
Abstract
The formation and maintenance of social attachments are fundamental to human biology. Because deficits in the ability to form such attachments are associated with a variety of psychological disorders, an understanding of the neural basis of social attachment may provide insights into the causes of such disorders. Comparative studies using several closely related species of voles that display different social organizations and behaviors have begun to provide important insights into the neurochemical events underlying social attachment. Here we review recent developments in the study of social attachment, focusing on the roles of specific neurochemical systems in pair-bond formation.
Interpersonal interactions, such as parent-child, spousal, and work relationships, all rely to some extent on the ability to form and maintain social ties. Deficits in the ability to form meaningful social bonds are associated with human psychological impairments, such as schizophrenia and autism. Moreover, the loss of a significant relationship can exacerbate depression. Thus, understanding the neural basis of social attachment may provide insights into the causes of some psychological disorders.
THE VOLE MODEL
Only about 3% of mammalian species are typically monogamous, and there appears to be no phylogenetic pattern to explain the distribution of monogamy among mammals. Nonetheless, there is evidence that the neurochemical basis of pair bonding may be similar among mammalian species (Insel & Young, 2000); thus, observations in one species may be applicable to others. The microtine rodents (voles) provide an excellent animal model for comparative studies on the neurochemical bases of social attachment. Although closely related, different vole species display a range of life strategies and social behaviors. Meadow and montane voles are promiscuous and show little in the way of social attachment or paternal care of offspring. In contrast, the prairie and pine voles are monogamous and exhibit strong social attachments. In prairie voles, the more studied of the latter two species, pairs share a common nest even beyond the breeding season, and both sexes provide parental care and display aggression against unfamiliar individuals.
Studying complex behaviors such as pair bonding requires a reliable behavioral index. Mating facilitates pair bonding in monogamous voles and provides a benchmark by which the “beginning” of pair-bond formation may be measured. Prairie voles display pair bonds after as few as 6 hr of mating, whereas 24 to 48 hr of cohabitation are needed for pair-bond formation in the absence of mating. This difference has been exploited in the development of the partner-preference test, a choice test in which an animal has the option of associating either with a familiar partner or with an unfamiliar stranger (Fig. 1). Prairie voles paired for 6 hr without mating are equally likely to associate with either the partner or the stranger, but will show a preference for the partner after manipulations designed to facilitate pair bonding. Conversely, voles that mate repeatedly over 24 hr reliably display a robust preference for the familiar partner, although manipulations designed to interfere with pair bonding disrupt this preference. This behavioral paradigm has been employed widely to study the mechanisms underlying pair bonding.
