Duration Substrate (Pierce, Rockford, IL) was used to visualize protein signal. The developed blots were exposed and visualized with a charge-coupled device camera equipped Gel Doc 2000 chemiluminescent imaging documentation station (Bio-Rad, Hercules, CA).
Preparation of Nuclear Extracts
The mMEEC and HMEEC grown to 80% confluence in FGM were stimulated for specific time points with 0 to 200 μg/mL of S. epidermidis lysates. Cell suspensions were prepared by Trypsin-EDTA treatment, and cytoplasmic and nuclear protein extraction was then performed using a kit from Pierce (Rockford, IL) as previously described 24 according to the manufacturer's recommendations.
Elisa for Nf-κB Nuclear Translocation
The nuclear and cytoplasmic extracts of cells cultured in 6-well Transwell plates at ALI and treated with or without S. epidermidis lysates were assayed for p65 protein quantity using the InstantOne ELISA assay (eBioscience, San Diego, CA) following the manufacturer's recommendations. Extracts (50 μL) were added to each well of the plate. At the end of the reaction, the absorbance of the wells was read at 450 nm using an xMark microtiter plate reader (Biorad, Richmond, CA). The results were then normalized to the quantity of each extract, and the ratio nuclear extract/cytoplasmic extract was calculated.
Immunofluoresence Detection of p65 Protein
The mMEEC and HMEEC in FGM were cultured to 80% confluence in glass chamber slides (Thermo Scientific, Belfonte, PA) before the treatment of S. epidermidis lysates. At the end of the treatment, the cells were rinsed 3 times with phosphate buffered saline (PBS), fixed in paraformaldehyde 3% for 20 minutes, washed 3 times with PBS, and stored at 4°C. Then, the immunostaining was performed. Cells were permeabilized with PBS/Tween 20 0.05% for 5 minutes, and the nonspecific sites were saturated using PBS/Tween 20 0.01%/bovine serum albumin 3% for 30 minutes. This solution was then used for the incubation with the primary antibody p65 NF-κB (C-20) sc-372 used at 1/200 (from Santa Cruz, CA) and the secondary antibodies (Anti-goat Alexa 488 A11055, Anti-rabbit Alexa A10042) used at 1/500 (Invitrogen, Carlsbad, CA). DAPI 1:8000 in PBS was used to stain the DNA. The chambers were removed, and the slide was mounted using Permount mounting medium (Fisher Scientific, Suwanee, CA) with a coverslip. The slides were stored at 4°C in dark before analyzing using an Olympus FV1000 confocal microscope (Olympus, Rocklin, CA).
Histopathological Evaluation of Middle Ear Cells Grown at ALI
The mMEEC cells were cultured on Transwell plates at air-liquid interface and treated with serum-free medium versus 200-μg/mL S. epidermidis lysate in serum-free medium 2 times 2 hours at 48-hour interval (time point, 96 hours) or 6 times 2 hours at 48-hour interval (2-week time point) only at the apical side (1 mL per well). The time of exposure in the apical side was limited to 2 hours because we observed cell death for longer exposures with even medium alone in the apical side of cells cultured at ALI. For histology, cells were fixed with 10% formalin for 24 hours and embedded in paraffin before sectioning.
Sds-Page Electrophoresis and Peptide Preparation for Mass Spectrometry Analysis
Staphylococcus epidermidis lysates (50 μg) were dissolved in Laemmli buffer containing 0.1-mM DTT and were run in a 1-dimensional SDS gel electrophoresis gel at 200 V for 50 minutes. The gel was fixed with methanol and stained with Coomassie for protein visualization. Each gel lane was sliced into 30 segments, and each slice was digested with trypsin as follows. Briefly, the gel cuts were placed in 100 μL of water and then subjected to 2 washes with a 1:1 by volume solution of water and acetonitrile. The gel pieces were then dehydrated with acetonitrile and rehydrated using 100-mM ammonium bicarbonate, followed by a 1:1 by volume wash of 100-mM ammonium bicarbonate and acetonitrile. The gels were then dehydrated with acetonitrile, resuspended in digestion buffer containing 12.5 ng/μL of mass spectrometry (MS) grade Trypsin Gold (Promega Corp, Madison, WI), and incubated overnight at 37°C. Extraction of peptides from the gel was then conducted via 2 washes with 25 mM of ammonium bicarbonate, followed by 2 washes with a 1:1 by volume solution of 5% formic acid and acetonitrile. The extracted peptides were then completely dried in a SpeedVac (ThermoScientific, Waltham, MA).
MS and Protein Identification
Dried peptides were resuspended in 10 μL of 0.1% trifluoroacetic acid. Each sample (6 μL) was injected via an autosampler and loaded onto a C18 trap column (5 μm, 300 μm i.d. × 5 mm, LC Packings) for 10 minutes at a flow rate of 10 L/min, 100% A. The sample was subsequently separated by a C18 reverse-phase column (3.5 μm, 75 μm × 15 cm, LC Packings) at a flow rate of 250 nL/min using an Eksigent Nano-HPLC System (Dublin, CA). The mobile phases consisted of water with 0.1% formic acid (A) and 90% acetonitrile with 0.1% formic acid (B). A 65-minute linear gradient from 5% to 60% B was used. Eluted peptides were introduced into the mass spectrometer via a 10-μm silica tip (New Objective Inc, Ringoes, NJ) adapted to a nanoelectrospray source (Thermo Fisher Scientific). The spray voltage was set at 1.2 kV; the heated capillary, at 200°C. The linear trap quadrupole (LTQ) mass spectrometer (ThermoFisher Scientific) was operated in data-dependent mode with dynamic exclusion in which 1 cycle of experiments consisted of a full-MS (300–2000 m/z) survey scan and 5 subsequent MS/MS scans of the most intense peaks.
