
Abstract
In a standard inhibition-of-return (IOR) paradigm using a manual key-press response, we examined the effect of IOR both on the amplitude of early sensory event-related brain potential (ERP) components and on the motor-related lateralized readiness potential (LRP). IOR was associated with a delay of premotor processes (target-locked LRP latency) and reduced sensory ERP activity. No effect of IOR was found on motor processes (response-locked LRP latency). Thus, IOR must arise at least in part from changes in perceptual processes, and, at least when measured with manual key presses, IOR does not arise from inhibition of motor processes. These results are consistent with the results of attention-orienting studies and provide support for an inhibition-of-attention explanation for IOR.
In a typical visual spatial-cuing experiment with uninformative peripheral cues, reaction time (RT) to cued-location targets is facilitated (relative to that for targets at uncued locations) at short cue-target stimulus-onset asynchronies (SOAs) and inhibited at longer SOAs. This latter inhibitory effect is called inhibition of return (IOR). Since it was first reported by Posner and Cohen (1984), IOR has attracted a great deal of research interest (e.g., Klein, 2000). This interest is primarily motivated by the belief that IOR is closely associated with both covert and overt orienting of attention. Posner, Rafal, Choate, and Vaughan (1985) proposed that IOR serves to bias the visual system to acquire novel information at new locations. Expanding on this proposal, Klein (1988) suggested that IOR serves to facilitate visual search by biasing orienting responses away from recently inspected locations. This provocative proposal remains a popular topic of debate and has inspired numerous empirical studies (e.g., Klein, 2000). Given that compelling evidence of inhibitory effects in visual search has been obtained (Klein & MacInnes, 1999; Müller & von Mühlenen, 2000; Takeda & Yagi, 2000), an understanding of the mechanisms underlying IOR may provide valuable insights into how visual information is selected.
Despite a significant accumulation of empirical knowledge, no consensus has yet been reached regarding the stages of information processing affected by IOR. Explanations of IOR can be broadly classified into two groups, those that involve perception and attention and those that involve response selection and motor processes (e.g., Taylor & Klein, 1998). These types of explanations are not mutually exclusive, and IOR may arise from multiple mechanisms, or from a single mechanism that affects multiple stages of processing. Posner et al. (1985) proposed that IOR arises from attention being inhibited from returning to previously attended locations. Given the considerable evidence that spatial attention enhances perceptual processing (e.g., Wright & Ward, 1998), any mechanism that inhibits attention from being oriented to a spatial location should result in a relative inhibition of perceptual processing at that location. It has also been suggested that IOR may inhibit perceptual processing independently of attention (e.g., Posner & Cohen, 1984; Reuter-Lorenz, Jha, & Rosenquist, 1996). Evidence that IOR arises from an inhibition of perceptual processes has come from experiments showing that IOR affects the accuracy of unspeeded target-discrimination responses (Handy, Jha, & Mangun, 1999; Klein & Dick, 2002). In addition, Reuter-Lorenz et al. (1996) demonstrated that the magnitude of IOR and attentional facilitation were similarly affected by changes in target intensity and modality, providing support for the attentional-orienting explanation of IOR.
Other researchers have suggested that IOR may be associated with changes in response-related processes. Klein and Taylor (1994) proposed that IOR may arise from a reluctance to respond to events at the cued location and that the inhibition may arise in a spatial motor map that directs action. Alternatively, Tassinari, Aglioti, Chelazzi, Marzi, and Berlucchi (1987) proposed that IOR arises from inhibition in the motor system that results from suppression of overt orienting toward the cue. A response-related effect could conceivably arise at either decisional or motor stages of processing. Response-level explanations have received support from evidence that IOR is associated with a more conservative response criterion on valid trials than on invalid trials (Ivanoff & Klein, 2001) and that IOR can affect oculomotor programming (Ro, Pratt, & Rafal, 2000). Given that the currently available evidence provides support for both perceptual and response-related mechanisms, it has been proposed that IOR may arise from multiple mechanisms (Kingstone & Pratt, 1999; Taylor & Klein, 2000).