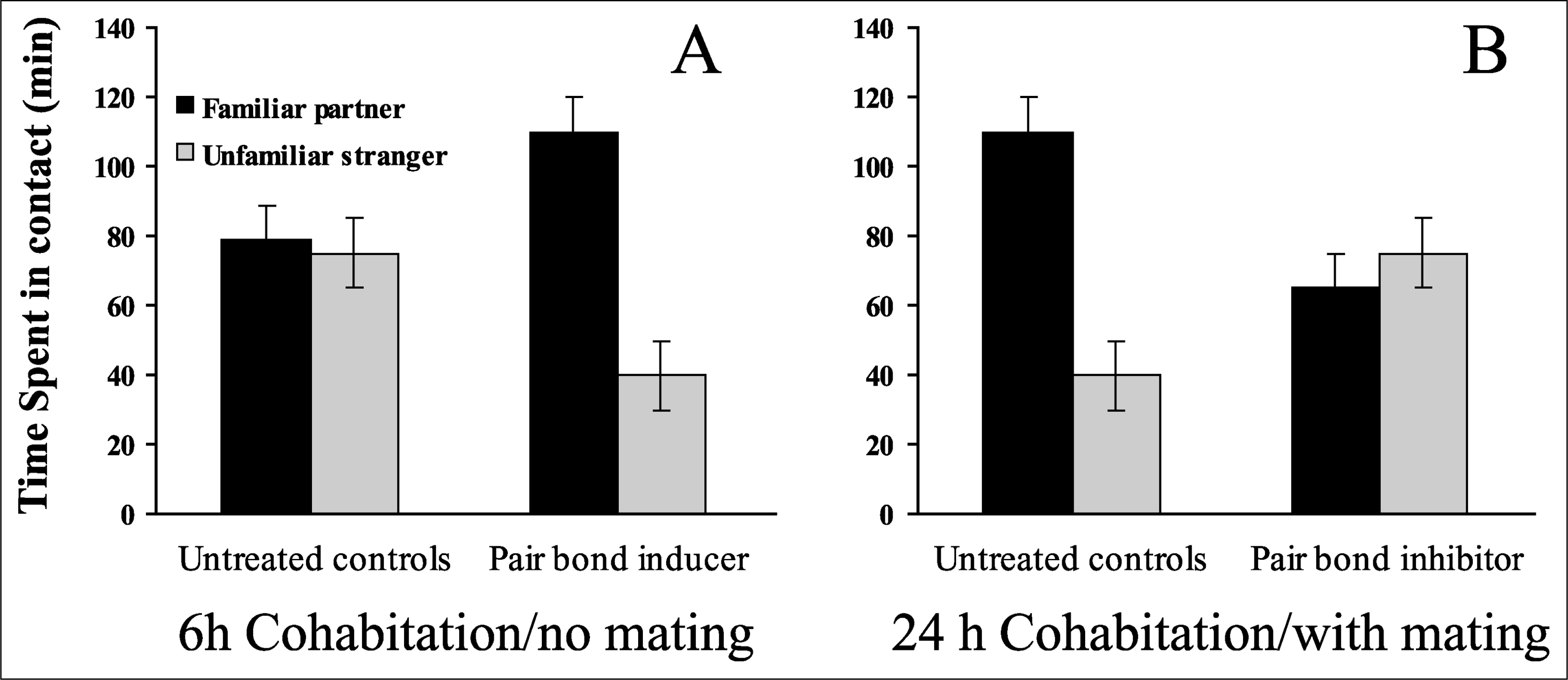
Typical results for control and experimental conditions in the 3-hr partner-preference test of pair bonding. The graph in (a) shows typical results for unmated pairs housed together for 6 hr. This amount of contact is generally insufficient to yield a preference for the familiar partner (left set of bars), but a partner preference can be induced experimentally (right set of bars). The graph in (b) shows typical results for mated pairs housed together for 24 hr. This amount of contact is usually sufficient to yield a preference for the familiar partner, but the formation of such preferences can be inhibited experimentally.
Here we review recent research examining the neurochemistry of pair bonding using the vole model. We focus on four neurochemicals—vasopressin, oxytocin, dopamine, and the stress hormone corticosterone—and briefly discuss their involvement in pair-bond formation in prairie voles.
NEUROCHEMICAL ACTIVITY
The effects of neurochemicals released within the brain are mediated by specialized proteins known as receptors that are located either on the cell surface or within the cell. Binding of a neurochemical to its receptor initiates an array of responses within the target cell. For many neurochemicals, there are different receptor subtypes that vary in the ability to bind the neurochemical and in the effects produced after binding. The receptor subtype and location within the brain where the receptor is produced are dictated by receptor genes that contain both directions for producing the receptor itself (gene expression) and promoter regions that control where in the brain the receptor is present. In many cases, drugs that mimic (agonists) or block (antagonists) the effects of neurochemicals have been developed. These drugs have allowed detailed analysis of the roles played by neurochemicals in a variety of behaviors, including pair-bond formation.
NEUROCHEMICALS INVOLVED IN PAIR BONDING
Vasopressin
Vasopressin is synthesized in the brain and strongly influences behavior and cognition when released centrally (within the brain). The distribution of vasopressin-synthesizing cells and their fibers in the vole brain shows pronounced sexual dimorphism, as in almost all rodent species that have been examined. Many of the same areas that show sexual dimorphism have been implicated in pair bonding. For example, when male prairie voles are exposed to an individual of the opposite sex, vasopressin appears to be released in the lateral septum, an area that shows sexual dimorphism and is involved in recognition of individuals. The fact that this does not occur in females, or in either sex in promiscuous voles (Bamshad, Novak, & De Vries, 1993), suggests that vasopressin mediates pair bonding in male prairie voles. This hypothesis is supported by studies showing that in male prairie voles, central administration of vasopressin induces pair bonding in the absence of mating, whereas a vasopressin receptor antagonist blocks mating-induced pair bonding (Winslow, Hastings, Carter, Harbaugh, & Insel, 1993). Using markers of neuronal activation, we have recently shown that some vasopressin-containing regions are activated during mating in prairie voles, and that site-specific administration of vasopressin into the lateral septum of male prairie voles induces pair bonding without mating (Liu, Curtis, & Wang, 2001).