Statistical Analysis
The statistical difference between experimental and control groups for all experiments was determined using 2-tailed Student t tests, analysis of variance test, followed by Dunnett test or Wilcoxon tests. Significance level was set at P < 0.05.
Results
S. Epidermidis Lysates Induce Mucin Gene Expression and Protein Production in Middle Ear Epithelial Cells
The RT-PCR data from experiments performed on submerged cells demonstrated that S. epidermidis lysates (3 different batches) increased murine MUC5B and MUC5AC messenger RNA (mRNA) levels in mMEEC at 200 μg/mL (P < 0.05 for MUC5B and P = 0.0625 for MUC5AC) at 24 hours and 100 μg/mL at 48 hours (P < 0.05) (Fig. 1, A and B). The graphs show the same variations for MUC5B and MUC5AC as well as a variability of the results due to the use of different batches of S. epidermidis lysates that have been characterized by proteomic analysis in the supplementary data (Supplemental Table 1, Supplemental Digital Content 1, http://links.lww.com/JIM/A23). The more reactive batch was chosen for the other experiments. These effects on MUC5B and MUC5AC mRNAs were correlated to a 1.2- and 1.4-fold induction of the pMuc5bLuc promoter after 100 and 200 μg/mL of S. epidermidis lysate exposure and a similar 1.4- and 1.3-fold induction of the pMuc5acLuc promoter with the same doses (all P < 0.05) (Fig. 1C). The mMEECs grown under differentiating conditions at ALI showed a stronger mucin response at 48 hours for both MUC5B (5.2-fold) and MUC5AC (12.2-fold) (all P < 0.05), which was sustained at 96 hours for both mucins, but leveled off to control levels of mucin expression after 2 and 3 weeks of lysate exposure (Fig. 1D). Taken together, these data indicate that these murine middle ear epithelial cells overexpress mucin genes upon exposure to S. epidermidis lysates in a time- and dose-dependent manner, with 48 hours of exposure eliciting the maximal level of mucin gene activation. Over time (2–3 weeks) on differentiating conditions, these cells constitutively express mucins over baseline, with lysate stimulation making little difference in terms of mucin gene expression. However, after 2 weeks of exposure, there is marked influence in the thickness of the epithelial layer, with the lysates inducing a dramatic 5- to 10-fold increase in the thickness of mMEEC cultures at ALI over control, whereas no difference was noted after 96 hours of exposure corresponding to 2 treatments with S. epidermidis lysates (Fig. 1E). In summary, results indicate that release of these lysates in the middle ear space is likely to produce an early (1–2 days) mucogenic response in patients with OM, with the potential for middle ear epithelial hyperplasia upon sustained exposure.