One technique that has proven to be especially useful in investigating the information processing stages generating experimental effects is the recording of event-related brain potentials (ERPs) of the electroencephalogram (EEG). Relatively few studies have examined the ERP consequences of IOR, however, and the results have, so far, been somewhat inconsistent. McDonald, Ward, and Kiehl (1999) found that IOR was associated with a reduction in the amplitude of the visual P1 component and a negative difference (Nd) between ERPs on valid-cue and invalid-cue trials in the P2 latency range. These results indicate that IOR is associated with a suppression of perceptual processing in extrastriate visual areas. However, two other studies challenged this conclusion. Hopfinger and Mangun (1998) found a significant P1 reduction accompanied by a nonsignificant 2-ms IOR effect. Conversely, in a subsequent study (Hopfinger & Mangun, 2001), they found a significant 13-ms IOR effect but a nonsignificant P1 reduction. Although both of these studies found behavioral and P1-amplitude effects in the same direction as in the study by McDonald et al. (1999), Hopfinger and Mangun (2001) concluded that IOR was dissociable from the P1 reduction. Because the available data seem to allow no firm conclusions about the ERP correlates of IOR that would clarify its mechanism, additional study of these correlates seems warranted.
In the present study, we investigated the electrophysiological correlates of IOR in a visual form discrimination task. Unlike in previous studies, we examined the effect of uninformative visual cues both on the amplitude of early sensory ERP components and on the motor-related lateralized readiness potential (LRP). The LRP is an electrophysiological measure of brain activity generated in motor cortex that is related to the selection and preparation of motor responses (see Coles, 1989, and Fig. 1). The latency of LRP onset is dependent on response selection, so that the interval between target onset and the onset of the target-locked LRP provides a relative measure of the duration of processes involved in stimulus evaluation and response selection. Similarly, the interval between the onset of the response-locked LRP and the response provides a relative measure of the duration of motor processes (e.g., motor planning and execution). If IOR arises from inhibition of motor processes, then the interval between the onset of the response-locked LRP and the response should be longer on valid-cue trials than on invalid-cue trials. Conversely, if IOR arises from inhibition of processing occurring prior to the onset of motor planning and execution, then the target-locked LRP should begin sooner on invalid-cue trials than on valid-cue trials. The presence of both effects would implicate both perceptual-attentional and motor mechanisms of IOR. In addition, if a premotor effect of IOR due to an inhibition of perceptual processing is present, IOR should be accompanied by a reduction in the amplitude of the short-latency ERP components at occipital sites.
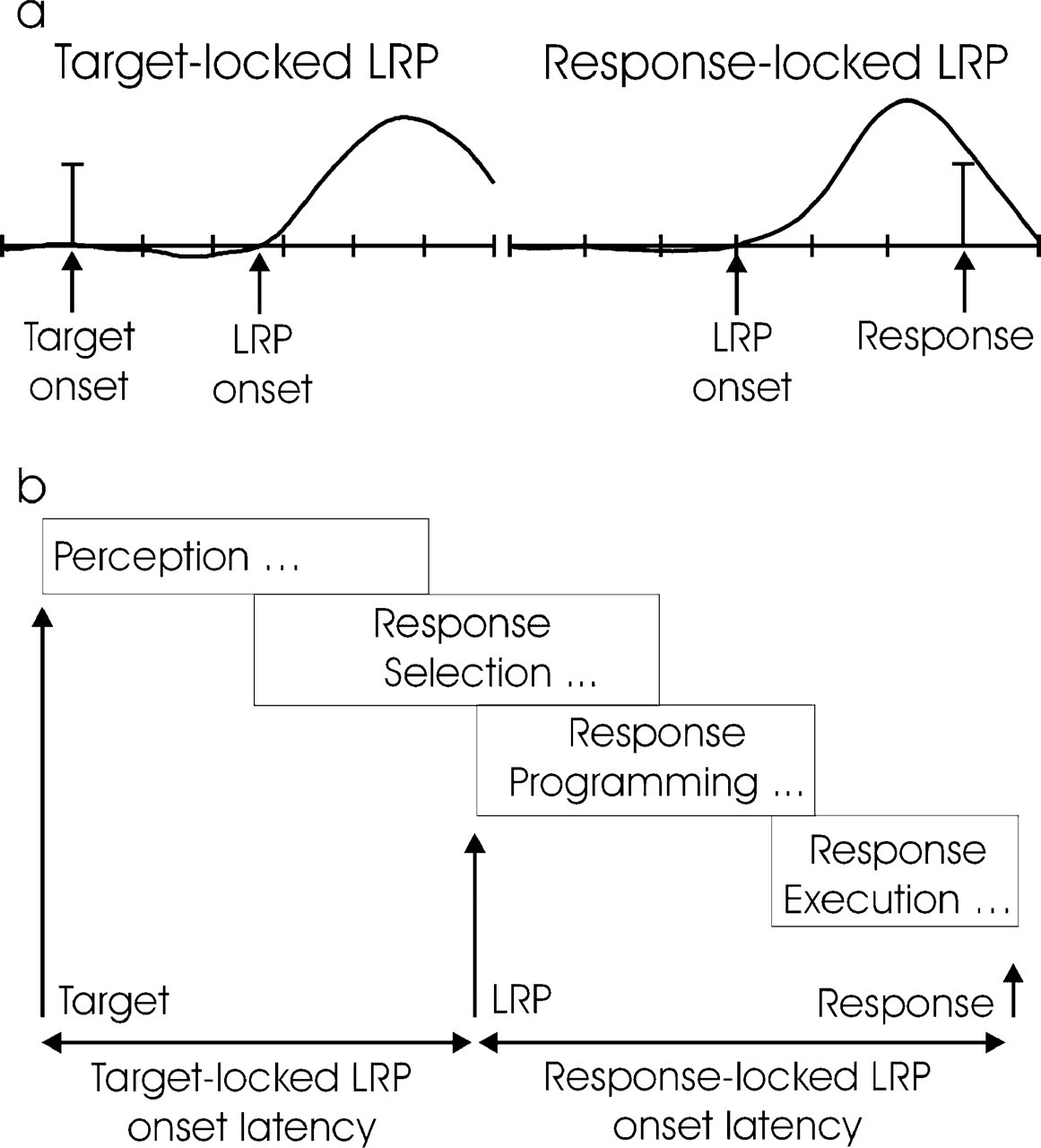