In contrast to its effects in monogamous voles, central vasopressin administration does not induce pair bonds in promiscuous voles, suggesting fundamental differences in the vasopressin systems of the two types of voles. How might these species differences be mediated? Comparative studies have shown that throughout postnatal development and into adulthood, monogamous and promiscuous voles differ in the distribution pattern of vasopressin receptors (Wang, Young, Liu, & Insel, 1997). Central vasopressin binds primarily to vasopressin V1a receptors. Prairie and montane voles have almost identical gene sequences for the receptor itself, but differ in the promoter region. Could differences in gene promoters result in species-specific patterns of receptor distribution? Recently, genetically altered mice that express the prairie vole vasopressin receptor gene have been produced. These mice express vasopressin receptors in the brain in a pattern similar to that of monogamous voles, and in fact, show increased affiliative behavior after administration of vasopressin (Young, Nilsen, Waymire, MacGregor, & Insel, 1999).
Oxytocin
Oxytocin also plays an important role in pair-bond formation, and in many ways, the oxytocin system in voles parallels that for vasopressin. The first experiments examining the involvement of oxytocin in pair bonding were based in part on observations that oxytocin facilitated the formation of another type of social bond—that between mother and offspring. Administration of oxytocin into the lateral ventricles (large cavities within the brain; drugs administered in the ventricles may reach nearly any part of the brain) in female prairie voles can indeed induce pair bonding, whereas an antagonist specific to the oxytocin receptor blocks both mating- and oxytocin-induced pair bonding (Williams, Insel, Harbaugh, & Carter, 1994). Similar treatments are ineffective in promiscuous vole species.
Is the species-specific effect of oxytocin, like that of vasopressin, receptor mediated? Comparative studies on the distribution of oxytocin-containing cells and fibers within the brain do show subtle species differences between voles with differing social structures, but no consistent pattern differentiates social and nonsocial species. However, the distribution pattern of oxytocin receptors is most similar among vole species that share similar social structures (Insel & Shapiro, 1992). Again, as we have seen with vasopressin, the species-specific receptor distribution patterns are apparent throughout development and are not the result of different receptor subtypes, because all voles express the same type of oxytocin receptor. Are the species differences in the distribution of oxytocin receptors the result of differences in the promoter region for oxytocin, much as we suggested for vasopressin? The data addressing this question are not conclusive; however, subtle differences in the oxytocin promoter region may be sufficient to drive differential expression. Support for this possibility came when researchers were able to cause a reporter gene (a gene not normally expressed) to be expressed in socially relevant brain regions in mice by linking the reporter gene to the oxytocin promoter from social voles (Young et al., 1997).
Interestingly, after the birth of young, the distribution of oxytocin receptors in the nonsocial montane vole female grows to resemble the distribution seen in social voles (Insel & Shapiro, 1992). This to some extent brings the field full circle, because the original experiments examining a potential role for oxytocin in pair bonding were based on oxytocin's role in maternal bonding. The maternal “circuit” (brain areas associated with maternal behavior) includes most of the neural components implicated in pair bonding. Thus, one possibility for the origin of pair bonding is that pair-bonding species have co-opted the mechanism (or mechanisms) by which maternal bonds are formed. This possibility is further supported by observations that even sexually naive male prairie voles display maternal-type behaviors when exposed to pups, and that prairie vole mothers display considerably more maternal care than do meadow vole mothers.
The data we have presented suggest sex-specific roles for oxytocin and vasopressin: Oxytocin is the “pair-bond hormone” in females, whereas vasopressin performs this function in males. Recent studies, however, have shown that the roles of vasopressin and oxytocin in the regulation of pair bonding may not be quite so straightforward. Central administration of either compound at relatively high doses can induce pair bonding in both sexes. Similarly, pair bonding in both sexes can be blocked by antagonists specific to either type of receptor (Cho, DeVries, Williams, & Carter, 1999). Further, when injected into the lateral septum in male prairie voles, oxytocin, like vasopressin, can induce pair bonding (Liu et al., 2001). These data indicate that both vasopressin and oxytocin are involved in pair bonding in both sexes of prairie voles. Despite these findings, however, there are important sex differences in responses to various doses of these neurochemicals, indicating that the sexes may differ in their relative sensitivities to vasopressin and oxytocin. Further, when sex differences are found, they tend to reinforce the notion that vasopressin is more effective in males than in females, whereas oxytocin is more effective in females than in males. Finally, sex differences may also exist in other systems that interact with vasopressin and oxytocin in regulating pair bonding.