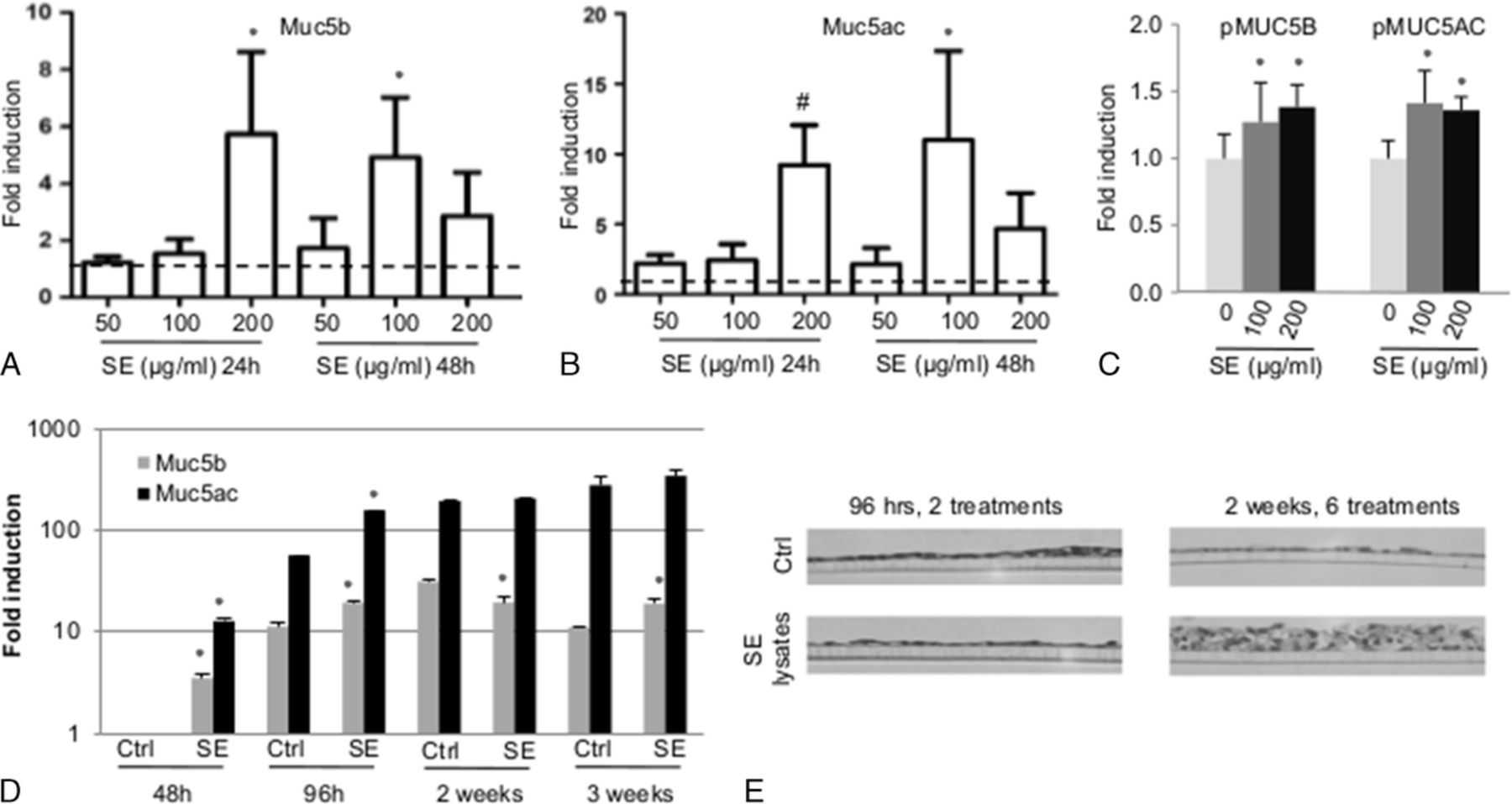
The S. epidermidis lysates induce mucin gene expression and promoter activation as well as epithelium thickening in mMEEC. A and B, The mMEECs were grown until 80% of confluence, deprived of serum 6 hours and treated for 24 to 48 hours with 50 to 200 μg/mL of S. epidermidis lysates. At the end of the treatment, the cells were lysed in Trizol, RNA was extracted, and an RT-qPCR was performed on MUC5B and MUC5AC genes with β-actin as the gene of reference. The dotted bar fixed at 1 is the control. C, The mMEECs were cultured until 50% to 60% of confluence and transfected with the different plasmid contructs containing MUC5B or MUC5AC promoter and the β-galactosidase as an internal control. After recovery and incubation in serum-free medium, the mMEECs were treated 16 hours with S. epidermidis lysates and lysed to perform a luciferase assay. D and E, The mMEECs were cultured at ALI during 3 weeks and were chronically treated 3 times a week with S. epidermidis lysates during 2 hours at the apical side of the epithelium in serum-free medium (48 hours = 1 treatment, 96 hours = 2 treatments, 1 week = 3 treatments, 2 weeks = 6 treatments, 3 weeks = 9 treatments). RNA was extracted and an RT-qPCR was performed on MUC5B and MUC5AC genes, with β-actin as the gene of reference (D). The cells were fixed, dehydrated, embedded, and then prepared for histologic analysis of transversal cuts of the epithelium (E) as well as observed with a 40× objective. The asterisk shows a statistical difference to the control with P < 0.05. The number sign shows a difference to the control with P = 0.0625.
Because we have found that mMEECs do not actively secrete mucin proteins (likely owing to a lack of goblet cells), we tested the human middle ear cell line, HMEEC-1, in a similar fashion to the conditions presented previously but also assayed for mucin protein production. At 96 hours, the human cells cultured at ALI demonstrated an increase in both MUC5B and MUC5AC expression after S. epidermidis exposure, with a fold induction of 2 for MUC5B and almost 5 for MUC5AC (all P < 0.05) (Fig. 2A). No induction was observed at the 48-hour exposure. This mRNA induction by S. epidermidis lysates was confirmed at the level of intracellular proteins because MUC5B proteins were slightly increased after the administration of 50 to 200 μg/mL of S. epidermidis lysates. (Fig. 2B). The induction of the proinflammatory cytokine IL-8 was also demonstrated with submerged and ALI cultures, with the effect starting earlier as the maximum-fold induction of this gene is reached at 24 hours and is sustained at 48 hours and 96 hours (Fig. 2, A and C).
