
Abstract
Background
Peroxisome proliferator-activated receptor γ (PPARγ) and Wnt play different roles in bone homeostasis. Thiazolidinediones are PPARγ agonists that cause bone mineral density loss. This study investigated the relationship between PPARγ and Wnt/β-catenin signaling in mouse osteoblastic MC3T3-E1 cells.
Methods
MC3T3-E1 cells were treated with either pioglitazone (Pio) or rosiglitazone (Rosi), thiazolidinediones, for 24 hours at 10 to 40-μM concentrations. Recombinant mouse Wnt3a protein (50 ng/mL) for 6 hours was also used to treat the 20-μM Pio and Rosi pretreated cells. Cell proliferation was measured by MTT, and apoptosis with flow cytometry using annexin V/propidium iodide staining; reverse transcriptase–polymerase chain reaction measured mRNA expression levels of LRP5/6 (low-density lipoprotein–related protein 5/6), glycogen synthase kinase 3β (GSK3β), TCF7L2 (transcription factor 7–like 2), PPARγ, and cyclin D1, and Western blots detected β-catenin and p-GSK3β proteins.
Results
Pioglitazone and Rosi decreased MC3T3-E1 cell viability by 28.07% and 18.14% at 20 μM, respectively (P < 0.05). Apoptosis increased compared with controls (7.21%), after 20-μM treatment with Pio or Rosi, to 10.45% and 12.10%, respectively (P < 0.05). Both Pio and Rosi decreased β-catenin protein levels and increased p-GSK3β, but the LRP5/6, GSK3β, and TCF7L2 mRNA levels were constant. Upon activation of the Wnt pathway by mouse Wnt3a protein, β-catenin and p-GSK3β protein levels were reversed, accompanied with increased proliferation, but apoptosis remained high.
Conclusions
Activation of PPARγ in osteoblasts accompanied Wnt signaling suppression. Activation of Wnt signaling alleviated the PPARγ proliferation decreases but not the apoptosis increases. The thiazolidinedione PPARγ agonists act in part through inhibition of the Wnt signaling pathway, showing there is a relationship between PPARγ and Wnt signaling.
Peroxisome proliferator-activated receptor γ (PPARγ), a member of the nuclear receptor family, is expressed in many tissues and involved in the regulation of genes involved in lipid metabolism, adipogenesis, insulin sensitivity, the immune response, growth, and differentiation. 1 Peroxisome proliferator-activated receptor γ has been shown to regulate glucose metabolism, bone mass, and energy expenditure.2,3 Activation of PPARγ requires heterodimerization with another nuclear receptor, retinoid X receptor, and binding of specific ligands including polyunsaturated fatty acids, oxidized metabolites of linoleic acid 9-hydroxy-octadecadienoic acids and 13-hydroxy-octadecadienoic acids, 15-deoxy-(12,14)-prostaglandin J2, certain phospholipids, and antidiabetic drugs such as thiazolidinediones (TZDs). 4 In clinical practice, 2 TZDs, rosiglitazone (Rosi) and pioglitazone (Pio), are widely used in the management of type 2 diabetes mellitus (T2DM) as a result of evidence that treatment with Rosi increases the duration of satisfactory glycemic control in T2DM compared with metformin and glyburide 5 and slows the rate of progression from impaired glucose tolerance to T2DM. 6
An essential role of PPARγ is in bone homeostasis. Peroxisome proliferator-activated receptor γ is expressed in bone marrow stromal cells, 7 osteoblasts, 8 and osteoclast precursors. 9 Within the bone microenvironment, activation of PPARγ signaling promotes differentiation of mesenchymal stem cells into adipocytes at the expense of osteoblasts, suppressing the expression of early osteoblast genetic markers such as cbfa1/runx2, and inhibiting the expression of an osteoblastic phenotype. 10 In cells of a mesenchymal lineage, TZD-activated PPARγ induces adipocyte and suppresses osteoblast differentiation. 11 In cells of a hematopoietic lineage, the activation of PPARγ positively regulates differentiation of osteoclasts. 12 Peroxisome proliferator-activated receptor γ decreases production of insulinlike growth factor 1 by stromal cells 13 and promotes differentiation of osteoclast precursors into mature bone-resorbing osteoclasts. The net effect is to decrease bone formation and increase bone resorption. In heterozygous PPARγ-deficient or hypomorphic mutation of PPARγ gene locus mice, decrease in PPARγ activity led to increased bone mass due to increased osteoblast number. 14 Administration of Rosi to mice and rats results in significant decreases in bone mineral density (BMD) and bone volume and changes in bone microarchitecture.2,15,16 In humans, therapeutic modulation by TZDs elicits unwanted effects on bone, which result in progressive bone loss and diminished levels of circulating bone formation markers in women, especially older postmenopausal women,17-19 and Rosi apparently increases the frequency of fractures in women.5,20,21 In addition, clinical evaluation of PPARγ gene polymorphisms in different human populations strongly suggests a role in the regulation of bone mass; several novel polymorphisms were also identified in the coding region of PPARγ that, particularly in women, correlated with BMD independently of body weight and T2DM.22-24 Activation of PPARγ signaling at extraskeletal sites, adipose tissue, and the liver and the ensuing effects of insulin sensitization on the β cells of the pancreas may indirectly impact upon bones by altering the circulation levels of several hormones and cytokines, such as leptin, adiponectin, and amylin. 