Relationship between lateralized readiness potential (LRP) onsets and information processing stages: target- and response-locked LRPs indicating time of LRP onset (a) and model of target-related information processing between stimulus and response (b). See the text for details.
METHOD
Participants viewed a computer monitor from a distance of 42 cm and were instructed to maintain fixation on a centrally located fixation cross during the experimental blocks. The screen background was black, and two gray square-outline boxes (1.5°×1.5°) were centered 5° above and below fixation. After a 700-ms intertrial interval, each trial began with a 133-ms offset of the fixation cross, followed 800 ms later by the cue. The cue consisted of a 200-ms brightening of one of the two square boxes and was equally likely to occur at either location. Two hundred fifty milliseconds after cue offset, a white circle 0.75° in diameter was presented at fixation for 160 ms. This reorienting event was intended to redirect the participants' attention back to fixation from the cued location. After a variable delay of 290 to 590 ms, the target was presented for 1,000 ms. The target, either a plus sign (+) or the letter x (0.75°×0.75°), was presented with equal probability within one of the two peripheral boxes (chance coincidence of cue and target locations). Participants pressed the “z” key on a standard keyboard if the x was presented and the “/” key if the plus sign was presented. Both speed and accuracy were emphasized in the instructions. The total cue-target SOA ranged from 900 to 1,200 ms. This entire interval is well within the range of SOAs that produce a reliable IOR effect and also ensures that overlap of high-frequency components of ERPs to cues, reorienting events, and targets is minimized. Trials on which cue and target occurred at the same location were classified as valid-cue trials, whereas trials on which cue and target occurred at the opposite locations were classified as invalid-cue trials.
Twenty participants performed one practice and eight experimental blocks of 72 trials. Two participants were excluded from the analysis because of excessive eye movement artifacts.
Scalp potentials were recorded from the following scalp sites: C1, C2, C3, C4, Pz, PO7, and PO8. These electrodes were referenced to the right mastoid and subsequently rereferenced to averaged mastoids. Eye position was monitored by both the horizontal and the vertical electro-oculogram (EOG). The EEG and EOG were sampled at 250 Hz. Trials containing eye movement, muscle, and blocking artifacts were removed prior to averaging by applying automated artifact-detection routines. In addition, trials with errors and those with RTs outside of 100 to 1,000 ms were excluded from the analysis. ERPs were calculated separately for valid-cue and invalid-cue trials at electrodes sites Pz, PO7, and PO8. Valid-cue and invalid-cue LRPs were calculated at the C1-C2 and C3-C4 electrode pairs using the averaging method (Coles, 1989). To reduce the smearing of the LRP slope due to time jitter, we excluded trials with RTs greater than 1.5 SDs from the mean in each condition from the LRP averages. After averaging, the ERPs and LRPs were low-pass filtered (12 Hz and 4 Hz, respectively) to eliminate high-frequency artifacts in the waveforms.