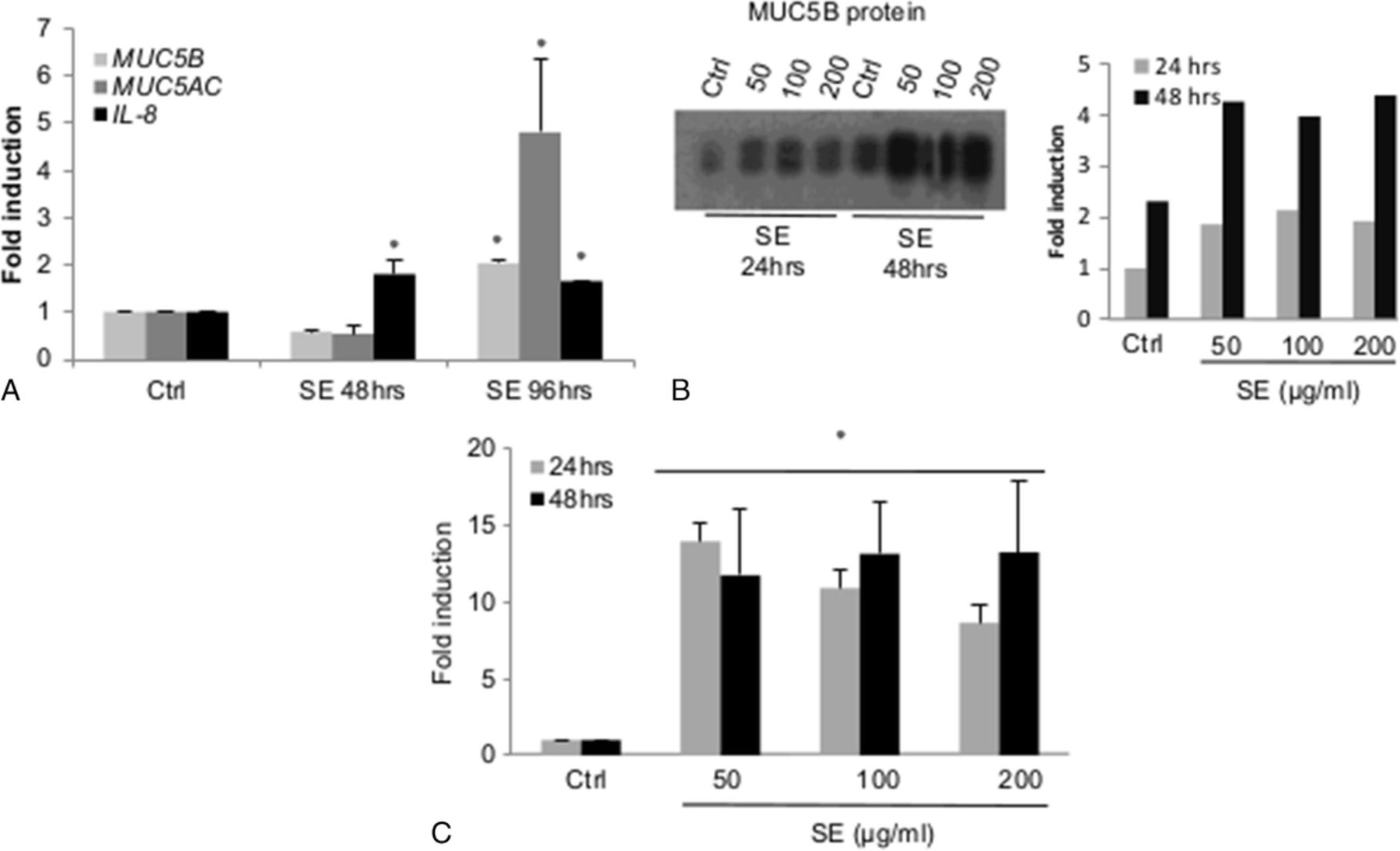
The S. epidermidis lysates induce mucin and IL-8 gene expression and protein production in HMEEC. The HMEECs were grown at ALI for 3 weeks (A) or in submerged conditions until 80% of confluence (B, C) and treated 24 hours to 48 hours with S. epidermidis lysates. At the end of the treatment, the cells were lysed in Trizol for RNA extraction and RT-qPCR analysis (A, C) or in RIPA buffer for MUC5B Western blot analysis (B). C, Robust IL-8 induction at 24 and 48 hours. The pixel count normalized to the control (performed using ImageJ software) is represented in the right graph. The asterisk shows a statistical difference to the control with P < 0.05.
S. Epidermidis Increases Nf-κB Activity in Mmeec
NF-κB is reported to be an important mediator of middle ear proinflammatory responses,20,29,30 and its activation has been shown with S. epidermidis stimulation in cultured nasal epithelial cells. 18 To elucidate whether S. epidermidis activates NF-κB in middle ear cells lines, we performed reporter assays, enzyme-linked immunosorbent assay (ELISA), Western blot, and immunostaining for NF-κB p65 subunit as 4 complementary methods to test for NF-κB activation. Reporter gene assays demonstrated a modest trend toward increases in activity of the pIgκB Luc plasmid relative to β-gal in mMEEC with 100 μg/mL of S. epidermidis, inducing statistically significant activation of the reporter plasmid (Fig. 3A). The ELISA demonstrated increased presence of p65 of NF-κB in nuclear extract of differentiated mMEEC at ALI stimulated with 200 μg/mL of S. epidermidis lysates, at the early 30-minute, 2-hour, and 6-hour time points compared with control, but not later, the 1-hour time point being not statistically significant because of the variability of the S. epidermidis–treated cells but still showing an increase of p65 in the nucleus of treated cells compared with the control condition (Fig. 3B). This was confirmed using the Western blot analysis of p65 in the cytoplasmic and nuclear fractions of mMEEC treated with 200 μg/mL of S. epidermidis lysates for 4 hours in submerged conditions (Fig. 3C). Nuclear fraction of the treated cells was shown to have more p65 than that of the control cells. This early-response pattern is typical for cell signaling resulting in NF-κB activation. This result correlated qualitatively with increased immunofluorescent presence of NF-κB p65 subunit localization into the nucleus of mMEEC (submerged) after 200-μg/mL lysate exposure at 30 minutes, with partial return to the cytoplasm at 6 hours (Fig. 3D). Taken together, the results shown in Figure 3 qualitatively and quantitatively demonstrate that S. epidermidis at 200 μg/mL causes early and notable nuclear translocation of NF-κB in mMEEC cultured in submerged conditions or differentiated at ALI.