25 Activation of PPARγ in osteoblastic cells promotes apoptosis 26 ; low concentrations of PPARγ agonist (5 μM) enhance osteoblastic differentiation, whereas a relatively high concentration of PPAR activator or PPARγ overexpression inhibits osteoblastic differentiation. 27
Wnts are secreted glycoproteins that play an important role in development and maintenance of many organs and tissues, including bone. 28 Wnt proteins act through several pathways to regulate cell growth, differentiation, function, and death. In bone, the Wnt/β-catenin or canonical pathway appears to be particularly important, 29 as it regulates bone development and accrual through different mechanisms at different stages of life. Wnt/β-catenin is active in all cells of an osteoblastic lineage and involves the regulation of multiple transcription factors and the stabilization of β-catenin. Activation of canonical Wnt-signaling promotes osteoblast differentiation from mesenchymal progenitors at the expense of adipogenesis, which leads to improved bone strength, whereas suppression of the pathway causes bone loss. 30 Wnt/β-catenin signaling promotes osteoblast differentiation, proliferation, and mineralization activity while blocking osteoblast apoptosis.31,32 As a key factor in the canonical Wnt pathway, β-catenin plays crucial role in both early and late stage of osteoblastogenesis. Mice with a targeted deletion of β-catenin in mesenchymal precursors of chondrogenesis and osteogenesis show a reduction in all relevant markers of osteogenesis and an absence of both endochondral and intramembranous bone. 33 In fact, the absence of β-catenin in the early stages of osteoblastogenesis steers the fate of mesenchymal precursors toward chondrogenesis. 34 During the later stage of osteoblast development, constitutively active β-catenin causes bone resorption and osteoclastogenesis. 35
Despite their different actions in bone metabolism, there is significant cross-regulation between PPARγ and Wnt/β-catenin signaling. Activation of PPARγ by exposure of Swiss mouse fibroblasts to troglitazone stimulates the degradation of β-catenin, dependent on glycogen synthase kinase (GSK) 3β activity. 36 Thiazolidinedione-mediated activation of PPARγ inhibits growth through an inhibition of β-catenin expression at the posttranslational level. 37 In adipose tissue, the PPARγ pathway functions via antagonism of the β-catenin pathway. On the other hand, sustained activation of β-catenin in preadipocytes blocks adipogenesis by inhibiting PPARγ-associated gene expression. 38 In order to explore whether the activation of PPARγ affects bone tissue in a Wnt/β-catenin-related manner, our study used osteoblastic MC3T3-E1 cells to research the influence on cell proliferation and molecular expression levels by Pio and Rosi, with particular attention to the cross-regulation between these 2 pathways.
Materials and Methods
Cell Culture
Mouse osteoblastic MC3T3-E1 cells were purchased from American Type Culture Collection and cultured in α-minimal essential medium (HyClone; GE, Logan, UT) with ribonucleosides and deoxyribonucleosides but without ascorbic acid with 2 mM L-glutamine and 1 mM sodium pyruvate that contained 10% fetal bovine serum (HyClone; GE) and 100 U/mL penicillin/streptomycin. Cells were maintained in an incubator at 37 °C in 5% CO2. The study was approved by the ethics committee of Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, China.
Cell Proliferation Assay
MC3T3-E1 cells were seeded into 96-well plates at a density of 1 × 105 cells/mL (100 μL/well) in complete culture medium and then treated with different concentrations (0, 10, 20, 30, and 40 μM) of PPARγ agonists Pio (Jiangsu Hengrui Medicine Co, Ltd, Jiangsu, China) and Rosi (Sigma, St. Louis, MO) or recombinant mouse Wnt3a protein (R&D Systems, Shanghai, China) (50 ng/mL) for 24 hours, then 5 mg/mL MTT solution (Beyotime, Shanghai, China) 10 μL was added to each well for 4 hours. After careful removal of culturemedia, 100 μL dimethyl sulfoxidewas added to fully dissolve the purple crystals by shaking for 10 minutes. Absorbance at 492 nm was determined using a microplate reader (Bio-Rad, Hercules, CA). The cell viability percentage was calculated according to the following formula:
Cell viability (%) = (average absorbance (A) of the experimental group/average absorbance (A) of the control group) × 100%.