Target- and response-locked LRP onsets were determined using the one-degree-of-freedom (1DF) regression technique (Mordkoff & Gianaros, 2000). 1 Differences in LRP onsets were statistically assessed with a jackknifing procedure (J. Miller, Patterson, & Ulrich, 1998; R.G. Miller, 1974). 2 For the ERP data, amplitude differences were analyzed by separate repeated measures multivariate analyses of variance for each effect. The mean amplitudes corresponding to the latency of the P1 (100–140 ms) and N1 (156–196 ms) components were analyzed at sites PO7 and PO8. These 40-ms latency windows are centered on the peaks of the P1 and N1 components in the grand-average waveforms. The mean amplitude of the Nd was measured in a latency range (240–280 ms) corresponding to the component's maximal amplitude and analyzed separately at parietal (Pz) and occipital (PO7, PO8) sites.
RESULTS AND DISCUSSION
As expected, participants were significantly faster in responding to targets on invalid-cue trials (564 ms) than on valid-cue trials (585 ms), F(1, 18)=29.8, p<.0001; this is a typical IOR effect. There was no significant difference between the error rates on valid (4.2%) and invalid (5.4%) trials. The target-locked LRP began at a shorter latency for invalid-cue than for valid-cue trials at both the C1-C2 and C3-C4 electrode pairs, t(18)=2.1, p=.05, and t(18)=2.2, p<.05, respectively (Fig. 2a, Table 1). By contrast, no onset difference was found between valid-cue and invalid-cue response-locked LRPs at either electrode pair (both t values <1; Fig. 2a, Table 1). These results strongly indicate that IOR in this standard paradigm arises from delays in perceptual and decisional processes that occur before LRP onset, and not from later-occurring motor processes.
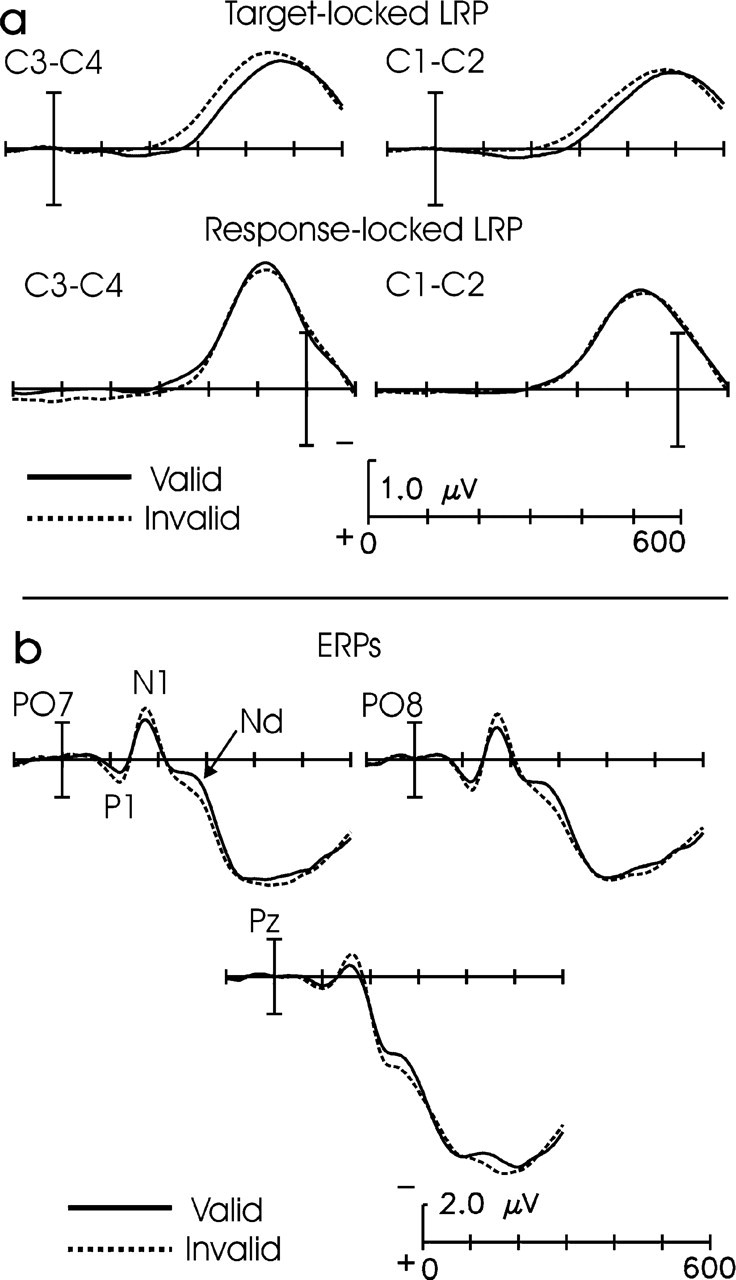
Electrophysiological data: target- and response-locked lateralized readiness potentials (LRPs) for valid and invalid trials (a) and event-related potentials (ERPs) to valid and invalid targets at posterior electrode sites (b). Vertical bars indicate target onset for target-locked waves and time of response for response-locked LRPs. Nd=negative difference.