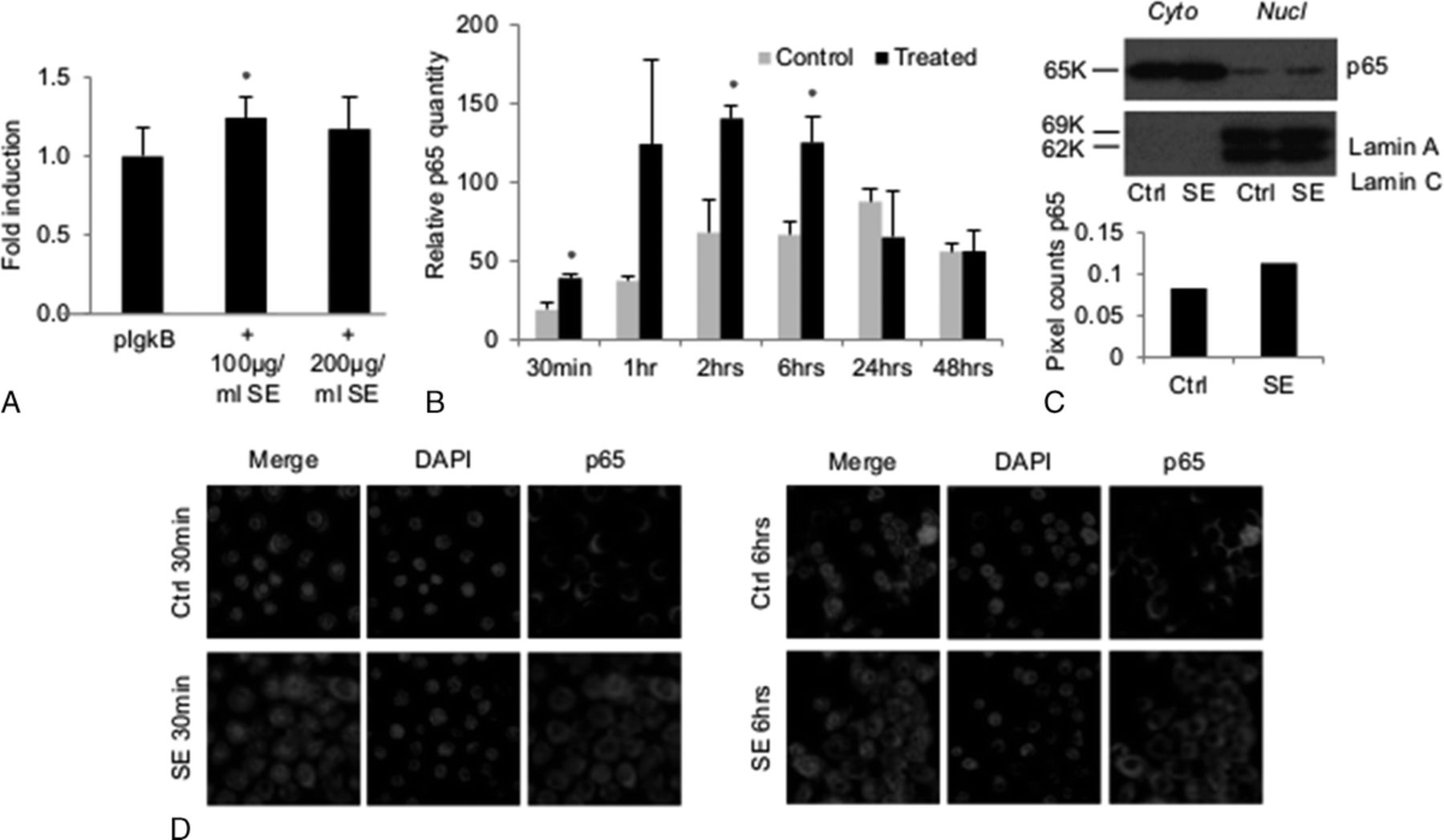
The S. epidermidis lysates induce increased NF-κB activity in mMEEC. A, The mMEECs were cultured until 50% to 60% of confluence and transfected with pIgκB and the β-galactosidase as an internal control. After recovery and incubation in serum-free medium, the mMEECs were treated 16 hours with S. epidermidis lysates and lysed to perform a luciferase assay (n = 18 biological replicates). The asterisk shows a statistical difference to the control with P < 0.05. B, The mMEECs were cultured at ALI during 3 weeks and were treated with S. epidermidis lysates at the apical side of the epithelium in serum-free medium for a maximum of 2 hours. The cells were then recovered and fractionated after 30 minutes to 48 hours after the beginning of the treatment, and an ELISA assay for p65 was performed on cytoplasmic and nuclear fractions. Nuclear fraction p65 content was normalized to the total quantity of proteins in each fraction as well as cytoplasmic fractions. Only the nuclear fraction data are represented because the cytoplasmic content was similar for control and treated at the same time point. C and D, The mMEECs were grown until 80% of confluence, deprived of serum and treated for 30 minutes and/or 4 hours with 200 μg/mL of S. epidermidis lysates. The cells were lysed for protein detection using the Western blot analysis (C) or fixed and stained for DNA (DAPI, blue) and p65 of NF-κB (AlexaFluor 548, red) for fluorescent microscopy analysis (D). The asterisk shows a statistical difference to the control with P < 0.05.