Cell Apoptosis Detected by Annexin V/Propidium Iodide Staining
To evaluate the apoptotic activity, annexin V staining was performed using the annexin V-FITC Apoptosis Detection Kit (Keygen, Nanjing, China) in accordance with the manufacturer's instructions. MC3T3-E1 cells were cultured in a 6 well plate at a density of 2x105/well, then 20 μM Pio and Rosi, as well as recombinant mouse Wnt3a protein (50 ng/mL) were added for 24 hours. Apoptosis was determined by flow cytometric analysis using a FACScan flow cytometer (BD Biosciences, San Jose, CA, USA). annexin V was set as the horizontal axis and propidium iodide (PI) was set for the vertical axis. Mechanically damaged cells were located in the upper left quadrant, apoptotic or necrotic cells in the upper right quadrant, dual-negative and normal cells in the lower left quadrant, and early apoptotic cells in the lower right quadrant of the flow cytometric dot plot.
Semiquantitative Reverse Transcriptase–Polymerase Chain Reaction
Total RNA was extracted from MC3T3-E1 cells using the Trizol reagent (Invitrogen, Carlsbad, CA), according to the manufacturer's instructions. The purity of RNA was identified in a UV spectrophotometer by diluting 2 μL RNA in 198 μL DEPC water and measuring the OD260 and OD280. A ratio of OD260/OD280 between 1.8 and 2.0 was of the required purity. The RNA was then transcribed to cDNA using the First Strand cDNA Synthesis Kit (Fermentas; Thermo Fisher, Waltham, MA). Glycogen synthase kinase 3β, low-density lipoprotein–related protein 5 (LRP5), transcription factor 7–like 2 (TCF7L2), PPARγ, cyclin D1, and β-actin primers (Table 1) were synthesized by Sangon (Shanghai, China). Polymerase chain reaction (PCR) conditions are shown in Table1. The corresponding PCR products were analyzed by 1.5% agarose gel electrophoresis. The gels were analyzed by a BioRad imaging system with Quantity One analysis software (Hercules, CA). Gray values were obtained, and relative mRNA expressions were normalized to β-actin.
Primer Sequences for PCR and PCR Reaction Conditions
Western Blot Analysis
For Western blot analysis, total protein was extracted from MC3T3-E1 cells, using the cell lysis buffer for Western blot from Beyotime. Protein content was determined by Coomassie brilliant blue. Equal amounts of protein (50 μg) were separated by 8% sodium dodecyl sulphate-polyacrylamide gel electrophoresis, transferred to polyvinylidene fluoride membranes (Millipore, Darmstadt, Germany), blocked with Tris-buffered saline and Tween 20–confining liquid containing 20% nonfat milk for 2 hours at room temperature, and then probed with the respective primary antibodies (mouse β-catenin antibody [BD Bioscience Pharmingen, Danvers, MA], rabbit p-GSK3β antibody [Cell Signaling Technology, Danvers, MA], tubulin antibody [Beyotime, Jiangsu, China]) overnight. Horseradish peroxidase–conjugated secondary antibodies for 2 hours (goat anti–rabbit IgG [Fly Implement Technology, Shanghai, China] and goat anti–mouse IgG [Invitrogen]) and the ECL detection kit (BeyoECL Plus, Shanghai, China) were used for the detection of specific proteins. All images were taken by a Bio-Rad imaging system with Quantity One analysis software. Relative protein expressions were normalized to the tubulin expression level.
Statistical Analyses
All statistical analyses were conducted using SPSS 18.0 software package (SPSS Inc, Chicago, IL). All data are presented as means ± SD from at least 3 independent experiments. Statistical significance was evaluated by t test or 1-way analysis of variance with least significant difference test for post hoc analysis. P < 0.05 was considered to be statistically significant.
Results
Pio and Rosi Inhibited MC3T3-E1 Cell Proliferation in Part through the Wnt Signaling Pathway
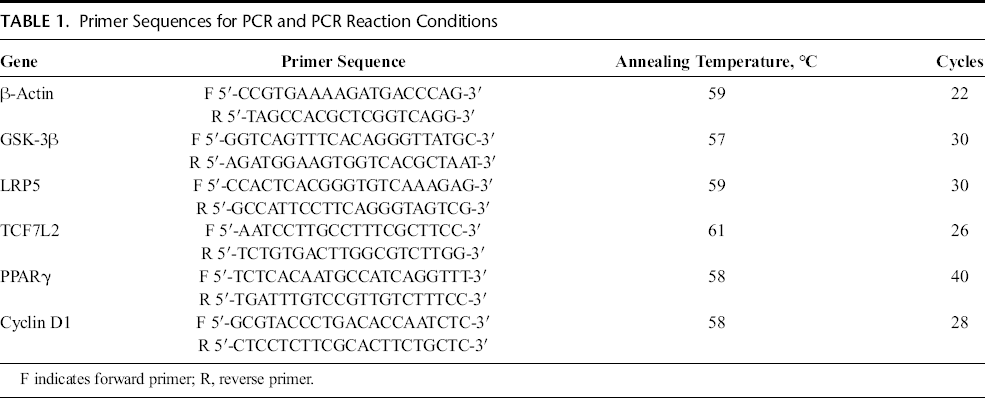
Peroxisome proliferator-activated receptor γ agonists Pio and Rosi both reduced the proliferation of MC3T3-E1 cells. The TZDs showed different influences on cell proliferation with increasing concentration. Compared with the control group, Pio reduced proliferation by 100%, 88.33% ± 2.71%, 71.93% ± 2.13%, 71.05% ± 3.01%, and 64.93% ± 3.99% at 0, 10, 20, 30, and 40 μM, respectively. At 20 μM, Pio reduced proliferation significantly (P < 0.05), but there was no further significant reduction between 20 and 40 μM. In the case of Rosi, cell activity was 100%, 90.62% ± 2.97%, 81.86% ± 3.48%, 74.62% ± 2.93%, and 73.52% ± 3.34% with progressively increased concentrations, respectively. The relative cell proliferation activity showed a dose-dependent decline from 10 to 30 μM, which was significant at 20 μM (P < 0.05) but no further significant decline between 30 and 40 μM. Pioglitazone showed more intensive reduction in cell proliferation than did Rosi (Fig. 1A).