Mean Event-Related Potential Amplitudes and Lateralized Readiness Potential Onset Latencies
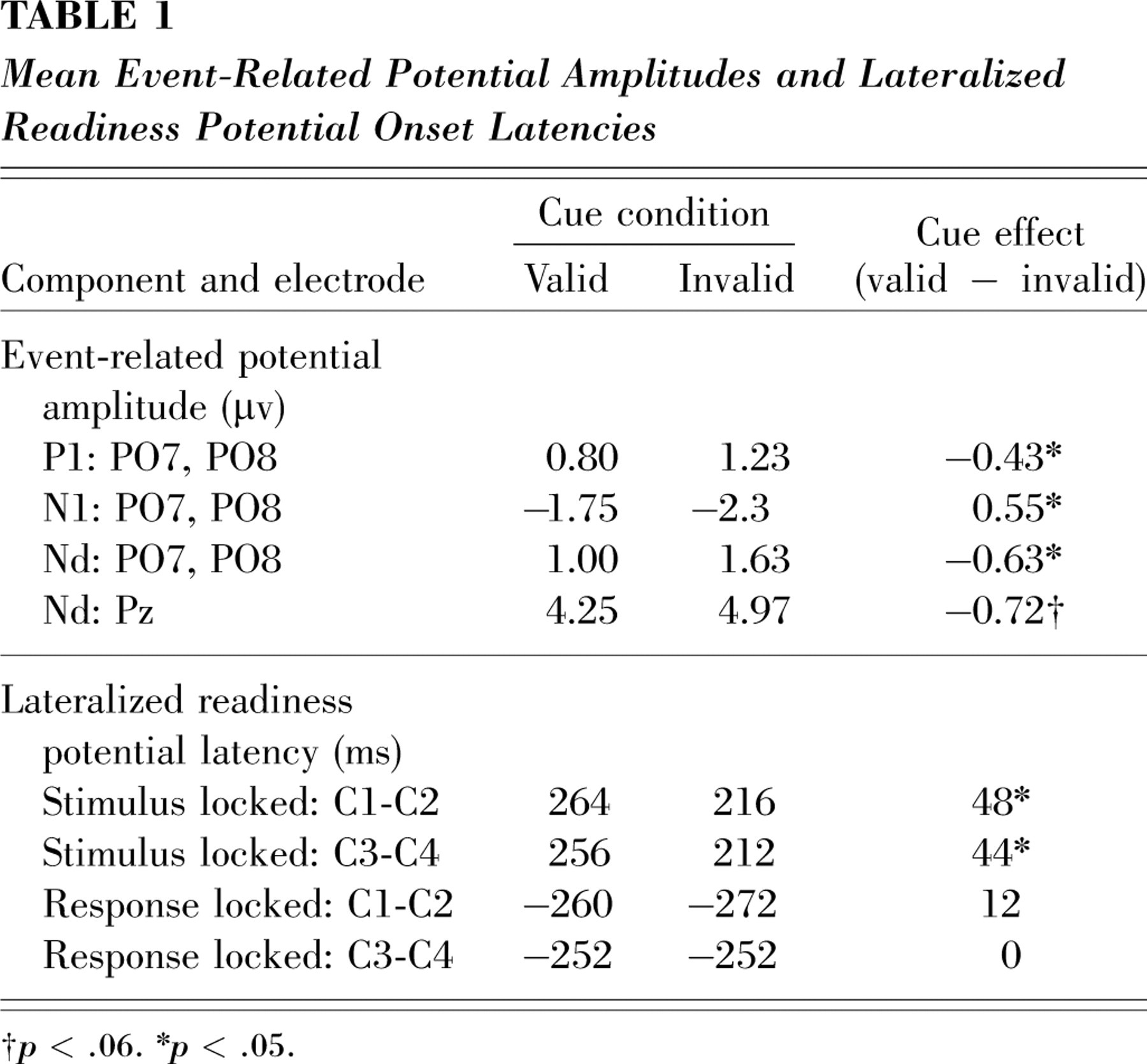
† p<.06.