S. Epidermidis Lysates Induce a Proinflammatory and Oxidative Stress Response in Middle Ear Epithelial Cells
Upper respiratory epithelial cells are known to activate proinflammatory cytokines upon bacterial stimulation via TLR2 signaling induction of NF-κB. The typical pathogens known to elicit this response are H. influenzae and S. pneumoniae species; however, whether S. epidermidis can induce a similar response in the middle ear epithelium is completely unknown. For this reason, we measured the amount of mRNA signal for NF-kB–responsive and classic middle ear proinflammatory cytokines after S. epidermidis exposure.
The RT-PCR data showed a nonstatistically significant increase in Cxcl2 mRNA at 200 μg/mL of S. epidermidis lysates exposure for 48 hours in mMEEC with 4 independent experiments (each done in duplicate) (Fig. 4A), presumably owing to a variability of the response to the different batches of S. epidermidis lysates, as shown in Figure 1 for MUC5B and MUC5AC. Tnf-α cytokine showed a statistically significant increase in expression after 200 μg/mL of S. epidermidis lysates at 24 hours (P < 0.05) and an increase at 48 hours for the 100-μg/mL condition, which is not statistically different from the control (Fig. 4B). In addition, there were more modest, but statistically significant, increases in Tgf-α and Tgf-β at 48 hourrs but not IL-6 (Fig. 4C). Finally, the oxidative stress marker heme oxygenase 1 (Ho-1) (Fig. 4D) followed a similar transcriptional activation profile at 48 hours upon 200-μg/mL S. epidermidis stimulation at 48 hours. Taken together, these data indicate that, much like other classic upper respiratory invasive pathogens, the commensal S. epidermidis is also able to induce a proinflammatory chemokine response from the middle ear epithelium, in line with the noted NF-κB response effects.
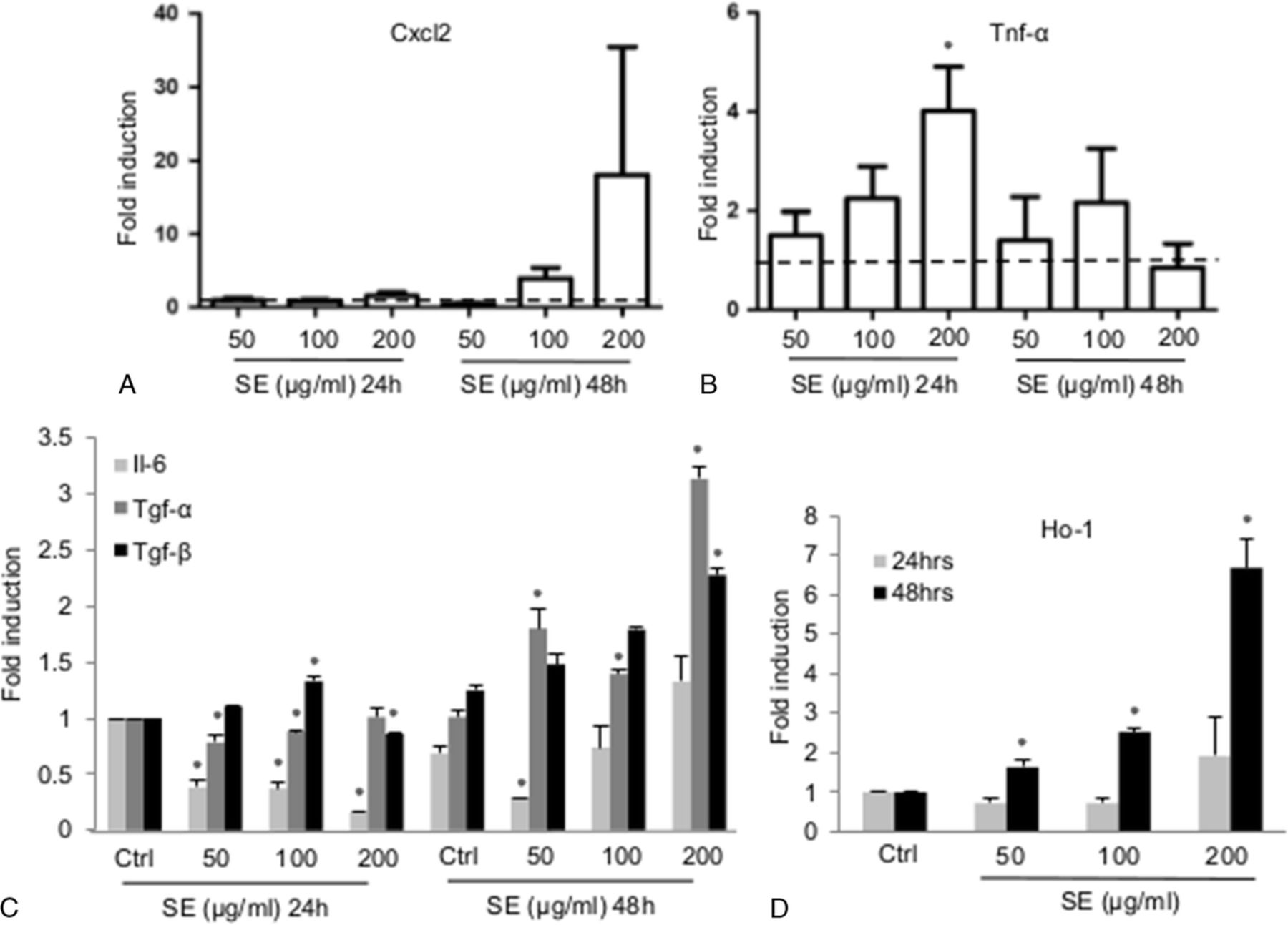
The S. epidermidis lysates induce proinflammatory cytokines expression and an oxidative stress marker in mMEEC. The mMEECs were grown until 80% of confluence, deprived of serum and treated for 24 to 48 hours with 50 to 200 μg/mL of S. epidermidis lysates. At the end of the treatment, the cells were lysed in Trizol, RNA was extracted, and an RT-qPCR was performed on Cxcl2 (A), Tnf-α (B), Il-6, Tgf-α, Tgf-β (C), and Ho-1 (D) genes, with β-actin as the gene of reference. The dotted line fixed at 1 is the control. The asterisk shows a statistical difference to the control with P < 0.05.