Pioglitazone and Rosi inhibited MC3T3-E1 cell proliferation through the Wnt signaling pathway. Cell proliferation was determined by MTT. A, MC3T3-E1 cells were treated with different concentrations (0, 10, 20, 30, and 40 μM) of PPARγ agonists Pio or Rosi for 24 hours. *P < 0.05 versus control group (0 μM Pio or Rosi). B, MC3T3-E1 cells were pretreated with 500 ng/mL DKK1 (antagonist of Wnt signaling) for 1 hour and then treated with 50 ng/mL recombinant mouse Wnt3a protein (Wnt3a) for 24 hours. *P < 0.05 versus control group (untreated cells); #P < 0.05 versus Wnt3a group. C, MC3T3-E1 cells were pretreated with 20 μM Pio or Rosi for 24 hours and then treated with 50 ng/mL recombinant mouse Wnt3a protein for another 6 hours. *P < 0.05 versus control group (untreated cells); #P < 0.05 versus Pio or Rosi; ΔP < 0.05 versus Wnt3a. Percentages of cell viability are shown as mean ± SD.
Meanwhile, a significant increase in cell proliferation was observed in cells treated with recombinant mouse Wnt3a protein (50 ng/mL), a protein that specifically activates the Wnt/β-catenin signaling pathway, for 6 hours as compared with the control (124.87%) (P < 0.05). In contrast, adding DKK1 (500 ng/mL) (R&D Systems) an hour before Wnt3a protein, to block Wnt/β-catenin signaling, decreased proliferation to the control level (P > 0.05) (Fig. 1B). These results demonstrated that the activation of Wnt/β-catenin signaling by recombinant mouse Wnt3a protein enhanced the proliferation of osteoblasts.
In order to explore whether the negative effects of TZDs on osteoblast proliferation act in relation to Wnt signaling, cells treated with Pio and Rosi for 24 hours were then treated with Wnt3a protein sequentially for another 6 hours. The results showed that Wnt3a protein in part increased cell proliferation in both Pio and Rosi groups (Fig. 1C). So Wnt3a protein ameliorated the reduction in osteoblast proliferation caused by the PPARγ agonists Pio and Rosi.