∗ p<.05.
ERP results were consistent with a perceptual locus of IOR. The amplitudes of the occipital P1 and N1 components were both reduced on valid-cue relative to invalid-cue trials, F(1, 18)=4.8, p<.05, and F(1, 18)=4.8, p<.05, respectively (Fig. 2b). Dipole modeling techniques have localized both of these components to areas of extrastriate cortex (Di Russo, Martínez, Sereno, Pitzalis, & Hillyard, 2001). These effects indicate that IOR is associated with an inhibition of early sensory processing of validly cued targets relative to invalidly cued targets, an effect opposite that of spatial attention (e.g., Mangun & Hillyard, 1991). In addition, an Nd was present at both occipital and parietal sites, F(1, 18)=6.1, p<.03, and F(1, 18)=4.2, p<.06, respectively (Fig. 2b, Table 1). This effect is similar to the Nd found by McDonald et al. (1999) and may reflect either perceptual or decision processes.
CONCLUSION
The results presented here clearly show that IOR is associated with a delay in premotor processes. The target-locked LRP began at a longer latency on valid-cue than on invalid-cue trials. This effect is consistent with either a suppression of perceptual processing or a delay in response selection. The onset of the response-locked LRP was not affected, suggesting that when measured with manual key presses, IOR does not arise from inhibition of motor processes. Although it is not possible to rule out an effect of IOR on motor processes on the basis of null results, these results do indicate that premotor effects were primarily responsible for the observed differences in RT in this experiment. However, these results do not exclude the possibility that an inhibition of motor processes may contribute to the IOR effect when oculomotor and, perhaps, spatially directed pointing or reaching movements are used.
Although a change in the latency of the target-locked LRP does not distinguish between perceptual and decisional effects, the modulations of sensory ERP components indicate that IOR arises at least in part from changes in sensory processes. The amplitudes of the occipital P1 and N1 components were smaller on valid-cues trials than on invalid-cue trials. These results are consistent with the results of attention-orienting studies that have found that enhanced P1 and N1 amplitudes are accompanied by RT facilitation (e.g., Mangun, 1995). This parallel between the effects of attention orienting and IOR provides support for the inhibition-of-attention explanation of IOR. Unlike in the current study, McDonald et al. (1999) found that IOR in a simple detection task was associated only with a reduction of the P1 component. Although there are several differences between these experiments, it is possible that this difference in results arises from the nature of the response tasks. Previous research has shown that the visual N1 is associated with discrimination processes (Vogel & Luck, 2000) and that attentional enhancement of the N1 is typically observedonly for discrimination tasks (Mangun & Hillyard, 1991). Thus, our observation of a reduction in N1 magnitude in the presence of IOR is consistent with an IOR-related reversal of the effects of attentional enhancement on the N1 in our discrimination task. Taken together, the LRP and ERP effects observed in this experiment indicate that IOR must arise at least in part from changes in perceptual processes and that, at least when measured with manual key presses, IOR does not arise from inhibition of motor processes.
Footnotes
1Regression-based techniques define the onset of the LRP as the intersection of two straight lines that are fitted to segments of the LRP waveform (Schwarzenau, Faulkenstein, Hoormann, & Hohnsbein, 1998). The 1DF technique sets the preonset line to have a height and slope of zero. The terminus of the postonset line is locked to the peak of the LRP so that only the time intersection can vary. The postonset line is then found using the least squares technique to find the best fit to the LRP.
2The jackknifing procedure involves determining the LRP onset latency for N different subsample grand-averages, with each participant omitted from one of the subsamples. The values of these onset latencies across subsamples are then used to calculate a standard error for pair-wise comparisons between conditions.
Acknowledgments
This research was supported by a Natural Sciences and Engineering Research Council (NSERC) Discovery Grant to L.M.W. and by an NSERC Predoctoral Research Fellowship to D.J.P. We thank Carey Huh, Mirona Motoc, and Daryl Wile for help with collecting the data.