Proteomic Characterization of S. Epidermidis Lysates
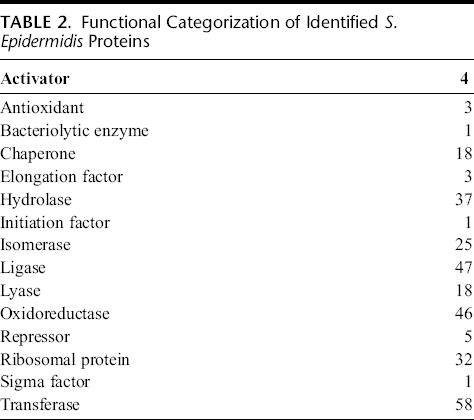
To interrogate potential mediators of proinflammatory responses, the global protein composition of 3 separate lysate batches was analyzed by LTQ-MS/MS proteomic techniques. Results identified 354 unique S. epidermidis proteins (Supplemental Table 1, Supplemental Digital Content 1, http://links.lww.com/JIM/A23). Of these, 33 were classified as membrane associated, 128 as cytoplasmic, and 11 as secreted proteins. Not surprisingly, the most common Uniprot functional classifications categories of these proteins correspond primarily to enzymatic activity, with transferase, ligase, oxyreductase, hydrolase, and isomerase occupying the top 5 functional categories (Table 2). The membrane-associated proteins (more likely to interact with the human epithelium) are listed in Supplemental Table 1 (Supplemental Digital Content 1, http://links.lww.com/JIM/A23). Importantly, very little is known or has been investigated or reported about the predicted human biological effects for a majority of these unique S. epidermidis proteins.
Functional Categorization of Identified S. Epidermidis Proteins
Discussion
It has been recognized that coagulase-negative staphylococcal species, most typically, S. epidermidis, are commonly cultured in chronic OME middle ear fluid specimens.10,31 Whether this organism is actually a contaminant or contributes to the disease has long been a matter of debate. 32 Because the potential role of biofilms in COME has been slowly elucidated and S. epidermidis has been identified as a common pathogen in COME epithelial and ossicular biofilms,11-14 we undertook initial studies into its potential effects on middle ear epithelium cellular responses.
Our data show a direct link between lysates from S. epidermidis bacteria and increased genetic transcription of the secreted airway mucin glycoproteins MUC5AC and MUC5B in both mouse and human middle ear epithelial cell lines. This finding is important because, to date, there is little information known as to how pathogens that can hide from the immune system within the context of a biofilm matrix may be able to contribute to chronic mucoid middle ear fluid secretion. Our results suggest that it is possible that bacterial lysates chronically shed or released from such bacterial reservoirs could directly contribute to the overproduction of mucins in the middle ear. Further, our results demonstrate that chronic time point stimulation of the mouse middle ear epithelium with S. epidermidis lysates also induced marked epithelial layer thickening in vitro, akin to the known middle ear hyperplasia frequently seen in COME biopsy specimens.33,34 This suggests that S. epidermidis could play a role in OM much because it has been widely demonstrated in the literature for NTHi or S. pneumoniae, known for their reactivity on middle ear epithelial cells in vitro. Lysates of NTHi and S. pneumoniae have been shown to increase MUC5AC mRNA and promoter activation in HMEEC.35-38 However, despite the predominance of MUC5B in middle ear effusions,15,16 no in vitro study has been conducted on analyzing the regulation of MUC5B in response to bacterial component exposure. Furthermore, it is worthy to note that, in the mMEEC used in this report, as opposed to our findings with S. epidermidis, we have not seen NTHi directly induce MUC5B mRNA. 19 Indeed, our study is the first one showing MUC5B upregulation by lysates of a bacterium, specifically the commensal organism S. epidermidis.