Dopamine
Drugs targeting the dopamine system are frequently used to treat disorders associated with social attachment, such as autism and social phobia. Further, dopamine is strongly implicated in learning and memory, processes likely to be important in pair bonding. The first evidence for dopamine's involvement in pair bonding came from observations that, in female prairie voles, peripheral (outside the brain) administration of apomorphine, a dopamine agonist, induced pair bonding absent mating, whereas haloperidol, a dopamine antagonist, blocked mating-induced pair-bond formation (Wang et al., 1999). The same study found that dopamine acts on the D2 family of dopamine receptors to regulate pair bonding.
In female prairie voles, dopamine is released in the nucleus accumbens during mating. Blockade of D2 receptors, but not the D1 family of receptors, in the nucleus accumbens inhibits mating-induced pair bonding, whereas site-specific administration of D2, but not D1, agonists induces pair bonding absent mating (Gingrich, Liu, Cascio, Wang, & Insel, 2000). Recently, we have found that dopamine also is involved in pair bonding in male prairie voles. Interestingly, facilitation of pair bonding requires higher doses of dopamine agonists in female voles than in male voles, indicating a potential sex difference. However, at very high doses, apomorphine becomes ineffective in males. Apomorphine activates primarily D2 receptors, but at high doses, it may also activate D1-type receptors. Thus, activation of D1 receptors may in fact inhibit pair-bond formation (Aragona, Liu, Curtis, Stephan, & Wang, in press).
Corticosterone
Prairie voles have very high basal levels of corticosterone circulating in their blood. When corticosterone levels are reduced after removal of the adrenal glands (adrenalectomy), female prairie voles form pair bonds after as little as 1 hr of nonsexual cohabitation with a male (DeVries, DeVries, Taymans, & Carter, 1996). Stress-induced or artificial elevation of corticosterone in females actually seems to produce an aversion to the familiar partner. Interestingly, the effects of corticosterone on pair bonding are sexually dimorphic: In male prairie voles, adrenalectomy inhibits pair-bond formation, whereas stress-induced or artificial elevation of corticosterone facilitates pair-bond formation. These opposite effects occur despite the fact that stress elevates circulating levels of corticosterone similarly in male and female prairie voles.
Neurochemical Interactions
Although the neurochemicals we have discussed all have been implicated in pair bonding, it is unknown whether they act in concert or independently to regulate behavior. In addition, the nature of their interactions, if any, and the brain circuits in which such interactions occur remain to be determined. Recently we have begun to address these questions. In male prairie voles, vasopressin administration in the lateral septum induces pair bonding, but this effect is blocked by co-administration of either vasopressin or oxytocin receptor antagonists, suggesting that access to both kinds of receptors is essential for vasopressin to induce pair bonding (Liu et al., 2001). Administration of either oxytocin or D2 agonists in the nucleus accumbens, which contains both oxytocin and D2 receptors, induces pair bonding in female prairie voles. Interestingly, the abilities of both oxytocin and dopamine to induce pair bonds can be blocked by antagonists to either type of receptor. These results suggest that oxytocin and dopamine act synergistically in the nucleus accumbens to regulate pair bonding.
FUTURE DIRECTIONS
Considerable work is still needed to provide a comprehensive understanding of the neurochemical basis of pair bonding. Very little is known about responses at the cellular level in brain areas that are involved. We recently have begun to examine the control of gene transcription, which may affect learning, in mediating pair-bond formation. In addition, studies examining the role of tissue-specific gene expression in social behavior have begun. Monogamous voles that overexpress the vasopressin receptors in some brain areas display increased affiliative behavior (Pitkow et al., 2001). These results highlight the potential of genetic research in the study of social attachment. Further, recent studies have revealed that new neurons are constantly being produced in adult mammalian brains. We have found that exposure to the opposite sex can significantly increase the number of new neurons in brain areas important for pair-bond formation in prairie voles (Fowler, Liu, Ouimet, & Wang, 2002). The prairie vole, therefore, may provide a model in which to investigate the role of new neurons in social behaviors such as pair-bond formation. Finally, researchers and practitioners must begin to apply the lessons learned in the study of pair bonding to the diagnosis and treatment of human psychological disorders. For example, current methods of treatment for schizophrenia often produce negative side effects, and understanding the neuroanatomy and neurochemistry of social attachment may allow more specifically targeted treatments to be developed.
Footnotes
Acknowledgements
We would like to thank Y. Liu, C. Fowler, B. Aragona, and J. Stowe for helpful comments. Preparation of this article was partly supported by National Institutes of Health Grants NICHD 40722 (to J.T.C.) and NIMH 54554 and 58616 (to Z.W.).