Pio and Rosi Induced MC3T3-E1 Cell Apoptosis but Not through the Wnt Signaling Pathway
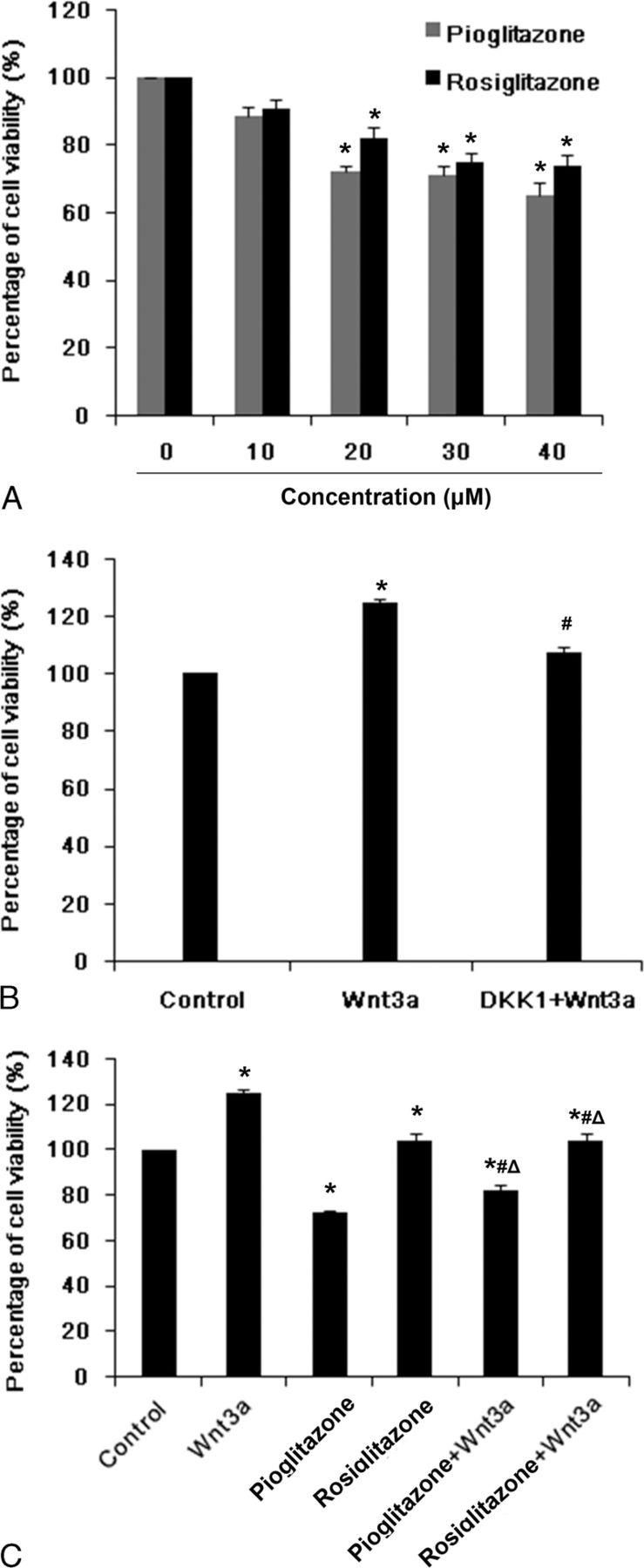
Compared with the control group (7.21%), the apoptosis rate of MC3T3-E1 cells treated with Pio or Rosi for 24 hours increased significantly, by 10.45% or 12.10%, respectively, but no significant difference was observed between the Pio and Rosi groups (Fig. 2). Intervention with Wnt3a protein alone decreased the cell apoptosis rate (5.13%) compared with the control group, but the apoptosis rate was not significantly changed after Wnt3a protein treatment in cells pretreated with Pio or Rosi; this suggests that Wnt signaling may not mediate the apoptosis caused by PPARγ agonists (Fig. 2).
Pioglitazone and Rosi induced MC3T3-E1 cell apoptosis not through the Wnt signaling pathway. MC3T3-E1 cells were pretreated with 20 μM Pio or Rosi for 24 hours and then treated with 50 ng/mL recombinant mouse Wnt3a protein for another 6 hours. Cell apoptosis were determined by flow cytometric analysis using annexin V/PI staining. Total apoptotic cells (% of total cells) are shown as mean ± SD. *P < 0.05 versus control group (untreated cells); #P < 0.05 versus Pio or Rosi; ΔP < 0.05 versus Wnt3a.
Pparγ Revealed No Significant Regulation of the Major Factors of Wnt/β-Catenin Signaling Pathway at the Mrna Levels
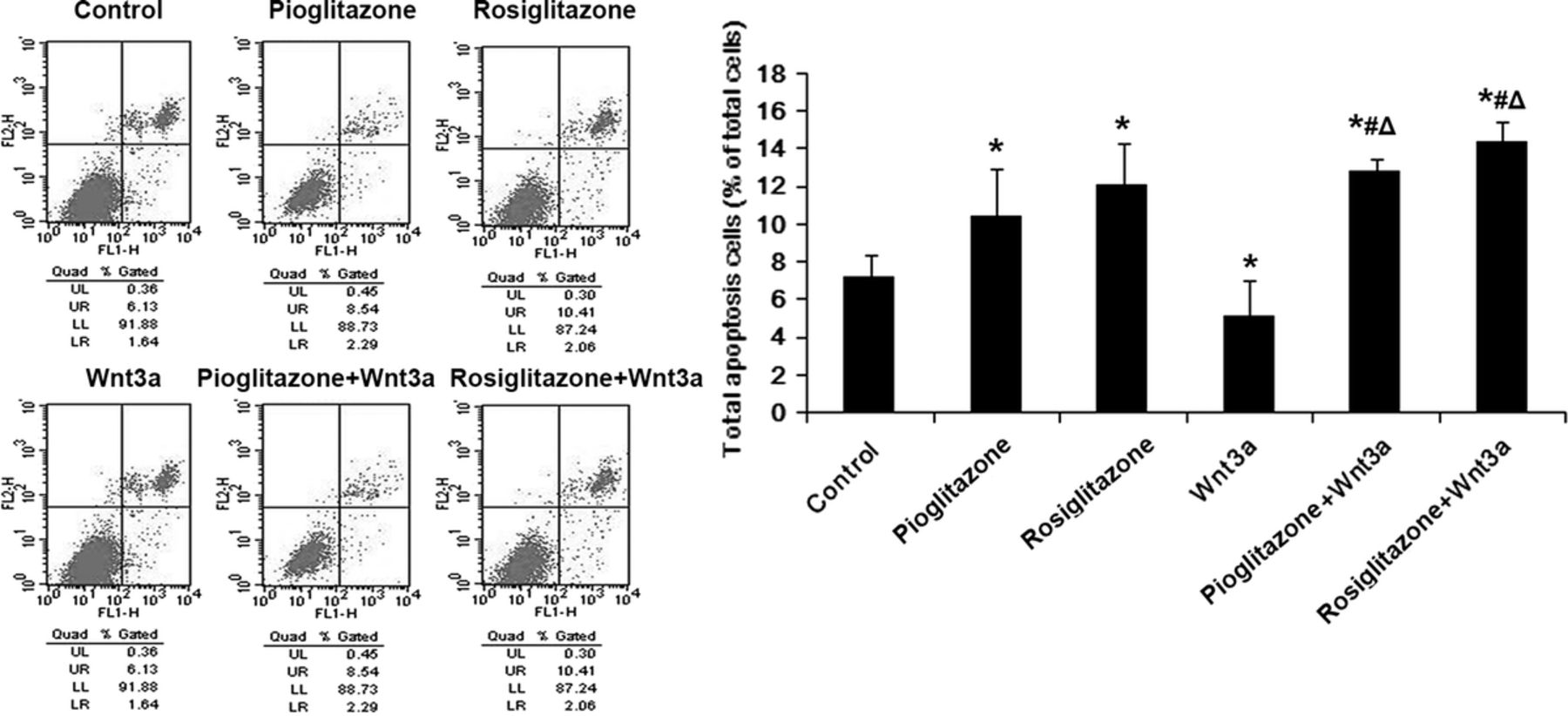
We used semiquantitative reverse transcriptase (RT)–PCR to detect any alteration in the levels of the major factors of the Wnt pathway and PPARγ. Compared with the control group, neither Pio and Rosi nor Wnt3a protein regulated the mRNA expression levels of the major factors of the Wnt/β-catenin pathway, including