Data from this report also identified that lysates from S. epidermidis induced a proinflammatory cellular response in mMEEC, with marked upregulation of Cxcl2/IL-8 and TNF-α, 2 cytokines that have been shown to be in high concentrations in middle ear effusions from patients having chronic OM. 39 IL-6, TGF-α, and TGF-β have also been found in middle ear effusions and implicated in in vitro and/or in vivo studies,35,39,40 but only TGF-α and TGF-β were increased in our study. This suggests that mouse epithelial cells mMEEC are able to produce certain cytokines in response to the bacteria lysates, whereas the presence of other cell types, such as immune cells in in vivo patient samples, likely accounts for the production of other observed cytokines. These inductions of proinflammatory mediators may imply pathogenic potential of S. epidermidis to initiate and/or sustain inflammatory responses in middle ear cells and argue against their role as “nonpathogenic contaminants” in the ear. A similar response to these strains has been observed in vitro with nasal-derived epithelial cells 18 and human bronchial epithelium. 41 IL-8 and its murine homologue Cxcl2 have been recognized by us and others as a critical mediator of bacterial-induced inflammation in the airways and in the ear, where it may contribute to mucin hypersecretion,19,42 which seems to be relevant with the induction of IL-8 expression in HMEEC as early as 24 hours after S. epidermidis, whereas MUC5B and MUC5AC mRNAs are induced at 48 hours. This link between cytokine release and mucin production is currently under investigation in our laboratory. The antioxidant enzyme Ho-1 was also shown to be induced in response to S. epidermidis lysates. This enzyme is regulated by NF-κB transcription factor and oxidative stress mediators because the promoter of this gene also contains antioxidant-responsive elements.43,44 Heme oxygenase 1 constitutes a biomarker of oxidative stress that is demonstrated to be induced at 48 hours of S. epidermidis lysate exposure, suggesting that oxidative stress could be implicated in S. epidermidis effects on middle ear epithelial cells. This is supported by the study of Sprenger et al. 45 that showed an antioxidant-driven decrease in MUC5AC mRNA induction due to bacterial stimulation.
Given that these mediators are known to be transcriptionally responsive to the transcription factor NF-κB and the fact that NF-κB has also been purported to be mediator of the chronic inflammatory response seen in OM,24,29 we sought to investigate whether S. epidermidis lysates would be able to activate NF-κB in mMEEC as a potential mechanism for the noted cytokine and mucin gene upregulation. Indeed, our data demonstrate early translocation of the activated NF-κB subunit, p65, into the cell nucleus both qualitatively and quantitatively. Data of IF showed intranuclear localization as early as 30 minutes, with some sustainment up to 6 hours relative to control stimulation. These findings were confirmed with Western blot and ELISA data. Whether this activation is directly responsible for the subsequent increases in mucin and cytokine secretion at 24 and 48 hours is the subject of ongoing investigations in our laboratory. At this point, we can only comment on the correlation of both events occurring in a timed fashion (with the NF-κB induction occurring early and the mucin and cytokine induction happening later). The presence of binding sites of NF-κB in the promoters of MUC5B and MUC5AC genes strongly suggests NF-κB implication in the regulation of these genes in response to S. epidermidis lysates exposure. Other investigators have found that NTHi lysates also activate NF-κB pathway by both translocation dependent and independent mechanisms in HMEEC, suggesting that other factors should be considered in the regulation of this pathway. 46 Indeed, we noted that S. epidermidis induces only a mild, but clear, effect on early NF-kB nuclear translocation. Further, in separate experiments, when we have used live S. epidermidis bacteria to stimulate these cells and have assayed for NF-κB nuclear translocation with IF, we have not noticed the same type of p65 activation as with the bacterial lysate preparations (Supplemental Fig. 1, Supplemental Digital Content 2, http://links.lww.com/JIM/A24), which describes that live S. epidermidis does not induce NF-κB translocation to the nucleus. The mMEECs were grown until 80% of confluence, deprived of serum and antibiotics 6 hours and treated for 6 hours to 24 hours with live S. epidermidis [106 to 2 × 106 of colony-forming units]. The cells were fixed and stained for DNA (DAPI, blue) and p65 of NF-κB (AlexaFluor 548, red) for fluorescent microscopy analysis. Further work exploring the comparative effects of live versus lysed S. epidermidis is ongoing in our laboratory, but initial results seem to indicate that the lysates are much more proinflammatory, perhaps suggesting that live bacteria induce less biological responses than when they are lysed, implicating intracellular bacterial components in biological responses of the middle ear epithelial cells.
Finally, in an effort to identify novel S. epidermidis–derived proteins capable of inducing proinflammatory responses, the global protein composition of 3 separate lysate batches was subjected to LTQ-MS/MS proteomic analysis. Findings were able to identify a comprehensive list of unique proteins present in 3 separate preparations of these lysate batches. Most of the proteins were found in all the batches of lysate, but some were present in 1 or 2 of the batches. This variability, also considering the probable difference in quantity of proteins (not assayed by label-free proteomics), induces different ranges of biological responses in mMEEC. The majority of the proteins found in the 3 batches of S. epidermidis analyzed were classified as enzymatic, proteases, and ligases corresponding to a big proportion (indirectly represented by peptide counts). Although little work has been carried out to investigate the unique biological effects of these mediators, serine proteases, much like those found in our lysate batches, have been purported to induce IL-8 secretion via NF-κB activation. 18 In addition, we identified the known S. epidermidis virulence factors autolysin, elastin-binding protein, and accumulation-associated protein. Each of these factors has been postulated to contribute to potential biofilm formation. 47 Nonetheless, a more extensive analysis of these proteins to determine their capability of activating host inflammation seems warranted.
Conclusions
In conclusion, our data have demonstrated that S. epidermidis lysates activate a potent and robust proinflammatory response in MEEC, including mucin gene expression and mucin protein production as well as the activation of the transcription factor NF-κB. Although typically considered a nonpathogenic commensal organism in the ear, these results suggest that they may play a role in the perpetuation of an inflammatory and monogenic response in COME, especially considering that these bacteria are known to evade host immunity by “hiding” in biofilm matrices.
Footnotes
Acknowledgments
The authors thank Dr. Rose and Dr. Colberg-Poley of the Center of Genetic Medicine for their kind advice in this work. The authors also thank Dr. Kristy Broan of the Proteomics Core for her guidance with the proteomic experiments.