LRP5/6, GSK3β, and TCF7/L2. The levels of cyclin D1 mRNA were down-regulated by the TZDs and up-regulated by Wnt3a protein (Fig. 3). Meanwhile, both Pio and Rosi at 20 μM increased PPARγ mRNA levels compared with the controls (P < 0.05), and Wnt3a protein significantly decreased (P < 0.05). These results demonstrated that Wnt signaling impacted upon PPARγ transcription, even after intervention with Pio and Rosi (Fig. 3).
Peroxisome proliferator-activated receptor γ revealed no significant regulation of the major factors of the Wnt/β-catenin signaling pathway on mRNA levels. MC3T3-E1 cells were pretreated with 20 μM Pio or Rosi for 24 hours and then treated with 50 ng/mL recombinant mouse Wnt3a protein for another 6 hours. mRNA expression was determined by RT-PCR. β-Actin was used as an internal reference. Relative mRNA expression is shown as mean ± SD. *P < 0.05 versus control group (untreated cells); #P < 0.05 versus Pio or Rosi; ΔP < 0.05 versus Wnt3a.
Pparγ Induced a Decrease in β-Catenin Protein Expression in Osteoblasts, and the Decrease Required P-Gsk3β Enhancement
The studies above have demonstrated that activation of PPARγ induced a decline in cell proliferation, whereas activating Wnt/β-catenin signaling ameliorated this influence. In an attempt to understand the mechanism regulating the process and the relationship between PPARγ and the Wnt pathway, we used Western blots to detect the β-catenin and p-GSK3β expression in MC3T3-E1 cells after TZDs intervention for 24 hours. Compared with the control group, Pio and Rosi at 20 μM both down-regulated the levels of β-catenin protein (P < 0.05), but there was no significant difference between the 2. At the same time, the level of p-GSK3β was up-regulated (P < 0.05); no significant difference could be found between the groups (Fig. 4A).
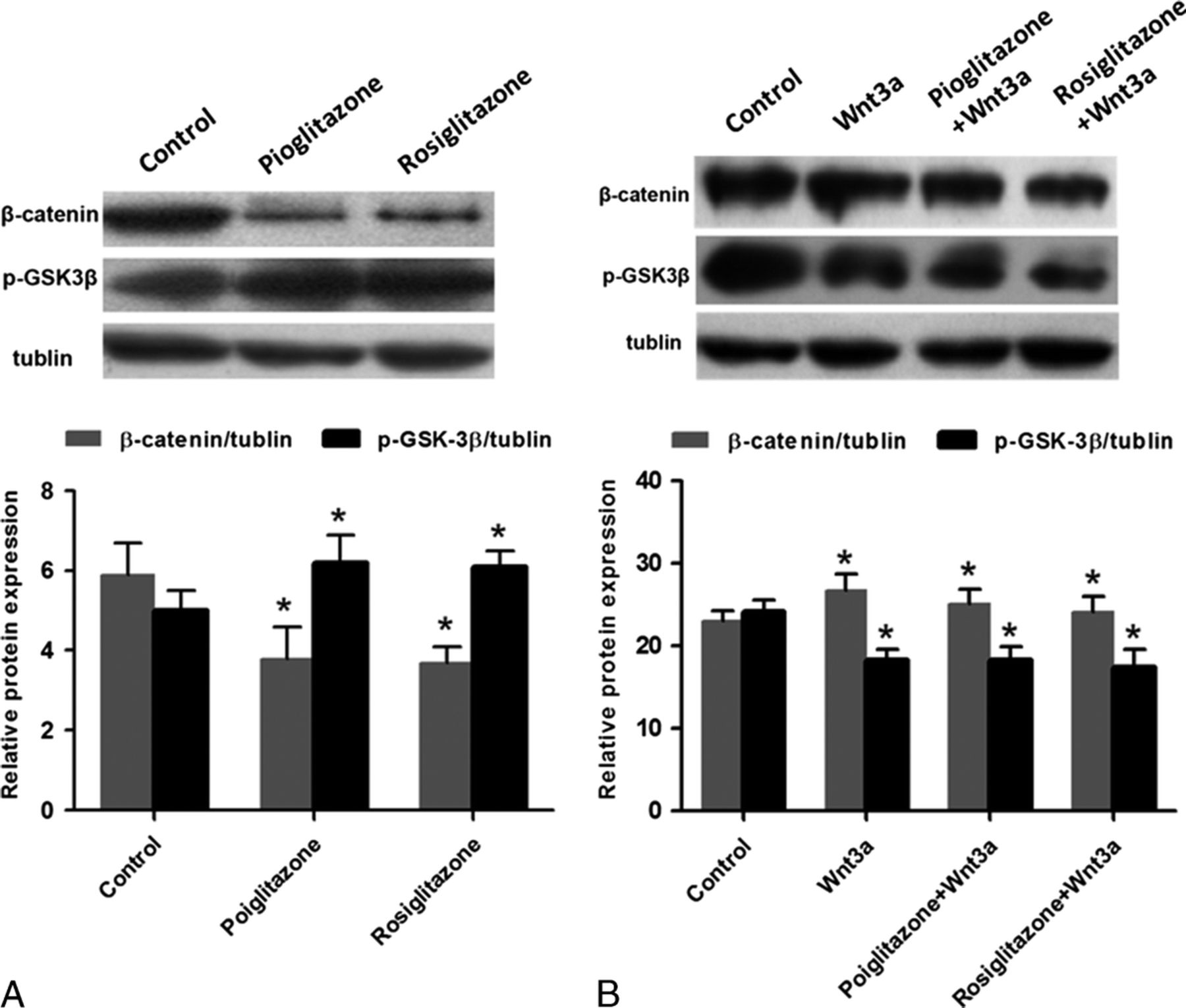
Pioglitazone and Rosi regulated the protein expression levels of β-catenin and p-GSK3β through the Wnt signaling pathway in MC3T3-E1 cells. Protein expression was determined by Western blot. Tubulin was used as an internal reference. A, MC3T3-E1 cells were treated with 20 μM Pio and Rosi for 24 hours. *P < 0.05 versus control group (untreated cells). B, MC3T3-E1 cells were pretreated with 20 μM Pio and Rosi for 24 hours and then treated with 50 ng/mL recombination mouse Wnt3a protein for another 6 hours. *P < 0.05 versus control group.
Recombinant mouse Wnt3a protein activated Wnt/β-catenin signaling by increasing β-catenin protein levels and decreasing p-GSK3β levels, which can be seen in Figure 4B (P < 0.05). To further test whether PPARγ induced declined proliferation through a Wnt-related pathway, we added Wnt3a protein (50 ng/mL) to activate Wnt signaling after the activation of PPARγ by TZDs; as a result, the β-catenin expression levels were positively regulated and p-GSK3β level negatively regulated (Fig. 4B), and cell proliferation increased as observed above.
Discussion
The aim of this study was to investigate PPARγ activity in osteoblasts and the relationship between its action and the Wnt signaling pathway, to understand more clearly how PPARγ agonists cause BMD loss. We found that there was a complex relationship between PPARγ and Wnt in relation to osteoblast activity.
The PPARγ agonists Pio and Rosi at 20 μM significantly reduced the proliferation of osteoblast cells. Rosiglitazone provided a dose-dependent response over 10 to 30 μM, but a similar effect was found for Pio over just 10 to 20 μM, although Pio provided a statistically significantly greater decrease than Rosi at 20 μM. Osteoblast apoptosis increased after intervention with the TZDs; in this case, there was no significant difference in the action of the 2 agonists. Thiazolidinediones caused no obvious alteration in mRNA levels for LRP5/6, GSK3β, and TCF7L2, key factors involved in Wnt signaling. At protein level, the expression of β-catenin significantly decreased, and p-GSK3β significantly increased. As a member of the Wnt family, Wnt3a can activate Wnt/β-catenin signaling in osteoblasts as well as increase the accumulation of β-catenin in the cytoplasm.39-41 Wnt proteins prolong survival of osteoblasts and progenitors of osteoblasts via the Src/ERK and PI3K/Akt signaling cascades. 39 Canonical, or β-catenin dependent, Wnt signaling is regulated to control the progression of mesenchymal cells into either chondrocytes or osteoblasts and is also required for the progression of osteoblast progenitor cells. 42 We used recombinant mouse Wnt3a protein to activate Wnt signaling; 50 ng/mL Wnt3a enhanced proliferation and reduced apoptosis, as expected. There was no obvious alteration in mRNA levels for LRP5/6, GSK3β, and TCF7L2. At protein level, the expression of β-catenin increased notably, and p-GSK3β decreased.
While both PPARγ and Wnt signaling have obvious effects upon the osteoblasts individually, it is important to try and understand their relationship. Treating the osteoblasts with both Wnt3a and TZDs together to some extent reduced the effect of the TZDs, as proliferation was increased, but apoptosis remained high. Both PPARγ and Wnt are widely expressed in organisms, and other studies have shown a complex pattern of cross-regulation; for example, in adipogenesis, canonical Wnt signaling represses PPARγ gene expression. 43 Peroxisome proliferator-activated receptor γ inhibits hepatocyte cell growth and attenuates cyclin D1 transcription, and the process is associated with an inhibition of β-catenin expression at a posttranslational level. 37 β-Catenin activates cyclin D1 to promote cell division in osteoprogenitor cells. 44 In mice, loss of cyclin D1 disrupted bone mineralization and delayed ossification and probably interfered with the proliferation of naive cells early in bone development. 45 In mice of advancing age, the decrease in osteoblast number is related to a decrease in canonical Wnt signaling and increased PPARγ; oxidized lipids acting as ligands for PPARγ promote its binding to β-catenin, reducing β-catenin levels, and this attenuates Wnt3a stimulation of osteoblast proliferation and differentiation. 46 This role for PPARγ in aging mice also shows a potential relationship between oxidized lipids, PPARγ, and increased apoptosis. 46 For preadipocytes to differentiate into adipocytes, Wnt signaling needs to be actively suppressed because sustained overexpression of Wnt1 or Wnt10b blocks adipogenesis in 3 T3-L1 preadipocytes, and an extensive down-regulation of β-catenin expression is coincident with terminal differentiation. 47 Wnt proteins and many signaling factors in the pathway, including β-catenin, GSK3β, and Tcf-4, influence the gene expression and activity of PPARγ. 38 Inducers of osteoblastogenesis, such as bone morphogenetic protein 2 and Wnt ligands, use different mechanisms to suppress the transactivation function of PPARγ during osteoblastogenesis from mesenchymal stem cells. 48 Signaling via the canonical Wnt/β-catenin pathway inhibits PPARγ mRNA expression. 48 In osteoblastic MC3T3-E1 cells, we observed that after PPARγ was activated by 20 μM Pio and Rosi, a reduced cell proliferation was accompanied by up-regulation of p-GSK3β expression and down-regulation of β-catenin protein level; it seems that the enhanced degradation of β-catenin by PPARγ requires GSK3β activity. Meanwhile, when we activated Wnt/β-catenin signaling, PPARγ mRNA expression was down-regulated as detected by RT-PCR. These results demonstrate that Wnt and PPARγ signaling are connected and regulate cell proliferation in osteoblasts together. The activation of PPARγ inhibits Wnt/β-catenin signaling through down-regulation of β-catenin to decrease proliferation; on the other hand, initiation of the Wnt pathway decreases PPARγ mRNA expression and counteracts PPARγ signaling.
This study has some limitations, while these results suggest a relationship between PPARγ and Wnt signaling, further research will be required to fully understand the detailed mechanism by which this relationship occurs. The study was undertaken on 1 cell line in vitro; to ensure that these results are true in other cells, more cell lines should be used in future studies, and animal models would provide information at the in vivo level.
In summary, we investigated the association between PPARγ and the Wnt signaling pathway by using TZDs to activate PPARγ in MC3T3-E1 cells and then inhibited the Wnt signaling pathway. Cell proliferation was decreased on PPARγ activation but in part restored when Wnt signaling was activated. Apoptosis was increased on PPARγ activation, but this effect was not reversed by activation of Wnt signaling. Peroxisome proliferator-activated receptor γ activation had no influence on mRNA levels of components of the Wnt signaling pathway but decreased β-catenin and increased p-GSK3β protein expression. Wnt signaling activation increased β-catenin protein expression and decreased p-GSK3β protein expression even when PPARγ was activated. We conclude that activation of Wnt/β-catenin signaling may lessen the activity of PPARγ and ameliorate the inhibition functions of PPARγ. We hypothesize that activation of PPARγ induces lower cell proliferation through Wnt signaling, but the details of this relationship remain to be discovered and will require further investigation.